直立型半野生大豆葉綠體基因組分析
2023-12-04 06:21:56郭冉昊趙淑文米福貴候偉峰趙力興
草地學(xué)報(bào) 2023年11期
關(guān)鍵詞:大豆
郭冉昊, 趙淑文*, 米福貴, 候偉峰, 趙力興
(1.內(nèi)蒙古農(nóng)業(yè)大學(xué)草原與資源環(huán)境學(xué)院, 內(nèi)蒙古 呼和浩特 010000; 2.內(nèi)蒙古農(nóng)業(yè)大學(xué)植物學(xué)國(guó)家級(jí)實(shí)驗(yàn)教學(xué)示范中心,內(nèi)蒙古 呼和浩特 010000; 3.興安盟農(nóng)牧科學(xué)研究所, 內(nèi)蒙古 興安盟 137400)
葉綠體屬于一種半自主性細(xì)胞器,一般為共價(jià)閉合的環(huán)狀四分體結(jié)構(gòu),是高等植物細(xì)胞中光合作用的主要場(chǎng)所。葉綠體具有獨(dú)立的遺傳物質(zhì),即cpDNA,主要編碼與光合作用相關(guān)的上百種基因,大小介于120~180 kbp,包括長(zhǎng)單拷貝區(qū)(Large single copy,LSC)、短單拷貝區(qū)(Small single copy,SSC)及2個(gè)反向重復(fù)序列(Inverted repeat sequence,IRS)。cpDNA在物種分類、系統(tǒng)發(fā)育、遺傳多樣性、物種形成、適應(yīng)性進(jìn)化等諸多研究中具有重要作用[1-4]。隨著DNA測(cè)序技術(shù)進(jìn)步、成本降低,植物基因組學(xué)的研究也越來(lái)越深入。而相較于大豆核基因組結(jié)構(gòu)和遺傳多樣性的廣泛研究報(bào)道,大豆屬細(xì)胞質(zhì)基因組研究相對(duì)滯后[5]。
大豆屬(Glycine)屬于豆科(Leguminosae),蝶形花亞科(Papilionatae),含一年生Soja和多年生野生Glycine兩個(gè)亞屬[6]。栽培大豆(G.max)是我國(guó)重要的經(jīng)濟(jì)作物之一,與野大豆(G.soja)同屬Soja亞屬。野大豆具有蛋白含量高、抗逆性強(qiáng)、生物量較高等特點(diǎn),是重要的育種材料和優(yōu)質(zhì)飼草,但由于其莖纏繞,利用上有一定難度。除一年生栽培大豆和野生大豆外,還存在一種特殊類群,即性狀介于野生大豆與栽培大豆之間,被前蘇聯(lián)學(xué)者Skvortzow稱為的半野生大豆(G.gracilis)[7],其種子百粒重通常在3 g乃至10 g以上,形態(tài)上有黑、褐、黃、綠、雙色種皮及各種中間種皮顏色,莖呈纏繞、弱纏繞、葡匐、蔓生、半蔓生、半直立、甚至直立類型,形態(tài)豐富[8]。半野生大豆兼具栽培大豆和野生大豆性狀等特征,不僅是栽培大豆馴化研究的重要遺傳材料[5],與栽培大豆雜交,也可以比野大豆更便捷有地效拓寬大豆遺傳基礎(chǔ),創(chuàng)造優(yōu)異種質(zhì)。由于半野生大豆的百粒重與栽培大豆有重疊,而且其莖形態(tài)與野生大豆和栽培大豆也有重疊,因此半野生大豆的分類地位一直存在爭(zhēng)議。有學(xué)者認(rèn)為半野生大豆是野生大豆向栽培大豆馴化后的中間產(chǎn)物,而另一些學(xué)者提出半野生大豆是栽培大豆和野生大豆雜交后的產(chǎn)物[9-12]。
直立型半野生大豆既有半野生大豆高蛋白、多花莢、生育繁茂等優(yōu)良特性,也克服了野大豆莖纏繞,種植利用困難等問(wèn)題,具有較高的研究?jī)r(jià)值。本研究以興安盟農(nóng)牧科學(xué)研究所提供的一份直立型半野生大豆為試材,采用高通量測(cè)序技術(shù)對(duì)其cpDNA序列、密碼子偏好性、SSR位點(diǎn)及親緣關(guān)系等進(jìn)行分析,以期為半野生大豆細(xì)胞質(zhì)多樣性、系統(tǒng)分類地位、分子育種及種質(zhì)利用提供依據(jù)。
1 材料與方法
1.1 試驗(yàn)材料
試驗(yàn)材料為采自內(nèi)蒙古自治區(qū)興安盟科爾沁右翼前旗索倫鎮(zhèn)草根臺(tái)嘎查(120°54′E,46°45′N)的直立型半野生大豆。2021年秋季采收的種子于2022年種植在呼和浩特市內(nèi)蒙古農(nóng)業(yè)大學(xué)試驗(yàn)園區(qū)。
1.2 基因組DNA提取和測(cè)序
在田間采集健康新鮮植株葉片,置于液氮帶回實(shí)驗(yàn)室于-80℃冰箱存儲(chǔ),委托北京百邁客生物科技有限公司對(duì)樣品進(jìn)行高通量測(cè)序、DNA文庫(kù)構(gòu)建等工作。
1.3 葉綠體全基因組組裝和注釋
原始測(cè)序數(shù)據(jù)(raw data)經(jīng)Fastqc程序質(zhì)檢后,用trim_galore除去低質(zhì)量序列得到clean data,然后用Getorganelle[13]對(duì)cpDNA進(jìn)行組裝。進(jìn)一步借助在線工具GeSeq[14](https://chlorobox.mpimp-golm.mpg.de/geseq.html)對(duì)組裝結(jié)果進(jìn)行注釋(注釋參考基因組:NC_0229868.1)。注釋過(guò)程中,先依據(jù)注釋后IR區(qū)的位置手動(dòng)調(diào)整基因組起始位置,并將基因組按照LSC,IRb,SSC和IRa順序排列,最后將排列完成的cpDNA用在線軟CPGAVAS2[15](http://47.96.249.172:16019/analyzer/home)進(jìn)行cpDNA注釋。cpDNA物理圖譜由Chloroplot[16]R軟件包繪制。
1.4 cpDNA基本特征
用tRNAscan-SE software軟件確定編碼基因總數(shù)、編碼tRNA和rRNA的基因數(shù)目[17]。
1.5 密碼子偏好性和SSR分析
運(yùn)用CodonW 1.4.2軟件[18](http://mobyle.pas-teur/fr/cgi-bin/portal.py?from=codonw)分析材料cpDNACDS序列,獲得試驗(yàn)材料同義密碼子的相對(duì)使用度(relative synon-ymous codon usage,RSCU);采用在線網(wǎng)站(https://webblast. ipk-gatersleben. de/misa/index.php?action=1)掃描分析直立型半野生大豆cpDNA序列進(jìn)行微衛(wèi)星掃描分析,單核苷酸、雙核苷酸、三核苷酸、四核苷酸、五核苷酸和六核苷酸設(shè)置參數(shù)重復(fù)數(shù)分別不少于10,5,4,4,4,4。
1.6 野大豆及其近緣種系統(tǒng)發(fā)育樹(shù)構(gòu)建
在NCBI數(shù)據(jù)庫(kù)中下載所有已公布大豆屬植物cpDNA序列,包括Glycine和Soja兩個(gè)亞屬共8個(gè)材料同時(shí)以模式植物擬南芥[(Arabidopsisthaliana)(MZ323108.1)]、煙草[(Nicotianatabacum)(MZ707522.1)]作為外類群,將所有參試物種的cpDNA序列選擇MAFFT進(jìn)行多序列比對(duì),手動(dòng)校正,使用PhyloSuite軟件[19]中的RAxML,bootstrap為1 000,構(gòu)建系統(tǒng)發(fā)育樹(shù)。
2 結(jié)果與分析
2.1 cpDNA結(jié)構(gòu)、分類、功能及特征
供試直立型半野生大豆的cpDNA全長(zhǎng)為148 320 bp,屬典型環(huán)狀四分體結(jié)構(gòu)(圖1)。共編碼108個(gè)基因,包括66個(gè)蛋白編碼基因、29個(gè)tRNA基因和4個(gè)rRNA基因。按基因功能可將供試直立型半野生大豆cpDNA編碼基因分為3大類:57個(gè)基因表達(dá)相關(guān)基因、42個(gè)光合作用相關(guān)基因、7個(gè)開(kāi)放閱讀和其他蛋白編碼基因以及2個(gè)未知功能基因。其中基因表達(dá)相關(guān)基因包含5個(gè)小類,數(shù)量最多的為轉(zhuǎn)運(yùn)RNA基因,RNA聚合酶基因與核糖體RNA基因數(shù)量較少,僅為4個(gè)(表1)。
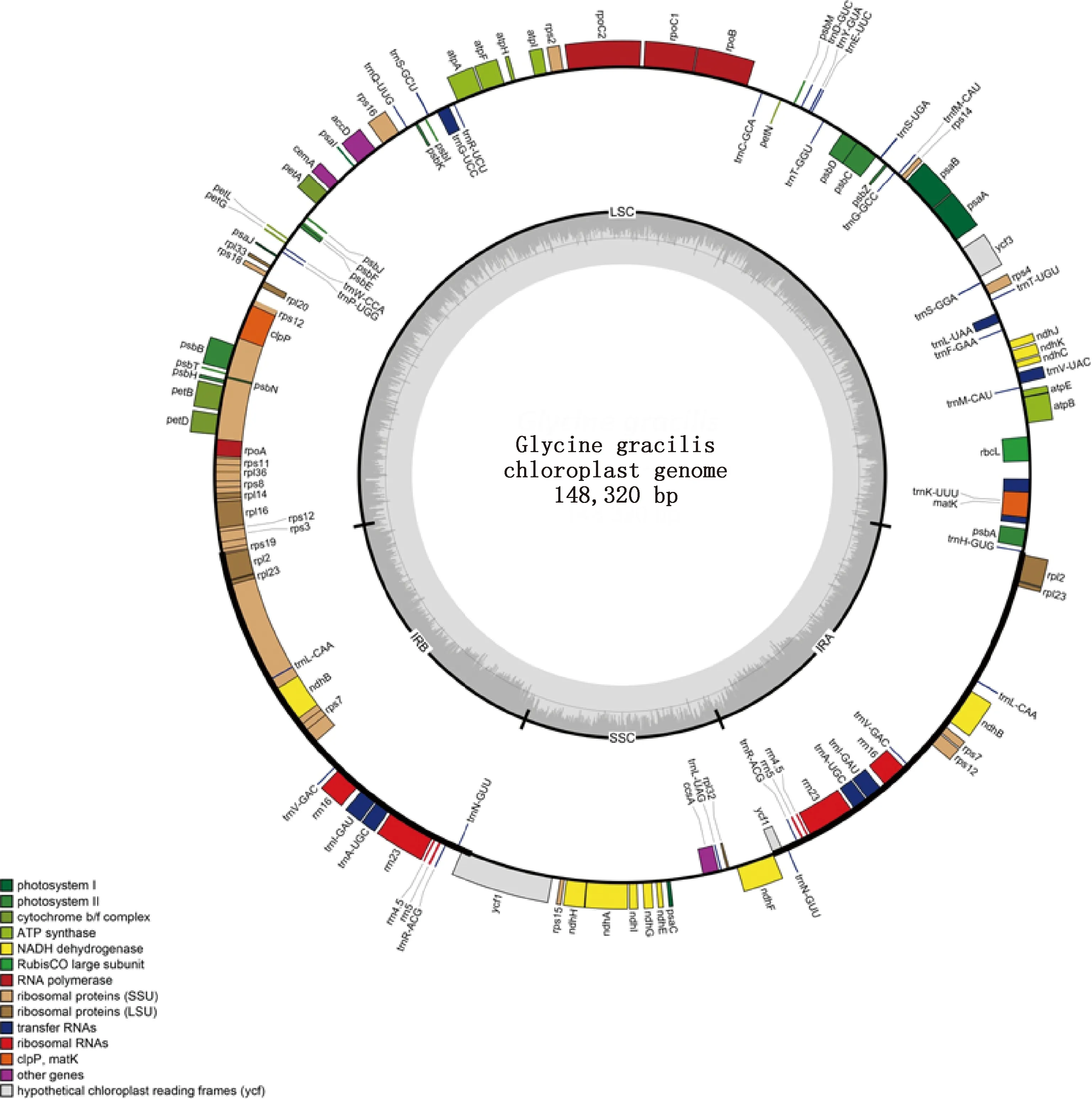
圖1 直立型半野生大豆(G.gracilis)cpDNA環(huán)形圖譜Fig.1 Circularized map of the chloroplast genome of the erect semi-wild soybean (Glycine gracilis)
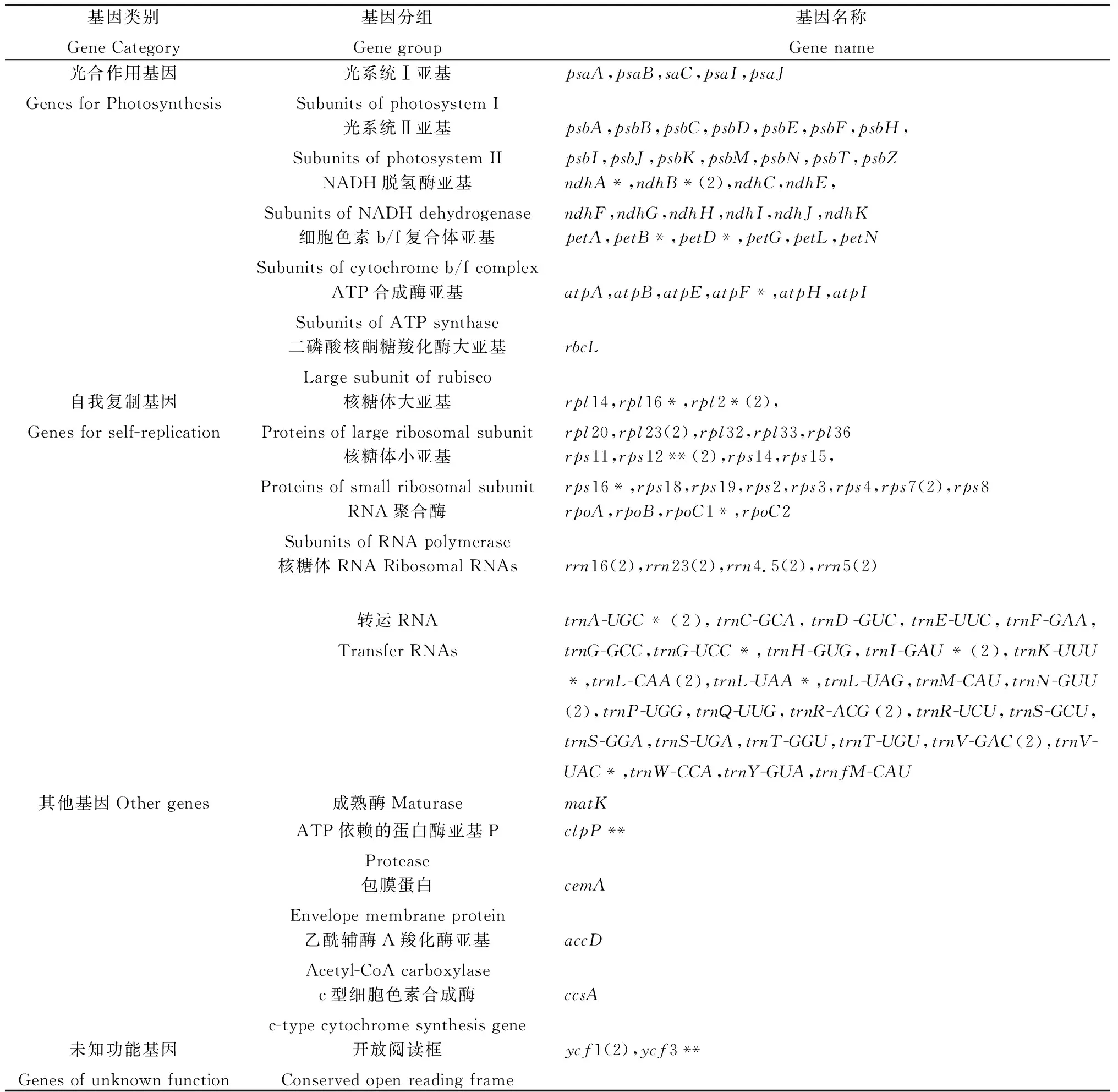
表1 直立型半野生大豆(G. gracilis)cpDNA注釋信息Table 1 Gene annotation of the chloroplast genome of the erect semi-wild soybean (Glycine gracilis)
2.2 密碼子偏好性
材料cpDNA密碼子偏好性等分析結(jié)果如圖2所示,可見(jiàn)編碼亮氨酸(L)、精氨酸(R)、絲氨酸(S)的密碼子數(shù)量較多,編碼色氨酸(W)的密碼子數(shù)量較少。RSCU值大于1的密碼子共有31個(gè),其中29個(gè)以A/U結(jié)尾,僅有2個(gè)以G/C結(jié)尾。
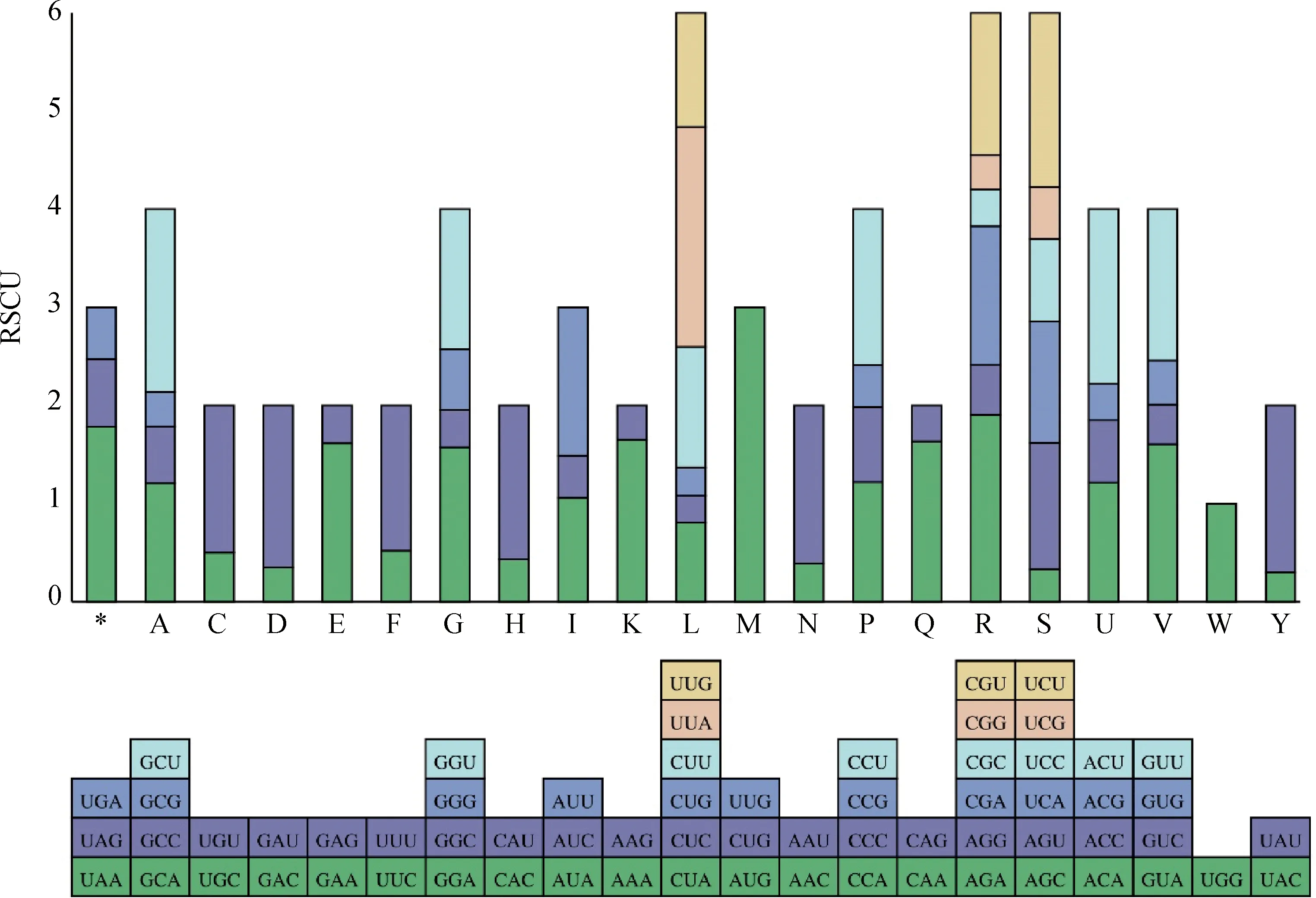
圖2 直立型半野生大豆(G.gracilis)葉綠體基因組各氨基酸的RSCU分析Fig.2 RSCU analysis of each amino acid in the erect semi-wild soybean (G. gracilis)
2.3 簡(jiǎn)單重復(fù)序列分析
從材料cpDNA中共鑒定出5種不同類型的SSR位點(diǎn)87個(gè)(表2)。其中單核苷酸重復(fù)序列最多,共55個(gè),分為A(31SSRs),T(23SSRs),C(1SSRs)三種類型;其次是雙核苷酸重復(fù)序列19個(gè),包含AT/TA兩種類型。此外,鑒定出三核苷酸重復(fù)序列4個(gè)、四核苷酸重復(fù)序列7個(gè)、五核苷酸重復(fù)序列2個(gè)。絕大多數(shù)SSR分布于大單拷貝區(qū),其余分布于小單拷貝區(qū)和反向重復(fù)序列a(表2)。
2.4 野生大豆及其近緣種系統(tǒng)發(fā)育分析
根據(jù)已公布的所有大豆屬材料cpDNA構(gòu)建系統(tǒng)發(fā)育樹(shù)。結(jié)果表明,大豆屬9個(gè)物種構(gòu)成一個(gè)單系類群,靴帶支持率(bootstrap)高達(dá)100%(見(jiàn)圖3)。該類群進(jìn)一步又可劃分為兩類,即一年生的半野生大豆G.gracilis.、栽培大豆G.max和野大豆G.soja與被測(cè)直立型半野生大豆組成一類,多年生大豆G.falcata,G.canescens,G.syndetika,G.dolichocarpa等組成一類。該結(jié)果與目前系統(tǒng)分類學(xué)對(duì)大豆屬植物的分類相符。
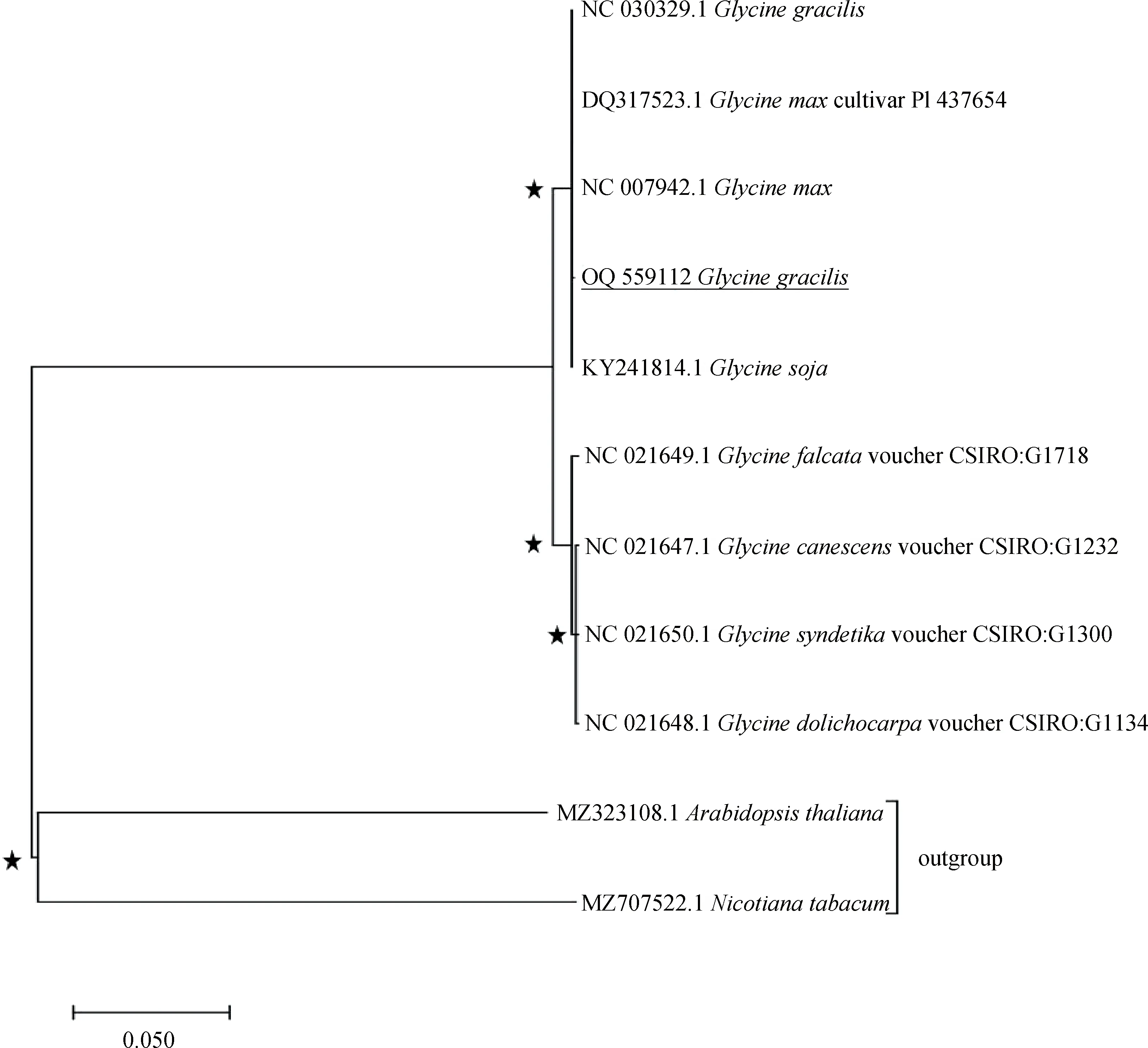
圖3 基于cpDNA序列構(gòu)建的大豆屬系統(tǒng)發(fā)育樹(shù)Fig.3 Phylogenetic tree constructed based on the DNA sequence in Glycine chloroplast genomes注:“★”代表分支節(jié)點(diǎn)的靴帶支持率為100%Note:“★”represents the bootstrap support rate of 100% on the branch
3 討論
葉綠體DNA與線粒體DNA一起被稱為細(xì)胞質(zhì)基因(Cytoplasmic genome),有遺傳多樣性和單親遺傳限制,異交重組等特點(diǎn),是植物系統(tǒng)發(fā)育研究、作物馴化追溯、作物改良和基因工程研究的重要對(duì)象[9,11,20-24]。隨著測(cè)序技術(shù)水平提高,測(cè)序成本不斷降低,植物cpDNA研究開(kāi)始興起。據(jù)報(bào)道,植物cpDNA長(zhǎng)度通常在120~180 kb,編碼基因一般在 100~130 個(gè),其中蛋白編碼基因多至 80 個(gè)、tRNA編碼基因30~32個(gè),rRNA編碼基因數(shù)穩(wěn)定,常有4種[25]。本文試驗(yàn)材料cpDNA大小、編碼基因數(shù)目和種類等與前人研究結(jié)果一致,符合cpDNA高度保守性的特性。
植物翻譯蛋白時(shí)并不平均地使用同義密碼子,某一或幾種特定密碼子使用頻率高于其他同義密碼子,這種現(xiàn)象即為密碼子偏好性。RSCU常被用作衡量密碼子偏好性,可在一定程度上反映基因乃至物種起源及進(jìn)化方式,并對(duì)基因功能及其編碼蛋白表達(dá)有一定影響[26]。當(dāng)某一密碼子RSCU值大于1,則表明其使用頻率相對(duì)較高。本研究中密碼子偏好性分析結(jié)果表明,RSCU值大于1的同義密碼子共有31個(gè),其中以A/T結(jié)尾的有29個(gè),與扁蓿豆、紫花苜蓿等豆科植物密碼子偏好性一致[27-28]。簡(jiǎn)單重復(fù)序列(SSR)又稱微衛(wèi)星,是整個(gè)基因組中1~6 bp的重復(fù)序列,多態(tài)性高、分布廣泛,是高等真核生物基因組的重要組成部分。又因?yàn)閏pDNA結(jié)構(gòu)簡(jiǎn)單、相對(duì)保守,cpSSR成為在遺傳分析、物種鑒定、群體遺傳多態(tài)性等研究中的重要工具[29]。在本研究中共鑒定出單核苷酸、雙核苷酸、三核苷酸、四核苷酸、五核苷酸共5種不同類型的SSR位點(diǎn)87個(gè),重復(fù)序列數(shù)及類型與已報(bào)道的其他被子植物cpDNA的SSRs序列構(gòu)成基本一致[30],同時(shí)印證了cpSSRs主要由短的poly A、poly T而非C或G的重復(fù)串聯(lián)構(gòu)成[31]。cpDNA尤其對(duì)近緣物種,能有效進(jìn)行鑒定和系統(tǒng)親緣關(guān)系分析[32]。本文基于cpDNA序列對(duì)大豆屬9個(gè)物種構(gòu)建系統(tǒng)發(fā)育樹(shù),結(jié)果與目前系統(tǒng)分類學(xué)對(duì)大豆屬植物的分類相符合。但半野生大豆在soja亞屬中的系統(tǒng)分類地位尚不能完全說(shuō)明。現(xiàn)已公布soja亞屬cpDNA數(shù)據(jù)僅有9個(gè),或許隨著數(shù)據(jù)量的增加可以從cpDNA角度解釋半野生大豆在soja亞屬中的分類地位。
4 結(jié)論
直立型半野生大豆cpDNA全長(zhǎng)148 320 bp,典型四分體結(jié)構(gòu),編碼基因108個(gè),其中蛋白編碼基因66個(gè)、tRNA基因29個(gè)、rRNA基因4個(gè)。基因組序列上共檢測(cè)出單核苷酸、雙核苷酸、三核苷酸、四核苷酸、五核苷酸,五種類型SSR位點(diǎn)87個(gè)。在系統(tǒng)發(fā)育上,直立型半野生大豆與已公布的普通型一年生半野生大豆(G.gracilis)、大豆(G.max)及野大豆(G.soja)親緣關(guān)系較近,但其在soja亞屬中的分類地位還有待進(jìn)一步研究。本研究對(duì)直立型野大豆cpDNA進(jìn)行了初步分析,為半野生大豆細(xì)胞質(zhì)多樣性、半野生大豆系統(tǒng)分類、分子育種及種植利用提供依據(jù),也為soja亞屬系統(tǒng)進(jìn)化和遺傳研究提供了重要的數(shù)據(jù)資源。
猜你喜歡
農(nóng)業(yè)科技通訊(2023年1期)2023-02-12 07:09:18
今日農(nóng)業(yè)(2022年16期)2022-11-09 23:18:44
中國(guó)化肥信息(2022年7期)2022-08-31 01:29:28
中國(guó)化肥信息(2022年5期)2022-08-30 01:58:26
今日農(nóng)業(yè)(2021年20期)2021-11-26 01:23:56
今日農(nóng)業(yè)(2021年14期)2021-10-14 08:35:34
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
農(nóng)產(chǎn)品市場(chǎng)周刊(2017年4期)2017-03-03 19:40:05
兒童故事畫(huà)報(bào)·智力大王(2015年10期)2016-01-27 01:01:35
讀寫(xiě)算(中)(2015年10期)2015-11-07 07:24:12
