不同林齡華山松針葉氮磷生態(tài)化學(xué)計(jì)量及再吸收特征
2023-12-02 01:34:24何斌李青李望軍鄒順白曉龍薛曉輝
江蘇農(nóng)業(yè)科學(xué) 2023年20期
何斌 李青 李望軍 鄒順 白曉龍 薛曉輝
摘要:分析不同林齡華山松葉片氮(N)和磷(P)再吸收特征及其與葉片、土壤養(yǎng)分指標(biāo)的關(guān)系,探討華山松生長(zhǎng)過程對(duì)生境的適應(yīng)性及養(yǎng)分利用機(jī)制。以10、16、22、47年生華山松人工林為研究對(duì)象,測(cè)定0~20 cm土層土壤、新鮮葉和凋落葉的養(yǎng)分含量,計(jì)算并分析葉片N、P再吸收率及其與葉片、土壤養(yǎng)分含量和化學(xué)計(jì)量特征的關(guān)系。結(jié)果表明,不同林齡華山松鮮葉N、P含量差異顯著(P<0.05),隨林齡增加均呈先升高后降低趨勢(shì);不同林齡華山松凋落葉N含量差異不顯著,P含量差異顯著(P<0.05),隨林齡的增加均未表現(xiàn)出明顯的規(guī)律性。不同林齡華山松新鮮葉 N ∶P 比差異不顯著(P<0.05),隨林齡增加呈先降低后增加趨勢(shì)。不同林齡華山松N、P的再吸收率存在顯著差異(P<0.05),N再吸收率明顯小于P再吸收率。N再吸收率與凋落葉N含量呈顯著負(fù)相關(guān)(P<0.05),與土壤有機(jī)碳(SOC)呈極顯著負(fù)相關(guān)(P<0.01);P再吸收率與凋落葉P含量、土壤C ∶N和C ∶P呈顯著負(fù)相關(guān)(P<0.05),與土壤速效磷含量呈顯著正相關(guān)(P<0.05),與土壤N ∶P呈極顯著負(fù)相關(guān)(P<0.01)。該地區(qū)華山松的生長(zhǎng)受氮素限制更加嚴(yán)重。N、P再吸收率均低于全球尺度上陸地植物的養(yǎng)分重吸收率,但P再吸收效率相對(duì)較高,說明華山松養(yǎng)分再吸收率不僅與生境條件有關(guān),還與營(yíng)養(yǎng)元素的獲取途徑有關(guān)。建議在喀斯特地區(qū)華山松人工林撫育管理時(shí)適當(dāng)施加氮肥,以促進(jìn)華山松的生長(zhǎng)發(fā)育。
關(guān)鍵詞:華山松;林齡;養(yǎng)分再吸收;養(yǎng)分含量;化學(xué)計(jì)量比
中圖分類號(hào):S718.45文獻(xiàn)標(biāo)志碼:A
文章編號(hào):1002-1302(2023)20-0137-07
植物養(yǎng)分再吸收(nutrient resorption)是指植物組織或器官在脫落之前將其部分營(yíng)養(yǎng)成分轉(zhuǎn)移運(yùn)輸?shù)狡渌L(zhǎng)組織供其重復(fù)利用的過程[1]。該過程不僅可以延長(zhǎng)養(yǎng)分在植物體內(nèi)的保留時(shí)間、提高養(yǎng)分利用效率、提供植物持續(xù)生長(zhǎng)所需的大部分養(yǎng)分[2-3],是植物增強(qiáng)競(jìng)爭(zhēng)力、提高生產(chǎn)力的重要策略之一[4-5];還可以降低植物對(duì)土壤養(yǎng)分的依賴、減緩生態(tài)系統(tǒng)養(yǎng)分損失[4,6],是植物對(duì)養(yǎng)分貧瘠環(huán)境的一種適應(yīng)進(jìn)化機(jī)制[7-9]。養(yǎng)分再吸收是植物獲取養(yǎng)分的重要途徑之一,在很大程度上會(huì)影響植物的養(yǎng)分循環(huán)和林分生產(chǎn)力[9]。Drenovsky等認(rèn)為,在植物的眾多器官中,葉片作為光合作用的主要器官,在養(yǎng)分轉(zhuǎn)移、運(yùn)輸和儲(chǔ)存中的貢獻(xiàn)最大[10]。Vergutz等認(rèn)為,全球植物N、P的再吸收率分別約為62.1%、64.9%[11]。因此,研究植物葉片養(yǎng)分再吸收特征有助于揭示植物對(duì)養(yǎng)分的利用能力和環(huán)境的適應(yīng)機(jī)制[12]。N、P作為植物生長(zhǎng)發(fā)育所必需的營(yíng)養(yǎng)元素,在植物生理代謝、養(yǎng)分循環(huán)和生態(tài)系統(tǒng)功能等發(fā)面起著重要作用[13]。植物葉片N、P的生態(tài)化學(xué)計(jì)量特征可揭示植物養(yǎng)分相對(duì)含量[14]和N、P限制格局以及再吸收能力[15],是研究植物養(yǎng)分限制和生態(tài)系統(tǒng)元素平衡的重要手段。有研究表明,植物的生態(tài)化學(xué)計(jì)量特征會(huì)受到生理過程需求、不同器官養(yǎng)分儲(chǔ)存和功能性分化差異的影響[16-17]。隨著林齡增加,森林生態(tài)系統(tǒng)的組成和內(nèi)部環(huán)境均會(huì)發(fā)生變化,導(dǎo)致養(yǎng)分分配格局的重組[18]。而養(yǎng)分再吸收與植物的養(yǎng)分需求和環(huán)境的養(yǎng)分供應(yīng)密切相關(guān)[19]。Gusewell等研究發(fā)現(xiàn),當(dāng)植物生長(zhǎng)受到某種元素限制時(shí),該元素的再吸收率會(huì)相對(duì)較高[20-21]。Vitousek等的研究表明,葉片和凋落物的化學(xué)計(jì)量比會(huì)影響?zhàn)B分的再吸收效率[22]。然而,關(guān)于養(yǎng)分再吸收與植物葉片養(yǎng)分化學(xué)計(jì)量特征之間的關(guān)系尚無定論,潛在的響應(yīng)機(jī)制尚不清楚。目前,對(duì)于養(yǎng)分再吸收的研究主要集中在與植物生活型[23-24]、葉片壽命長(zhǎng)短[25]、土壤養(yǎng)分含量[26-27]、群落演替[28]、不同種類[5,29-30]等關(guān)系方面,對(duì)不同林齡樹種養(yǎng)分再吸收效率的研究報(bào)道較少。因此,研究不同林齡植物葉片N、P再吸收規(guī)律及其與養(yǎng)分化學(xué)計(jì)量特征的關(guān)系,可以更好地闡釋人工林植被的養(yǎng)分利用機(jī)制及其環(huán)境適應(yīng)策略,對(duì)不同造林樹種制定有針對(duì)性、科學(xué)合理的管護(hù)措施具有重要意義。以貴州省為中心的中國(guó)西南喀斯特地區(qū)是全球喀斯特面積最大的集中分布區(qū),生態(tài)系統(tǒng)脆弱、土地貧瘠、土層薄且不連續(xù),石漠化現(xiàn)象嚴(yán)重[31]。改善喀斯特地區(qū)生態(tài)環(huán)境的根本途徑是恢復(fù)植被,而不同恢復(fù)階段生境的養(yǎng)分供應(yīng)能力和植物吸收利用養(yǎng)分的能力不同[32]。在喀斯特石漠化地區(qū)開展植物生態(tài)化學(xué)計(jì)量研究,有助于深入了解植物對(duì)貧瘠環(huán)境的生長(zhǎng)適應(yīng)策略以及喀斯特生態(tài)系統(tǒng)的養(yǎng)分循環(huán)[33]。此前有學(xué)者對(duì)喀斯特森林生態(tài)系統(tǒng)的植物、凋落物、土壤和微生物系統(tǒng)C、N、P生態(tài)化學(xué)計(jì)量特征進(jìn)行了大量研究,并指出不同植被類型群落可能受到N或P養(yǎng)分限制,然而面對(duì)養(yǎng)分限制條件,不同植物群落和生長(zhǎng)階段樹種的養(yǎng)分再吸收特征仍然不夠明確。因此,本研究選取貴州省西北部典型的植被恢復(fù)樹種華山松,分析其不同林齡階段針葉N、P化學(xué)計(jì)量比及再吸收特征,闡明華山松生長(zhǎng)過程中對(duì)養(yǎng)分限制環(huán)境的適應(yīng)性,以期為揭示喀斯特地區(qū)植物的養(yǎng)分循環(huán)、生態(tài)策略及環(huán)境適應(yīng)機(jī)制提供科學(xué)依據(jù)。
1 研究區(qū)概況與研究方法
1.1 研究區(qū)概況
研究區(qū)位于貴州省西北部的畢節(jié)市,地勢(shì)西高東低,山巒重疊,地貌形態(tài)多樣,有高原、山地、盆地、谷地、平壩、峰叢等,以沉積巖為主,占全市總面積的92.81%。海拔相對(duì)高差大,平均海拔1 600 m。屬亞熱帶高原季風(fēng)氣候,垂直氣候變化明顯,年平均溫度10~15 ℃,年日照時(shí)數(shù)1 096~1 769 h,降水量較充沛,年均降水量849~1 399 mm,無霜期 245~290 d。土壤主要為黃棕壤、石灰土和石質(zhì)土。20世紀(jì)60年代,該區(qū)域原有植被遭到嚴(yán)重破壞,之后通過人工造林,營(yíng)造了大量的華山松(P. armandii)、云南松(P. yunnanensis)和杉木(Cunninghamia lanceolata)人工林。
1.2 樣地設(shè)置與取樣方法
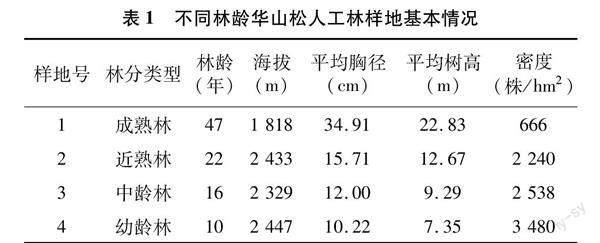
2021年5月采用“空間替代時(shí)間”的方法,在研究區(qū)內(nèi)選擇地形地貌、坡度等立地條件相似的10、16、22、47年生華山松人工林作為固定監(jiān)測(cè)樣地,在每個(gè)林齡內(nèi)隨機(jī)設(shè)置3塊20 m×20 m的重復(fù)樣方,樣方間距100 m,共12塊。調(diào)查樣方內(nèi)華山松的樹高、胸徑、冠幅等指標(biāo),同時(shí)記錄樣地的基本信息(表1)。
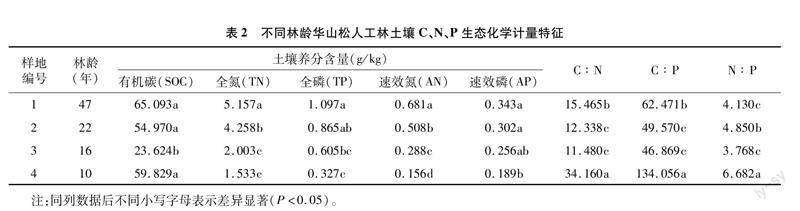
2021年8月,在每個(gè)樣方內(nèi)選取5株標(biāo)準(zhǔn)木,從東、西、南、北4個(gè)方位用高枝剪在每株標(biāo)準(zhǔn)木樹冠的上部、中部、下部外側(cè)采集枝條,從每根枝條上摘取健康、完整的新鮮成熟針葉,采集50 g/株,同一樣方內(nèi)的針葉混合均勻后裝入牛皮紙袋,作為1個(gè)樣品帶回實(shí)驗(yàn)室。新鮮針葉在90 ℃殺青30 min,65 ℃ 烘干至恒質(zhì)量,粉碎后過100目篩備用。同時(shí),在每個(gè)樣方內(nèi)用土鉆在0~20 cm土層取樣,按“S”形設(shè)置5個(gè)采樣點(diǎn),將5個(gè)土樣混合為1個(gè)土壤樣品裝入布袋帶回實(shí)驗(yàn)室。挑除石礫和植物根系殘?bào)w后,將土壤樣品置于陰涼處自然風(fēng)干,用四分法取土、研磨、過100目篩網(wǎng),用于測(cè)定土壤養(yǎng)分含量(表2)。
2021年10月初,在每株標(biāo)準(zhǔn)林木下布設(shè)4個(gè) 1 m×1 m的凋落物收集框,收集新近自然脫落而未分解的枯落物,月底將收集的枯落物去除其他雜質(zhì),將凋落葉混合均勻作為1個(gè)樣品裝入牛皮紙袋,帶回實(shí)驗(yàn)室。凋落葉在65 ℃烘干至恒質(zhì)量,粉碎后過100目篩備用。
1.3 樣品測(cè)定
針葉和土壤有機(jī)碳含量的測(cè)定采用重鉻酸鉀-外加熱法;針葉經(jīng)過H2SO4-H2O2加熱消煮后,全氮含量的測(cè)定采用凱氏定氮法,全磷含量的測(cè)定采用鉬銻抗比色法;土壤全氮含量的測(cè)定采用凱氏定氮法,速效氮含量的測(cè)定采用堿解擴(kuò)散法,全磷和速效磷含量的測(cè)定采用H2SO4-HClO4-鉬銻抗比色法。
1.4 數(shù)據(jù)處理
參考Milla等的研究方法[34],植物葉片養(yǎng)分再吸收率計(jì)算公式為
NuRE=【1-(w1/w2)】×100%。
式中:NuRE表示葉片N或P的再吸收率;w1表示凋落葉N含量或P含量,g/kg;w2表示新鮮成熟葉N或P含量,g/kg。
應(yīng)用軟件SPSS 22.0中的單因素方差分析(one-way analysis of variance)和最小顯著性差異法(least significant difference,LSD)計(jì)算分析不同林齡間各指標(biāo)的差異;應(yīng)用Pearson分析法分析養(yǎng)分再吸收率與新鮮針葉、凋落葉和土壤中C、N、P生態(tài)化學(xué)計(jì)量特征之間的相關(guān)性關(guān)系。
2 結(jié)果與分析
2.1 不同林齡華山松鮮葉和凋落葉N、P生態(tài)化學(xué)計(jì)量特征
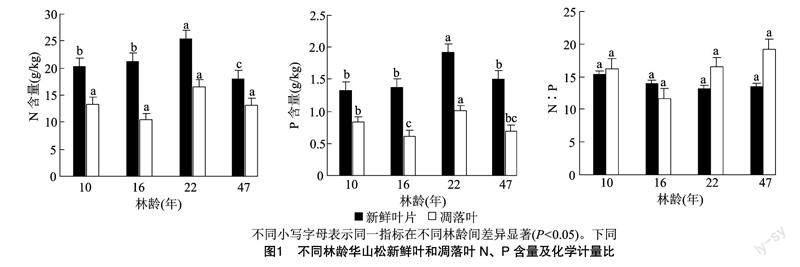
由圖1可知,不同林齡華山松新鮮葉N、P含量差異顯著(P<0.05),隨林齡增加均呈先升高后降低的趨勢(shì);不同林齡華山松新鮮葉N ∶P差異不顯著(P<0.05),隨林齡增加呈先降低后增加趨勢(shì)。新鮮葉N含量為18.086~25.464 g/kg,方差分析結(jié)果表明,22年生林分顯著大于其他林分,47年生林分顯著小于16年生和10年生(P<0.05),表現(xiàn)為22年生>16年生>10年生>47年生;P含量為1.328~1.923 g/kg,方差分析結(jié)果表明,22年生林分顯著大于其他林分(P<0.05),表現(xiàn)為22年生>47年生>16年生>10年生;N ∶P為13.280~15.347,表現(xiàn)為10年生>16年生>47年生>22年生。
不同林齡華山松凋落葉N含量差異不顯著,P含量差異顯著(P<0.05),隨林齡的增加未表現(xiàn)出明顯的規(guī)律性;不同林齡華山松凋落葉N ∶P差異不顯著(P<0.05),隨林齡增加呈先降低后增加趨勢(shì)。凋落葉N含量為10.379~16.598 g/kg,表現(xiàn)為22年生>10年生>47年生>16年生;P含量為0.611~1.009 g/kg,方差分析結(jié)果表明,22年生林分顯著大于其他林分,10年生林分顯著大于16 年生(P<0.05),表現(xiàn)為22年生>10年生>47年生>16年生;N ∶P為11.603~19.121,表現(xiàn)為47年生>22年生>10年生>16年生。
2.2 不同林齡華山松針葉N、P再吸收率
由圖2可知,不同林齡華山松N、P的再吸收率存在顯著差異(P<0.05),N的再吸收率明顯小于P的再吸收率。N的再吸收率為27.065%~50.655%,方差分析結(jié)果表明,16年生林分顯著大于其他林分(P<0.05),表現(xiàn)為16年生>22年生>10年生>47年生,隨林齡增加呈先升高后降低的趨勢(shì)。 P的再吸收率為[JP+1]37.348%~55.482% 方差分析結(jié)果表明,16年生和47年生林分顯著大于10年生(P<0.05),表現(xiàn)為16年生>47年生>22年生>10年生,隨林齡的增加未表現(xiàn)出明顯的規(guī)律性。
2.3 N、P再吸收率與針葉和土壤C、N、P化學(xué)計(jì)量特征的關(guān)系
由表3可知,華山松針葉N、P再吸收率與新鮮葉片和凋落葉的TN、TP含量以及C ∶N ∶P化學(xué)計(jì)量比均有一定的相關(guān)性。N再吸收率除與凋落葉TN含量呈顯著負(fù)相關(guān)外(P<0.05),與新鮮葉片和凋落葉其指標(biāo)顯著性均不顯著(P<0.05);而P再吸收率除與凋落葉TP含量呈顯著負(fù)相關(guān)外(P<0.05),與新鮮葉片和凋落葉其他指標(biāo)均不顯著(P<0.05)。
由表4可知,華山松針葉N、P再吸收率與土壤養(yǎng)分和化學(xué)計(jì)量比存在一定的相關(guān)性。N再吸收率與土壤養(yǎng)分各指標(biāo)均呈負(fù)相關(guān)性,但只與土壤有機(jī)碳(SOC)含量呈極顯著負(fù)相關(guān)(P<0.01);P再吸收率與土壤N ∶P呈極顯著負(fù)相關(guān)(P<0.01),與土壤C ∶N和C ∶P呈顯著負(fù)相關(guān)(P<0.05),與土壤速效磷含量呈顯著正相關(guān)(P<0.05),與土壤養(yǎng)分其他指標(biāo)相關(guān)性不顯著(P<0.05)。
3 討論
3.1 林齡對(duì)華山松針葉N、P生態(tài)化學(xué)計(jì)量特征的影響
葉片是植物代謝活動(dòng)最活躍的器官,其養(yǎng)分含量和計(jì)量比能反映植物生長(zhǎng)過程中的營(yíng)養(yǎng)狀況[35],而植物營(yíng)養(yǎng)狀況不僅受到多種環(huán)境因素(如土壤養(yǎng)分、水分供應(yīng)等)的綜合影響[36],還受到植物種類和生長(zhǎng)階段的影響[37]。本研究不同林齡華山松新鮮葉片N、P含量存在顯著差異,隨林齡增加均呈先升高后降低的趨勢(shì),22年生華山松針葉N、P含量最高。根據(jù)植物生長(zhǎng)策略,植物在接近成熟之前處于生長(zhǎng)旺盛期,具有較快的生長(zhǎng)速率,需要更多的rRNA來滿足蛋白質(zhì)的合成,導(dǎo)致葉片中N含量升高[38],而rRNA又是植物的主要磷庫(kù)之一[18],故22年生華山松針葉N、P含量顯著大于10、16、47年生林分華山松針葉N、P含量。成熟林階段,林下枯落物和微生物相對(duì)豐富,能為植物提供充足的營(yíng)養(yǎng)物質(zhì),根據(jù)自身的生長(zhǎng)需求,華山松選擇性吸收各種養(yǎng)分,因代謝增強(qiáng)積累的較多光合產(chǎn)物使各種營(yíng)養(yǎng)元素得到稀釋[39],導(dǎo)致47年生華山松針葉N、P含量的下降。
與新鮮葉片相比,不同林齡華山松凋落葉N、P含量明顯下降,這反映植物對(duì)N、P元素的再吸收特征。不同林齡華山松凋落葉N含量差異不顯著,P含量差異顯著,均表現(xiàn)為22年生>10年生>47年生>16年生。這與新鮮葉片中N、P含量達(dá)到最大的年齡一致,說明凋落葉N、P秉承了植物葉的特性;而16年生華山松凋落葉N、P含量最小,可能是由于植物對(duì)N、P的再吸收所致,16年生華山松對(duì)新鮮針葉N、P具有最高的再吸收率(圖2)。
葉片N ∶P常作為評(píng)價(jià)植物養(yǎng)分受限狀況的指標(biāo)[27]。Koerselman等認(rèn)為,葉片N ∶P<14受N限制,N ∶P>16受P限制,N ∶P在14~16之間受N、P共同限制[40-41]。本研究中的華山松N ∶P在13.280~15.347范圍內(nèi),僅10年生華山松針葉 N ∶P(15.347)>14,其他各林齡華山松針葉的 N ∶P 均小于14,說明研究區(qū)內(nèi)華山松生長(zhǎng)受氮素限制較嚴(yán)重,這與皮發(fā)劍等在喀斯特地區(qū)森林的研究結(jié)果[32,42]一致。隨著林齡的增加,華山松新鮮葉片N ∶P先降低后增加,這可能與華山松為適應(yīng)土壤N、P可利用性變化而改變生長(zhǎng)策略有關(guān)[43]。雖然不同林齡華山松針葉N、P含量有顯著差異,但是N ∶P差異不顯著、保持相對(duì)穩(wěn)定,說明華山松自身具有調(diào)節(jié)元素需求與養(yǎng)分吸收平衡的能力,進(jìn)一步驗(yàn)證了植物的內(nèi)穩(wěn)態(tài)維持機(jī)制[44]。除16年生林分外,華山松凋落葉N ∶P均高于新鮮葉,這是因?yàn)镻的再吸收率高于N的再吸收率。趙其國(guó)等認(rèn)為,凋落葉N ∶P影響凋落物分解和養(yǎng)分歸還速率,N ∶P越低,凋落物分解越快[45]。隨著林齡的增加,華山松凋落葉 N ∶P 先降低后增加,但是差異不顯著,可能是因?yàn)槿A山松近熟林和成熟林通過降低凋落葉的N ∶P加速凋落物的分解,補(bǔ)充土壤中的養(yǎng)分來滿足植物的生長(zhǎng)所需。
3.2 林齡對(duì)華山松針葉N、P再吸收率的影響
隨著植物的生長(zhǎng)發(fā)育,植物對(duì)養(yǎng)分的需求、敏感程度和吸收能力會(huì)發(fā)生變化,進(jìn)而導(dǎo)致養(yǎng)分再吸收率表現(xiàn)出不同的規(guī)律[4]。本研究隨林齡的增加,華山松針葉N再吸收率先升高后降低(圖2),與李榮華等對(duì)不同林齡馬尾松的研究結(jié)果[46]一致,這可能與樹木的生長(zhǎng)規(guī)律有關(guān)。盡管華山松針葉P再吸收率隨林齡的增加未表現(xiàn)出明顯的變化趨勢(shì),但是P的再吸收率仍然在22年生達(dá)到最大,這與N的再吸收率一致。這是因?yàn)?2年生華山松生長(zhǎng)旺盛,養(yǎng)分需求量大,促進(jìn)了N、P的再吸收。不同林齡華山松針葉中N、P的再吸收率分別為27.065%~50.655%、37.348%~55.482%,均低于全球陸生植物葉片N、P的再吸收率62.10%、64.90%[11],可能與樹種間的遺傳差異以及喀斯特地區(qū)特殊、復(fù)雜的立地條件有關(guān),同時(shí)本研究未考慮針葉凋落后的質(zhì)量損失也會(huì)導(dǎo)致計(jì)算結(jié)果偏低[11]。
Killingbeck認(rèn)為,當(dāng)枯落葉中N、P含量分別小于7、0.5 g/kg時(shí),植物對(duì)N、P完全再吸收;當(dāng)枯落葉中N、P含量分別大于10、0.8 g/kg時(shí),植物對(duì)N、P再吸收程度較低,為不完全吸收,養(yǎng)分含量越低,植物再吸收程度越高[47]。本研究不同林齡華山松凋落葉N含量為10.379~16.598 g/kg,P含量為0.611~1.009 g/kg,華山松針葉的N、P含量均表現(xiàn)為不完全吸收,表明華山松養(yǎng)分轉(zhuǎn)移較差。但是華山松凋落葉中P含量更低,說明華山松對(duì)P再吸收較多。盡管華山松生長(zhǎng)更大程度上受N限制,但是P再吸收率高于N再吸收率,與Han等提出的“相對(duì)再吸收假說”相矛盾[48]。這可能是由于高等植物的養(yǎng)分再吸收機(jī)制非常復(fù)雜,不僅與物種的養(yǎng)分需求和植物對(duì)養(yǎng)分組分的權(quán)衡有關(guān)[49],還可能會(huì)受到庫(kù)容大小[50]、葉片脫落機(jī)制[51]、韌皮部轉(zhuǎn)運(yùn)率[52]等許多內(nèi)稟遺傳特性的影響,受養(yǎng)分限制時(shí)不一定必須通過提高對(duì)該養(yǎng)分的再吸收方式來適應(yīng)環(huán)境[53-54],還可以采取一些其他途徑來適應(yīng)環(huán)境。Wright等認(rèn)為,在養(yǎng)分貧瘠的環(huán)境中,植物主要通過延長(zhǎng)葉片壽命和降低養(yǎng)分濃度來適應(yīng)養(yǎng)分限制,而不是依賴高的養(yǎng)分再吸收率[25]。
3.3 華山松葉片N、P再吸收率與葉片和土壤養(yǎng)分的關(guān)系
雖然養(yǎng)分再吸收是植物進(jìn)化過程中形成的固有特性[47],但是Vergutz等認(rèn)為,植物養(yǎng)分再吸收較復(fù)雜,除受自身的遺傳特性影響外,還可能受到成熟葉片、凋落物和土壤養(yǎng)分單獨(dú)或共同的影響[11,45]。目前,有關(guān)養(yǎng)分再吸收與葉片養(yǎng)分含量關(guān)系的研究結(jié)論還不一致。孫書存等認(rèn)為,成熟葉養(yǎng)分濃度升高會(huì)促進(jìn)養(yǎng)分的再吸收[55];Vergutz等認(rèn)為,成熟葉養(yǎng)分含量升高會(huì)降低N、P再吸收率[11]。而Aerts認(rèn)為,養(yǎng)分再吸收率與成熟葉片養(yǎng)分含量沒有顯著的相關(guān)性,與凋落葉養(yǎng)分含量呈顯著負(fù)相關(guān)[1]。本研究華山松N再吸收率與凋落葉N含量呈顯著負(fù)相關(guān)(P<0.05),P再吸收率與凋落葉P含量呈顯著負(fù)相關(guān)(P<0.05),說明當(dāng)養(yǎng)分不足時(shí),華山松會(huì)通過提高成熟葉片養(yǎng)分的再吸收率和降低凋落葉中養(yǎng)分含量來減少養(yǎng)分損失,重新利用體內(nèi)的養(yǎng)分,這也是植物提高養(yǎng)分利用率的重要策略之一[56]。華山松N、P再吸收率與新鮮葉和凋落葉中C ∶N ∶P的化學(xué)計(jì)量比均不顯著,這與邱嶺軍等的研究結(jié)論[57-58]存在差異。這可能是由不同植物、立地環(huán)境條件的影響和研究方法的差異造成的[59]。
Tang等認(rèn)為,養(yǎng)分再吸收作為植物應(yīng)對(duì)環(huán)境養(yǎng)分脅迫的一種策略,會(huì)受到環(huán)境養(yǎng)分供應(yīng)的影響[60]。Achat等認(rèn)為,隨著土壤養(yǎng)分含量的增加養(yǎng)分再吸收率會(huì)降低[61]。但本研究發(fā)現(xiàn)華山松P再吸收率與土壤速效磷含量呈顯著正相關(guān),這與龍靖等對(duì)西沙熱帶珊瑚島喬木植物養(yǎng)分再吸收特征研究的結(jié)果[62]類似。這可能與土壤速效磷的利用策略會(huì)影響華山松針葉中磷的供應(yīng),從而影響凋落葉中P的再吸收策略有關(guān)。華山松P再吸收與土壤 C ∶N ∶P 化學(xué)計(jì)量比呈顯著負(fù)相關(guān),比土壤養(yǎng)分對(duì)P再吸收率的影響更大,可能是由于化學(xué)計(jì)量比能更好地反映土壤養(yǎng)分的相對(duì)含量,土壤化學(xué)計(jì)量特征的變化能直接或間接驅(qū)動(dòng)植物葉片養(yǎng)分的再吸收策略。
4 結(jié)論
不同林齡華山松新鮮葉片和凋落葉N、P含量存在顯著差異,均在22年生林分達(dá)到最大值。隨著林齡增加,新鮮葉N、P含量表現(xiàn)為先上升后降低,凋落葉則未表現(xiàn)出明顯的規(guī)律性。華山松針葉的 N ∶P 在不同林齡間差異不顯著,且均小于14年生(10年生除外),可見該地區(qū)華山松的生長(zhǎng)受氮素限制更嚴(yán)重。因此,在喀斯特地區(qū)華山松人工林撫育管理時(shí),應(yīng)適當(dāng)施加氮肥促進(jìn)華山松的生長(zhǎng)發(fā)育,提高其生產(chǎn)力。不同林齡華山松針葉N、P再吸收率均低于全球尺度上陸地植物的再吸收率,但P再吸收效率相對(duì)較高,表明華山松葉片養(yǎng)分再吸收率不僅與生境條件有關(guān),還與營(yíng)養(yǎng)元素的獲取途徑有關(guān)。此外,N、P再吸收率與新鮮葉片中N、P含量相關(guān)性不顯著,與凋落葉中N、P含量呈顯著負(fù)相關(guān),說明凋落葉中養(yǎng)分含量越低,養(yǎng)分再吸收率越高,可見華山松是通過降低凋落葉中的養(yǎng)分含量來適應(yīng)喀斯特地區(qū)貧瘠的土壤條件的。本研究?jī)H從葉片養(yǎng)分再吸收特征及其與葉片和土壤養(yǎng)分關(guān)系的角度進(jìn)行分析,在將來的研究中還要從植物生理學(xué)方面深入探究,進(jìn)一步揭示養(yǎng)分再吸收的內(nèi)在機(jī)理。
參考文獻(xiàn):
[1]Aerts R. Nutrient resorption from senescing leaves of perennials:are there general patterns?[J]. The Journal of Ecology,1996,84(4):597-608.
[2]Sollins P,Grier C C,McCorison F M,et al. The internal element cycles of an old-growth Douglas-fir ecosystem in western Oregon[J]. Ecological Monographs,1980,50(3):261-285.
[3]Meier C E,Grier C C,Cole D W.Below- and aboveground N and P use by Abies amabilis stands[J]. Ecology,1985,66(6):1928-1942.
[4]陸姣云,段兵紅,楊 梅,等. 植物葉片氮磷養(yǎng)分重吸收規(guī)律及其調(diào)控機(jī)制研究進(jìn)展[J]. 草業(yè)學(xué)報(bào),2018,27(4):178-188.
[5]劉宏偉,劉文丹,王 微,等. 重慶石灰?guī)r地區(qū)主要木本植物葉片性狀及養(yǎng)分再吸收特征[J]. 生態(tài)學(xué)報(bào),2015,35(12):4071-4080.
[6]Lye X T,Reed S C,Yu Q,et al. Nutrient resorption helps drive intra-specific coupling of foliar nitrogen and phosphorus under nutrient-enriched conditions[J]. Plant and Soil,2016,398(1):111-120.
[7]Aerts R. Nutrient use efficiency in evergreen and deciduous species from heathlands[J]. Oecologia,1990,84(3):391-397.
[8]Regina I S,Leonardi S,Rapp M. Foliar nutrient dynamics and nutrient-use efficiency in Castanea sativa coppice stands of southern Europe[J]. Forestry,2001,74(1):1-10.
[9]Reed S C,Townsend A R,Davidson E A,et al. Stoichiometric patterns in foliar nutrient resorption across multiple scales[J]. New Phytologist,2012,196(1):173-180.
[10]Drenovsky R E,Richards J H. Low leaf N and P resorption contributes to nutrient limitation in two desert shrubs[J]. Plant Ecology,2006,183(2):305-314.
[11]Vergutz L,Manzoni S,Porporato A,et al. Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants[J]. Ecological Monographs,2012,82(2):205-220.
[12]Liang D,Zhang J J,Zhang S T. Patterns of nitrogen resorption in functional groups in a Tibetan alpine meadow[J]. Folia Geobotanica,2015,50:267-274.
[13]劉 順,羅 達(dá),劉千里,等. 川西亞高山不同森林生態(tài)系統(tǒng)碳氮儲(chǔ)量及其分配格局[J]. 生態(tài)學(xué)報(bào),2017,37(4):1074-1083.
[14]Drenovsky R E,Koehler C E,Skelly K,et al. Potential and realized nutrient resorption in serpentine and non-serpentine chaparral shrubs and trees[J]. Oecologia,2013,171(1):39-50.
[15]Han W X,F(xiàn)ang J Y,Guo D L,et al. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China[J]. New Phytologist,2005,168(2):377-385.
[16]Minden V,Kleyer M. Internal and external regulation of plant organ stoichiometry[J]. Plant Biology,2014,16(5):897-907.
[17]封煥英,杜滿義,辛學(xué)兵,等. 華北石質(zhì)山地側(cè)柏人工林C、N、P生態(tài)化學(xué)計(jì)量特征的季節(jié)變化[J]. 生態(tài)學(xué)報(bào),2019,39(5):1572-1582.
[18]崔寧潔,劉小兵,張丹桔,等. 不同林齡馬尾松(Pinus massoniana)人工林碳氮磷分配格局及化學(xué)計(jì)量特征[J]. 生態(tài)環(huán)境學(xué)報(bào),2014,23(2):188-195.
[19]Pugnaire F I,Chapin F S Ⅲ. Controls over nutrient resorption from leaves of evergreen Mediterranean species[J]. Ecology,1993,74(1):124-129.
[20]Gusewell S. Nutrient resorption of wetland graminoids is related to the type of nutrient limitation[J]. Functional Ecology,2005,19(2):344-354.
[21]Han W X,Tang L Y,Chen Y H,et al. Relationship between the relative limitation and resorption efficiency of nitrogen vs phosphorus in woody plants[J]. PLoS One,2013,8(12):e83366.
[22]Vitousek P M,Turner D R,Parton W J,et al. Litter decomposition on the Mauna Loa environmental matrix,Hawaii:patterns,mechanisms,and models[J]. Ecology,1994,75(2):418-429.
[23]Brant A N,Chen H Y. Patterns and mechanisms of nutrient resorption in plants[J]. Critical Reviews in Plant Sciences,2015,34(5):471-486.
[24]江大龍,徐 俠,阮宏華. 植物養(yǎng)分重吸收及其影響研究進(jìn)展[J]. 南京林業(yè)大學(xué)學(xué)報(bào)(自然科學(xué)版),2017,41(1):183-188.
[25]Wright I J,Westoby M. Nutrient concentration,resorption and lifespan:leaf traits of Australian sclerophyll species[J]. Functional Ecology,2003,17(1):10-19.
[26]劉佳慶,王曉雨,郭 焱,等. 長(zhǎng)白山林線主要木本植物葉片養(yǎng)分的季節(jié)動(dòng)態(tài)及回收效率[J]. 生態(tài)學(xué)報(bào),2015,35(1):165-171.
[27]趙 瓊,曾德慧. 林木生長(zhǎng)氮磷限制的診斷方法研究進(jìn)展[J]. 生態(tài)學(xué)雜志,2009,28(1):122-128.
[28]Kazakou E,Garnier E,Navas M L,et al. Components of nutrient residence time and the leaf economics spectrum in species from Mediterranean old-fields differing in successional status[J]. Functional Ecology,2007,21(2):235-245.
[29]鄧 健,種玉潔,賈小敏,等. 黃土高原子午嶺林區(qū)典型樹種葉片N、P再吸收特征[J]. 生態(tài)學(xué)報(bào),2020,40(11):3698-3705.
[30]申 奧,朱教君,閆 濤,等. 遼東山區(qū)主要闊葉樹種葉片養(yǎng)分含量和再吸收對(duì)落葉時(shí)間的影響[J]. 植物生態(tài)學(xué)報(bào),2018,42(5):573-584.
[31]Jiang Z C,Lian Y Q,Qin X Q. Rocky desertification in Southwest China:impacts,causes,and restoration[J]. Earth-Science Reviews,2014,132:1-12.
[32]皮發(fā)劍,袁叢軍,喻理飛,等. 黔中天然次生林主要優(yōu)勢(shì)樹種葉片生態(tài)化學(xué)計(jì)量特征[J]. 生態(tài)環(huán)境學(xué)報(bào),2016,25(5):801-807.
[33]曾昭霞,王克林,劉孝利,等. 桂西北喀斯特森林植物-凋落物-土壤生態(tài)化學(xué)計(jì)量特征[J]. 植物生態(tài)學(xué)報(bào),2015,39(7):682-693.
[34]Milla R,Castro-Díez P,Maestro-Martínez M,et al. Does the gradualness of leaf shedding govern nutrient resorption from senescing leaves in Mediterranean woody plants?[J]. Plant and Soil,2005,278(1):303-313.
[35]羅緒強(qiáng),張桂玲,杜雪蓮,等. 茂蘭喀斯特森林常見鈣生植物葉片元素含量及其化學(xué)計(jì)量學(xué)特征[J]. 生態(tài)環(huán)境學(xué)報(bào),2014,23(7):1121-1129.
[36]Elser J J,F(xiàn)agan W F,Denno R F,et al. Nutritional constraints in terrestrial and freshwater food webs[J]. Nature,2000,408(6812):578-580.
[37]Schreeg L A,Santiago L S,Wright S J,et al. Stem,root,and older leaf N ∶P ratios are more responsive indicators of soil nutrient availability than new foliage[J]. Ecology,2014,95(8):2062-2068.
[38]Andersen T,Elser J J,Hessen D O. Stoichiometry and population dynamics[J]. Ecology Letters,2004,7(9):884-900.
[39]盤金文,郭其強(qiáng),孫學(xué)廣,等. 不同林齡馬尾松人工林碳、氮、磷、鉀養(yǎng)分含量及其生態(tài)化學(xué)計(jì)量特征[J]. 植物營(yíng)養(yǎng)與肥料學(xué)報(bào),2020,26(4):746-756.
[40]Koerselman W,Meuleman A F M. The vegetation N ∶P ratio:a new tool to detect the nature of nutrient limitation[J]. The Journal of Applied Ecology,1996,33(6):1441-1450.
[41]Wright I J,Reich P B,Westoby M,et al. The worldwide leaf economics spectrum[J]. Nature,2004,428(6985):821-827.
[42]曠遠(yuǎn)文,溫達(dá)志,閆俊華,等. 貴州普定喀斯特森林3種優(yōu)勢(shì)樹種葉片元素含量特征[J]. 應(yīng)用與環(huán)境生物學(xué)報(bào),2010,16(2):158-163.
[43]李雪峰,韓士杰,胡艷玲,等. 長(zhǎng)白山次生針闊混交林葉凋落物中有機(jī)物分解與碳、氮和磷釋放的關(guān)系[J]. 應(yīng)用生態(tài)學(xué)報(bào),2008,19(2):245-251.
[44]汪宗飛,鄭粉莉. 黃土高原子午嶺地區(qū)人工油松林碳氮磷生態(tài)化學(xué)計(jì)量特征[J]. 生態(tài)學(xué)報(bào),2018,38(19):6870-6880.
[45]趙其國(guó),王明珠,何園球. 我國(guó)熱帶亞熱帶森林凋落物及其對(duì)土壤的影響[J]. 土壤,1991,23(1):8-15.
[46]李榮華,汪思龍,王清奎. 不同林齡馬尾松針葉凋落前后養(yǎng)分含量及回收特征[J]. 應(yīng)用生態(tài)學(xué)報(bào),2008,19(7):1443-1447.
[47]Killingbeck K T. Nutrients in senesced leaves:keys to the search for potential resorption and resorption proficiency[J]. Ecology,1996,77(6):1716-1727.
[48]Han W X,Tang L Y,Chen Y H,et al. Relationship between the relative limitation and resorption efficiency of nitrogen vs phosphorus in woody plants[J]. PLoS One,2013,8(12):e83366.
[49]Wang Z N,Lu J Y,Yang H M,et al. Resorption of nitrogen,phosphorus and potassium from leaves of lucerne stands of different ages[J]. Plant and Soil,2014,383(1):301-312.
[50]Silla F,Escudero A. Uptake,demand and internal cycling of nitrogen in saplings of Mediterranean Quercus species[J]. Oecologia,2003,136(1):28-36.
[51]del Arco J M,Escudero A,Garrido M V. Effects of site characteristics on nitrogen retranslocation from senescing leaves[J]. Ecology,1991,72(2):701-708.
[52]Chapin F S Ⅲ,Moilanen L. Nutritional controls over nitrogen and phosphorus resorption from Alaskan birch leaves[J]. Ecology,1991,72(2):709-715.
[53]王晶苑,王紹強(qiáng),李紉蘭,等. 中國(guó)四種森林類型主要優(yōu)勢(shì)植物的C ∶N ∶P化學(xué)計(jì)量學(xué)特征[J]. 植物生態(tài)學(xué)報(bào),2011,35(6):587-595.
[54]Luyssaert S,Staelens J,de Schrijver A.Does the commonly used estimator of nutrient resorption in tree foliage actually measure what it claims to?[J]. Oecologia,2005,144(2):177-186.
[55]孫書存,陳靈芝. 東靈山地區(qū)遼東櫟葉養(yǎng)分的季節(jié)動(dòng)態(tài)與回收效率[J]. 植物生態(tài)學(xué)報(bào),2001,25(1):76-82.
[56]Reich P B,Walters M B,Ellsworth D S. Leaf life-span in relation to leaf,plant,and stand characteristics among diverse ecosystems[J]. Ecological Monographs,1992,62(3):365-392.
[57]邱嶺軍,胡歡甜,林寶平,等. 不同林齡杉木養(yǎng)分重吸收率及其C ∶N ∶P化學(xué)計(jì)量特征[J]. 西北林學(xué)院學(xué)報(bào),2017,32(4):22-27.
[58]蘆奕曉,楊惠敏. 隴東黃土高原混播草地牧草葉片N、P、K重吸收及生態(tài)化學(xué)計(jì)量特征[J]. 蘭州大學(xué)學(xué)報(bào)(自然科學(xué)版),2020,56(3):285-293.
[59]Kobe R K,Lepczyk C A,Iyer M. Resorption efficiency decreases with increasing green leaf nutrients in a global data set[J]. Ecology,2005,86(10):2780-2792.
[60]Tang L Y,Han W X,Chen Y H,et al. Resorption proficiency and efficiency of leaf nutrients in woody plants in Eastern China[J]. Journal of Plant Ecology,2013,6(5):408-417.
[61]Achat D L,Pousse N,Nicolas M,et al. Nutrient remobilization in tree foliage as affected by soil nutrients and leaf life span[J]. Ecological Monographs,2018,88(3):408-428.
[62]龍 靖,黃 耀,劉占鋒,等. 西沙熱帶珊瑚島典型喬木葉片性狀和養(yǎng)分再吸收特征[J]. 生態(tài)環(huán)境學(xué)報(bào),2022,31(2):248-256.
收稿日期:2023-02-01
基金項(xiàng)目:貴州省科技計(jì)劃(編號(hào):黔科合基礎(chǔ)-ZK[2021]一般231);貴州省教育廳青年科技人才成長(zhǎng)項(xiàng)目(編號(hào):黔教合KY字[2022]120、黔教合KY字[2022]123);貴州省畢節(jié)市科學(xué)技術(shù)項(xiàng)目(編號(hào):畢科合重大專項(xiàng)字[2021]1號(hào));貴州省典型高原濕地生態(tài)保護(hù)與修復(fù)重點(diǎn)實(shí)驗(yàn)室開放基金項(xiàng)目(編號(hào):畢科聯(lián)合字貴工程[2021]8號(hào))。
作者簡(jiǎn)介:何 斌(1980—),男,山西朔州人,博士,教授,從事森林生態(tài)學(xué)和植物生理生態(tài)學(xué)研究。E-mail:hebin23kewen@163.com。
