輸卵管特異性表達艾塞那肽的轉基因雞制備
2023-11-27 06:51:46張自富秦清明麻冰潔陳思睿趙云煥
中國獸醫學報 2023年9期
關鍵詞:檢測
張自富,胡 靜,趙 瑜,秦清明,麻冰潔,趙 聘,陳思睿,趙云煥
(1.信陽農林學院 動物科技學院,河南 信陽 464000; 2.中國農業大學 動物科技學院 畜禽育種國家工程實驗室,北京 100193)
近年來制備轉基因動物作為生物反應器生產藥物蛋白已經取得了許多突破性進展,開創出了誘人、廣闊的應用前景[1-2]。從發展趨勢來看,在不久的將來必定會成為生物工程技術研究領域最活躍、最具有實踐應用價值的技術。最初,哺乳動物的乳腺反應器被認為是一個最有希望和前途的生物反應器之一,可是家畜世代間隔長、成本高,特別是乳汁中乳蛋白和乳脂肪的生化復雜性給重組蛋白的純化帶來許多意想不到的困難[3-4]。與哺乳動物相比,禽類具有體格小、成本低、世代周期短、繁殖力高、卵中天然存在蛋白酶抑制劑、良好的無菌環境、好儲存、易提純等優點,特別是卵中表達的重組蛋白形成的三維結構糖基化更接近于人類蛋白質[5]。所以,禽類輸卵管生物反應器將逐步成為轉基因動物和生物制藥研究領域中最具活力的熱點,并很可能成為未來最具有競爭力的新興高技術產業之一。
但是,由于禽類獨特的生殖、生理特點和胚胎發育的復雜性,致使在哺乳動物中已經廣泛應用的轉基因技術并不適用于禽類[6]。到目前為止,研究報道較多的是利用逆轉錄病毒載體法制備出了轉基因雞,尤其是屬于逆轉錄病毒科中的慢病毒,具有容納外源目的基因片段大、可感染非分裂期細胞、整合到細胞染色體上并能長期穩定表達、不易發生基因沉默、免疫反應弱、安全性高等優點,已逐漸成為轉基因研究中應用廣泛的載體工具之一,也被公認為制備轉基因禽類最有效和最成功的方法之一[7-9]。近年來,已有利用轉基因雞輸卵管生物反應器生產重組蛋白的研究報道,包括β-內酰胺酶[10]、人干擾素α-2b[11]、人單克隆抗體[12]、單鏈Fv-Fc融合蛋白[13]、治療性蛋白[14]、人溶菌酶[15]、人防御素4[16]等。但是,以上報道均是將慢病毒注射到發育第Ⅹ期的胚盤下腔,先生產出性系嵌合體,再生產出轉基因個體。制備轉基因家禽的整個技術流程繁雜,要求條件很高[17]。另外,也有報道通過分離早期雞胚血液或性腺中的原始生殖細胞(primordial germ cells,PGCs)生產轉基因雞,但是PGCs的分離、培養、傳代等關鍵技術還一直未能獲得完全突破[18-20]。
艾塞那肽(exenatide)是從墨西哥巨蜥蜴毒液中分離出來的一種含有39個氨基酸的多肽序列,相對分子質量4.186 kDa,分子式:C184H282N50O60S,等電點pH4.86。與胰高血糖素樣肽(glucagon-like peptide1,GLP-1)具有53%的同源性,與GLP-1同作用于G-蛋白偶聯受體,并具有更高的親和力[21]。艾塞那肽是首個獲準上市的腸促胰島素類似物抗糖尿病藥物,也是目前為止公認最好的治療型糖尿病的首選藥物。在治療Ⅱ型糖尿病方面,具有可促進胰島素分泌,保護β細胞功能,改善外周胰島素敏感性,減輕體重,保護肝臟功能,半衰期長的優點[22]。2012年美國食品藥品監督管理局(FDA)批準上市,商品名為百泌達(Byetta)。
本試驗構建了一個雞卵清蛋白基因特異性啟動子調控表達艾塞那肽的慢病毒表達載體,包裝出高滴度的慢病毒,利用實驗室早期建立的生產轉基因雞新技術平臺——血管顯微注射法[23],簡便、高效的生產出輸卵管特異性表達艾塞那肽的轉基因雞。該新型技術平臺的成功建立,為利用雞輸卵管生物反應器天然、高效合成并分泌蛋白的能力,在雞蛋中生產一些具有重要價值的藥用蛋白,具有廣闊的應用前景和經濟發展潛力。
1 材料與方法
1.1 材料三質粒慢病毒表達載體系統pCSⅡ-EF-MCS-IRES2-Venus、pCAG-HIVgp和pVSV-G-Rsv-Rev為本實驗室所有;人胚腎293T細胞,快速型人胚腎293T細胞,Ultra Rapid Lentiviral Titer Kit 購自Invitrogen公司;血液及精液基因組提取試劑盒、RNase A酶購自天根生物科技有限公司,DIG High Prime DNA Labeling and Detection Starter KitⅡ購自羅氏公司;PCR引物合成上海英駿生物技術有限公司,限制性內切酶EcoRⅠ、PstⅠ和HindⅢ購自NEB公司。實驗儀器設備均為信陽農林學院中心實驗室所有。
1.2 載體構建參照GenBank中墨西哥巨蜥蜴艾塞那肽基因的cDNA序列,進行密碼子優化后化學合成cDNA(117 bp)序列,連接入T載體;在白來航雞的基因組中,分別擴增雌激素反應元件(ERE,675 bp)和卵清蛋白基因(ovalbumin gene,OVA,ID:396058)啟動子序列(包括啟動子序列、第一外顯子序列、第一內含子序列和第二外顯子一部分序列)(2 785 bp)兩種片段[14],連接入T載體中;依次酶切加入各種元件,構建于表達載體pCSⅡ-EF-MCS中,將構建好的表達質粒載體pCSⅡ-ERE-OV-EXE進行酶切測序鑒定;脂質體轉染293T細胞和雞胚成纖維細胞,初步驗證艾塞那肽基因轉錄激活功能。
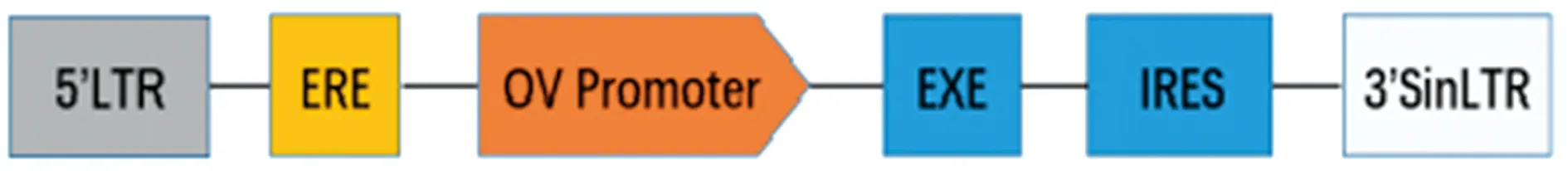
圖1 慢病毒載體pCSⅡ-ERE-OV-EXE結構示意圖
1.3 慢病毒包裝、滴度測定將包裝質粒pCAG-HIVgp、包膜蛋白質粒pCMV-VSV-G-RSV-Rev和表達載體質粒pCSⅡ-ERE-OV-EXE按1∶1∶2的比例應用磷酸鈣法轉染匯合率約為70%的293T細胞。在溫度37℃、3%的CO2培養箱中培養16 h,吸除細胞上清液,加入7.5 mL含有10 μmol/L Forskolin的完全培養基,在37℃、10%的CO2培養箱中孵育48 h,收集上清液4℃保存。收集的上清液用0.45 μm濾膜過濾,濾液50 000 r/min,溫度20℃離心2 h,棄上清,沉淀中加入500 μL HBSS,充分溶解后轉移至0.5 mL的超濾管中,14 000 r/min離心30 min,用HBSS清洗2次后,分裝-70℃保存備用。參照Ultra Rapid Lentiviral Titer Kit 試劑盒說明測定病毒滴度。
1.4 血管顯微注射慢病毒將BJ-40細玻管,規格為外徑100 μm,內徑80 μm,長度10 cm,在拉針儀拉針后用斷針儀熔斷,外徑為20~30 μm,然后在磨針儀上30度角斜面,快速磨針;在顯微鏡400倍下觀察針尖的完整性;將合格的玻璃針浸泡在75%乙醇中,用抽吸裝置清洗內壁5次,在無水乙醇中清洗5次,置于超凈臺中晾干紫外照射過夜待用。
新鮮白來航雞種蛋取回后,先用0.1%的新潔爾滅溶液清洗,除去表面污跡,然后用70%酒精噴灑消毒,待蛋殼表面晾干后,放進孵化器里進行孵化,孵化溫度為37.5℃,相對濕度為55%~65%,90°角,2 h間隔自動翻蛋,大約孵化時間52~55 h,將發育至14~15期的雞胚取出,用酒精棉擦拭表面消毒,上下輕輕晃動使胚胎脫離內層殼膜;用牙科鉆在赤道附近打出個直徑4 mm左右的小孔,仔細剔除蛋殼膜,在體式顯微鏡30倍下用顯微注射儀將1 μL 1×109TU/mL慢病毒溶液緩慢注射到雞胚卵黃外周血管中,液體石蠟封堵注射口,用parafilm封閉蛋殼開口,標記后繼續孵化至出雛。
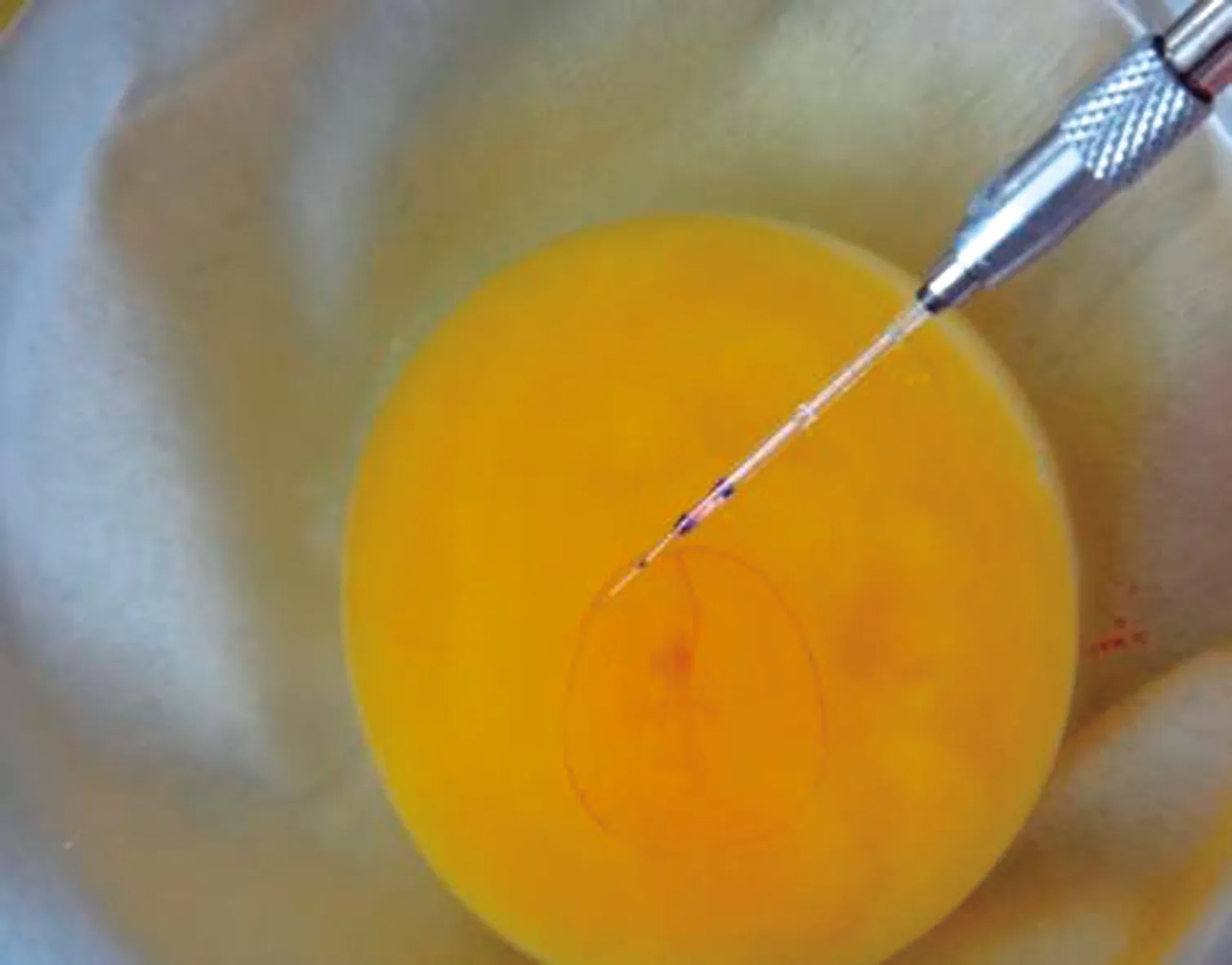
圖2 雞胚卵黃外周靜脈血管顯微注射慢病毒試驗操作演示圖
1.5 PCR檢測對發育至性成熟期G0代公雞精液基因組及G1后代血液基因組提取后PCR檢測。參照慢病毒載體pCSⅡ-ERE-OV-EXE中EXE序列信息,利用Primer 5.0軟件設計引物,進行PCR擴增。引物上游:5′-CATGCACATGAGAGGTGGATATAG-3′;引物下游:5′-GGTGTACATGATCAGCATGCTCTA-3′;擴增長度為465 bp的包含EXE基因片段。PCR擴增體系25 μL,基因組DNA 2 μL,上游引物(10 μmol/L)1.5 μL,下游引物(10 μmol/L)1.5 μL,PCR預混液12.5 μL,ddH2O 7.5 μL。PCR 反應程序:94℃預變性5 min;94℃ 30 s變性,62℃ 45 s 退火,72℃ 45 s延伸,共25個循環;72℃延伸5 min。取PCR產物5 μL,1%瓊脂糖凝膠電泳檢測。根據三磷酸甘油醛脫氫酶(GAPDH)基因序列設計特異性引物進行PCR擴增,擴增片段長度為248 bp作為參照。
1.6 Southern blot分析對PCR檢測陽性G1代個體及野生型個體,提取份血液DNA 10~20 μg,37℃水浴條件下,一份DNA用EcoRⅠ和PstⅠ雙酶切消化過夜,另一份HindⅢ酶單切消化過夜,分別用2%瓊脂糖凝膠電泳分離后轉移到尼龍膜上。根據質粒載體pCSⅡ-ERE-OV-EXE序列,隨機引物法標記探針,PCR擴增出641 bp DNA 片段,作為Southern blot的探針。地高鋅核酸探針標記Southern blot按羅氏公司DIG High Prime DNA Labeling and Detection Starter KitⅡ說明進行操作。
1.8 組織免疫化學檢測取G2代母雞輸卵管組織在PBS配制的4%多聚甲醛中固定2 h,然后用PBS洗滌20 min,4℃下30%蔗糖溶液中培養過夜。組織包埋于石蠟中,10 m厚度切片。將切片固定后封閉液中封閉2 h。小鼠抗體1∶100的比例稀釋,并在4℃下孵育過夜。免疫熒光二抗以1∶400稀釋并在室溫下孵育2 h。使用激光共聚焦顯微鏡拍照分析。野生型雞輸卵管組織切片作為陰性對照。
1.9 艾塞那肽蛋白提取及定量將來自G2代轉基因母雞和野生型各30枚蛋收集蛋清,與6倍體積冰凍的濃度為50 mmol/L醋酸鈉緩沖液(pH5.0)在4℃下混合2 h,去除大部分的卵黏蛋白。將所得混合物分裝后儲存在-20℃供進一步分析。使用ELISA試劑盒測定蛋清中人艾塞那肽濃度,每個樣品做2個重復,取2個重復均值作為1次測量值,每個樣品至少測定3次,然后根據吸光值、稀倍數和標準曲線計算出每個樣品的艾塞那肽濃度,再計算濃度均值和標準差。
2 結果
2.1 G0代公雞精液、G1代血液DNA的PCR及Southern blot檢測驗操作72枚白來航雞種蛋,雞胚發育至第14~15期(孵化52~55 h),于卵黃外周靜脈血管顯微注射1 μL病毒滴度為1×109TU/mL慢病毒,孵化出雛56只,孵化率77.8%(56/72)。至性成熟階段,采集到16只公雞精液提取DNA后進行PCR檢測,其中11只公雞陽性,陽性率68.7%(11/16)(圖3)。挑選5只陽性信號強的公雞單籠飼養,與3~5只野生型白來航母雞配種,收集種蛋,標記后進行孵化。對G1代進行血液DNA檢測,在60只G1代中檢測出5只陽性轉基因雞(編號:66,78,209,288,289)(圖4A),轉基因效率為8.3%(5/60)。為了進一步驗證,對G1代5只陽性個體進行了Southern blot 檢測,經PstⅠ和EcoRⅠ雙酶切雜交試驗結果顯示均為轉基因雞后代(圖4 C),經HindⅢ單酶切雜交實驗結果顯示外源基因單拷貝插入,且有不同的插入位點,符合孟德爾遺傳(圖4 B)。對G1代轉基因雞擴繁生產G2代。
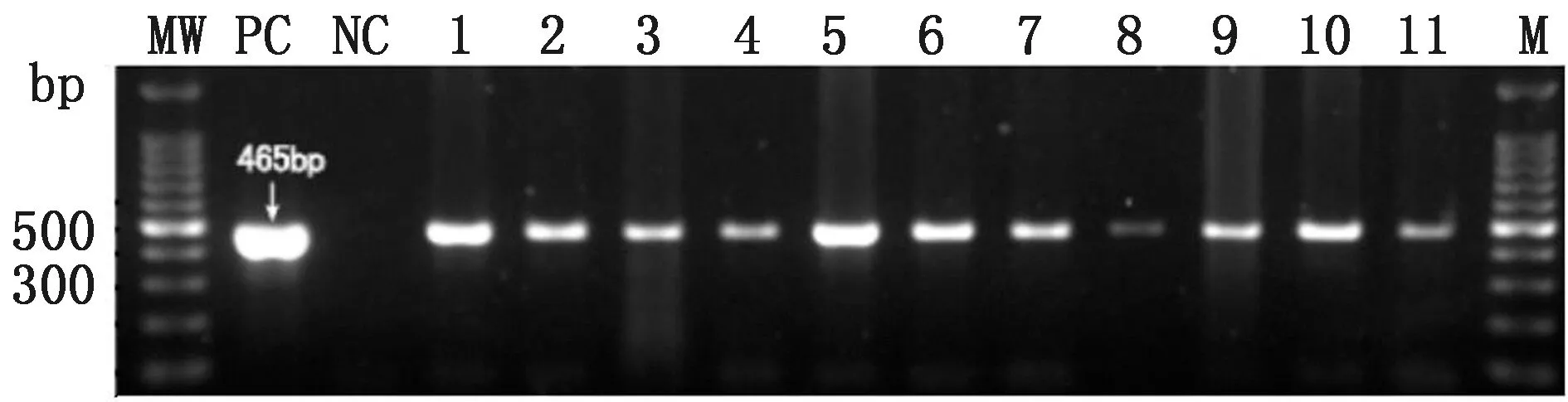
1~11.陽性個體;M.DL1000 DNA Marker;PC.陽性對照;NC.陰性對照
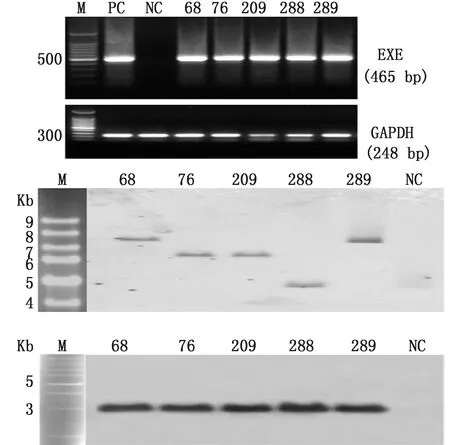
A.PCR檢測(68,76,209,288,289.為G1代中的5只陽性個體;GAPDH基因擴增作為內參;M.DL10 kb DNA Marker;PC.陽性對照;NC.陰性對照);B.血液基因組DNA的Southern blot檢測,血液基因組DNA(10~20 μg)經HindⅢ單酶切后用引物做探針雜交;C.血液基因組DNA經PstⅠ和EcoRⅠ雙酶切后雜交結果(PC,80 pg的pCSⅡ-ERE-OV-EXE質粒;NC,未注射慢病毒雞血液基因組DNA)
2.2 G2代轉基因雞卵清蛋白中艾塞那肽的檢測為了進一步檢測轉入外源基因是否出現基因沉默現象,本試驗對轉基因雞后代蛋清中的艾塞那肽的含量進行測定。本試驗對G2代陽性母雞(編號:209,288,289),待發育至性成熟期產蛋后,連續收集30個雞蛋,根據ELISA試劑盒說明書,對蛋清中的艾塞那肽進行測定,野生型白來航作對照。結果:只G2代轉基因母雞(編號:209,288,289)蛋清中艾塞那肽含量為分別為:(39.19±1.32),(42.78±2.10),(53.61±2.54) mg/L,平均為45.19 mg/L,野生型白來航雞未檢測出(圖5)。

圖5 G2代轉基因雞蛋清中艾塞那肽質量濃度測定
通過ELISA方法來測定3只G2代轉基因母雞和野生型母雞蛋清中艾塞那肽的表達量。試劑盒檢測范圍是0~1 000 ng/L。G2代轉基因母雞:G2-209,G2-288,G2-289;野生型白來航母雞未檢出。
2.3 G2代母雞輸卵管上皮免疫組織化學檢測為了進一步驗證外源基因(艾塞那肽)在卵清蛋白基因特異性啟動子調控下僅在輸卵管上皮特異性表達,對G2代轉基因母雞輸卵管組織進行了免疫組織化學檢測,野生型白來航雞作為對照。結果顯示,在G2代轉基因母雞輸卵管上皮組織中有較強的紅色抗體信號,對照組無陽性信號,進一步證實我們不僅成功制備了表達艾塞那肽的轉基因雞,且外源基因(艾塞那肽)僅在輸卵管上皮組織表達,具有組織表達特異性(圖6)。
3 討論
3.1 雞卵清蛋白特異性啟動子的表達機制本試驗構建了一個基于復制缺陷型慢病毒載體,該載體由雞卵清蛋白基因特異性啟動子調控表達外源基因(艾塞那肽),并在啟動子上游添加元件來增強表達。通過實驗室早期建立的雞胚血管顯微注射技術平臺,擬期望簡便、高效、穩定獲得轉基因雞后代,并證明外源基因(艾塞那肽)僅在輸卵管上皮組織特異地表達,既盡量減少對宿主的毒性作用,又能便于收集、分離和純化,還能保持外源蛋白的生物活性。
雞蛋蛋清是由輸卵管上皮細胞中的腺細胞分泌的,卵清蛋白基因編碼超過1/2以上的蛋清蛋白,其中卵清蛋白占54%,約為2.2 g,輸卵管上皮單個腺細胞中卵清蛋白mRNA拷貝數多達105個[24-25]。在試驗中,在白來航雞基因組中擴增卵清蛋白5′端啟動子調控序列,主要包括類固醇激素依賴調控元件(steroid dependent regulatury element,SDRE)和NRE(negative regulatury element)兩個調控元件,SDRE位于基因上游-892~-780區,可與雌激素和糖皮質激素結合,產生相應的應答反應,促進卵清蛋白基因的轉錄;NRE位于基因上游-308~-88,該元件具有雙重作用,在雌激素作用下會增強卵清蛋白基因的表達,反之抑制卵清蛋白基因的表達[26];另外擴增出包括OVA第1外顯子、第1內含子和第2外顯子開始部分,長度為2 785 bp雞卵清蛋白啟動子調控序列,既能保證外源基因的表達,又能保證表達的組織特異性[12,14]。為了進一步增加外源基因的表達,在OVA 啟動子上游插入了一個雌激素應答原件(ERE,675 bp)來進一步促進目的基因的轉錄[14-16]。艾塞那肽是一種含有39個氨基酸的多肽序列, cDNA序列(117 bp),連接在OVA啟動子上游5′端,總共長度約3.6 kb,符合選擇的復制缺陷型慢病毒系統要求,未超出其有效載荷,成功包裝出高滴度的慢病毒。對獲得的轉基因雞后代進行免疫組織化學檢測,結果證明了艾塞那肽蛋白僅特異地在輸卵管上皮組織管狀腺細胞內表達,即卵清蛋白合成區域,其他上皮組織細胞未見,具有表達特異性,符合試驗設計預期,也與其他研究者報道一致[14-16]。雖然本試驗成功獲得表達外源目的基因的轉基因雞,但是從卵清蛋白中檢測的目的蛋白含量與其他研究報道相比,具有一定的差異性。在試驗中測得G2代轉基因母雞蛋清中艾塞那肽含量平均為45.19 mg/L,與LILLICO等[14]和CAO等[15]報道結果接近,比LIU等[16]報道的高出多倍。構建同樣的表達載體及元件,在轉基因雞后代中目的蛋白的表達卻出現較大差異,推測可能是慢病毒載體的隨機插入造成目的蛋白的表達差異[27],也可能是對轉基因后代中檢測數據方法不一致造成的,如有的檢測G1代,有的檢測G2代,對轉基因雞后代檢測統計數目偏小也可能造成人為誤差;另外也有可能是表達載體不同引起的,如EIAV相對于HIV,同屬于慢病毒,對宿主造成的毒性較小,且攜帶的外源基因序列更大[14]。
3.2 轉基因雞制備效率近年來,利用轉基因雞輸卵管生物反應器生產藥用蛋白已取得了很多可喜的成果,其中胚盤下腔顯微注射慢病毒法和PGCs體外轉染法是公認比較成功的方法。ZHU等[12]利用構建的卵清蛋白特異性啟動子的慢病毒載體,通過胚盤下腔顯微注射慢病毒法首次制備出表達人源化融合性單克隆抗體的嵌合體雞,并且在蛋清中表達的單克隆抗體產量及生物活性都令人感到振奮,但未能獲得性系傳遞的轉基因后代。LILLICO等[14]將攜帶有卵清蛋白特異性啟動子調控序列分別表達miR24和hINFβ1a兩種慢病毒載體(EIAV),通過同樣的方法成功制備出轉基因雞后代,而且兩者都能檢測到輸卵管組織特異性表達具有完全生物活性的藥用蛋白及人源化抗體,且能穩定遺傳給后代。KWONT等[28]通過同樣的方法成功制備在卵清蛋白中特異性表達人干擾素的轉基因鵪鶉。BON KOO等[29]利用逆轉錄病毒載體胚盤下腔顯微注射成功制備了特異性表達人促紅細胞生成素的轉基因雞。在國內,有研究者通過同樣的方法成功制備在卵清蛋白中特異性表達重組人防御素4和人溶菌酶兩種藥用蛋白的轉基因雞[15-16]。以上報道是將病毒載體注射到發育第Ⅹ期的胚盤下腔,通過感染囊胚期40 000~60 000個細胞中的幾個PGCs,先生產出性系嵌合體。首先需要在雞胚孵化過程中進行繁雜的倒三期培養體系[17];其次,必須包裝出高滴度的慢病毒;第三,要求特定的試驗設備,同時要求顯微注射操作者必須精準、熟練,且經過嚴格訓練。另外,利用PGCs體外轉染法制備轉基因雞被認為是一種有效的方法[18-20],但是,由于家禽PGCs培養的特殊性(需要滋養層細胞,培養血清中需要添加多種特殊的抑制細胞分化因子),加上PGCs的分離、純化、培養、傳代、體外轉染、篩選等一系列技術環節周期長,繁瑣且費時費力,而且還不能確保轉染后的PGCs注射到同期發育胚胎后,能否遷移至受體雞胚的生殖原基進一步分化形成配子(精子或卵子),所以還一直未能做到真正的推廣應用。
本試驗構建了一個雞卵清蛋白基因特異性啟動子調控表達艾塞那肽的慢病毒載體,嘗試把顯微注射法、慢病毒載體法兩種轉基因技術與早期雞胚PGCs遷移規律的生殖生理特點結合起來,選擇雞胚發育至14~15期,血液中PGCs向生殖原基遷移高峰期,將高滴度的慢病毒載體注入血管中,隨血液循環而感染其中的PGCs,最終獲得了可表達外源基因(艾塞那肽)的轉基因雞。與經典的生產轉基因家禽方法相比,不僅省去復雜的倒三期培養體系及PGCs的分離、純化、培養、建系、轉染、篩選后再注回同期受體胚胎等一系列繁雜程序操作,而且節約了大量的人力、物力、財力,轉基因雞的制備綜合效率(G0代孵化率,G1代陽性率等)整整提高了上百倍。本研究組建立的簡便、高效、穩定生產轉基因雞的創新技術體系,在卵清蛋白特異性啟動子調控作用下,利用雞輸卵管天然、高效合成并分泌蛋白的能力,在雞蛋中生產一些具有重要價值的藥用蛋白,具有廣闊的應用前景和經濟發展潛力。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
