山西省2018-2020年豬圓環(huán)病毒3型的檢測及Cap基因遺傳演化分析
2023-11-27 06:51:36薛翼鵬米瑞娟
中國獸醫(yī)學報 2023年9期
孟 帆,薛翼鵬,趙 岳,米瑞娟,吳 忻
(山西農(nóng)業(yè)大學 動物醫(yī)學學院,山西 太谷 0030801)
豬圓環(huán)病毒 3 型(porcine circovirus type 3,PCV3)是由美國的兩位科學家于2015年首次分離到的[1-2],屬于圓環(huán)病毒科、圓環(huán)病毒屬的新成員,現(xiàn)已在全球養(yǎng)豬國家逐漸蔓延[3-12],給世界養(yǎng)豬業(yè)造成了不可估量的損失。作為世界上豬肉消費量最高的國家,中國在全球生豬生產(chǎn)中也發(fā)揮著不可低估的作用,自從PCV3病例在我國豬場的存在被學者初次報道以來[12],陸續(xù)有學者報道了PCV3在我國各個不同省市的感染[13-26],甚至有學者通過回溯性研究發(fā)現(xiàn)近半個世紀以來PCV3已存在于豬群中[14]。豬作為PCV3唯一的宿主,所有的豬均可感染,特別是妊娠母豬和仔豬更易感染[2,7],造成妊娠母豬繁殖障礙,產(chǎn)木乃伊胎、死胎等[1],仔豬出現(xiàn)神經(jīng)、呼吸、心血管、腸道、泌尿、淋巴、繁殖系統(tǒng)、皮膚等多器官系統(tǒng)的功能紊亂和先天性震顫等[13]。還有研究認為PCV3侵入豬的免疫系統(tǒng)后可造成豬的免疫力降低[24],更容易感染豬偽狂犬病病毒、豬繁殖與呼吸障礙綜合征病毒、豬圓環(huán)病毒2型、豬流行性腹瀉病毒等病原,引起混合感染,對養(yǎng)豬業(yè)造成重大威脅[25]。因此對PCV3的流行及遺傳變異情況進行綜合分析很重要,及時掌握PCV3的流行動態(tài)及遺傳變異情況,以便提前進行預警,及時進行防控。本研究運用 PCR 技術(shù)對2018-2020年在山西省豬場采集的1 125份血清和病、死豬組織樣品進行PCV3檢測,然后對獲得的5份PCV3 陽性樣品ORF2序列進行測序,并對ORF2序列和推導出的Cap蛋白氨基酸序列進行遺傳變異分析,旨在了解近年來山西省PCV3的流行情況和遺傳進化特點,希望為山西省PCV3的科學防控提供依據(jù)。
1 材料與方法
1.1 樣品來源2018-2020年在山西省忻州、呂梁等11地市豬場,隨機從存欄豬的耳靜脈或前腔靜脈抽取血液,分離血清,并無菌采集病、死豬的淋巴結(jié)、扁桃體等組織放入DNA 組織樣品保存液備用。
1.2 主要試劑Virus DNA/RNA Extraction Kit 2.0(南京諾唯贊生物科技有限公司提供);premix TaqTM(Ex TaqTMVersion 2.0)(大連寶生物工程有限公司提供);PCV3型PCR檢測試劑盒(哈爾濱元亨生物藥業(yè)有限公司提供);DNA膠回收試劑盒和普通質(zhì)粒小提取試劑盒(上海生工生物工程有限公司提供)。
1.3 引物設(shè)計及合成按照文獻[3]的方法于PCV3(GenBank中查閱)保守序列設(shè)計出1對特異性引物。上游引物PF2:5- AATAAACTATTGATTTTATTTGCACTTGTGT -3′;下游引物PR2:5′- GCCCGGCACCAAAATGA -3′。引物(上海生工生物工程有限公司合成)使用前用DEPC水溶解,濃度為10 μmol/L,-80℃保存?zhèn)溆谩?/p>
1.4 血清及病料樣品DNA的提取把采集到的血清及病料按照Virus DNA/RNA Extraction Kit 2.0操作說明書提取病毒基因組DNA,-20℃保存?zhèn)溆谩?/p>
1.5 PCR擴增反應按照TaKaRa LA PCRTMKit Ver.2.1試劑盒說明書,以提取的基因組DNA為模板進行擴增。經(jīng)過優(yōu)化,反應體系為25 μL:dNTP Mixture(2.5 mmol/L each)4 μL,10×LA Taq Buffer Ⅱ(Mg2+Plus)2.5 μL,上下游引物(10 μmol/L)各1 μL,TaKaRa LA Taq(5 U/μL) 0.25 μL,模板2 μL,DEPC水補充到25 μL。反應條件:94℃ 5 min;進入PCR循環(huán)(35)94℃ 30 s,56℃ 30 s,72℃ 45 s;72℃ 10 min。然后取 PCR產(chǎn)物5 μL進行凝膠電泳試驗,而后對擴增結(jié)果進行檢測。
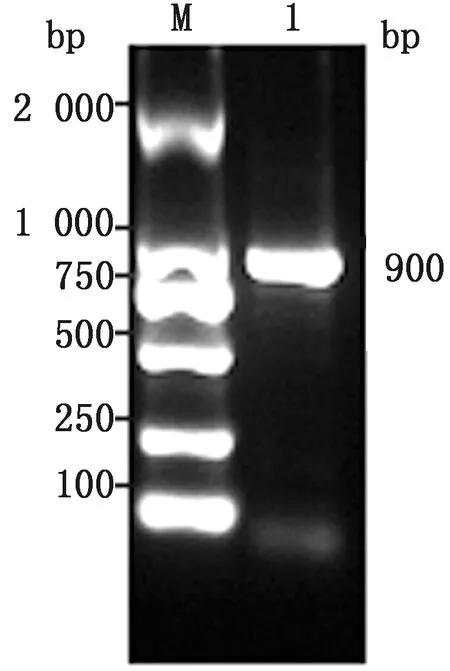
M.DL2000 DNA Marker;1.PCV PCR產(chǎn)物
1.6 PCV3擴增產(chǎn)物的克隆及序列測定參照文獻[25]的方法對PCV3擴增產(chǎn)物進行克隆及序列測定。
1.7 PCV3 ORF2 核苷酸序列分析參照文獻[25]的方法將PCV3 ORF2核苷酸序列及推導出的Cap蛋白氨基酸序列與GenBank中己發(fā)表的36株P(guān)CV3參考株進行同源性及遺傳進化分析。
1.8 臨床樣品的檢測用PCR 方法對 2018-2020年采集于山西省豬場的1 125份血清和病、死豬組織樣品進行 PCV3 檢測和分析。
2 結(jié)果
2.1 2018-2020年山西省PCV3型檢測結(jié)果對山西省 2018-2020年P(guān)CV3檢測結(jié)果顯示(表1),2018-2020年山西省PCV3平均陽性率為21.42%,豬場平均陽性率為44.17%。2018-2020年P(guān)CV3陽性率呈逐年上升趨勢,豬場陽性率整體也呈上升趨勢。
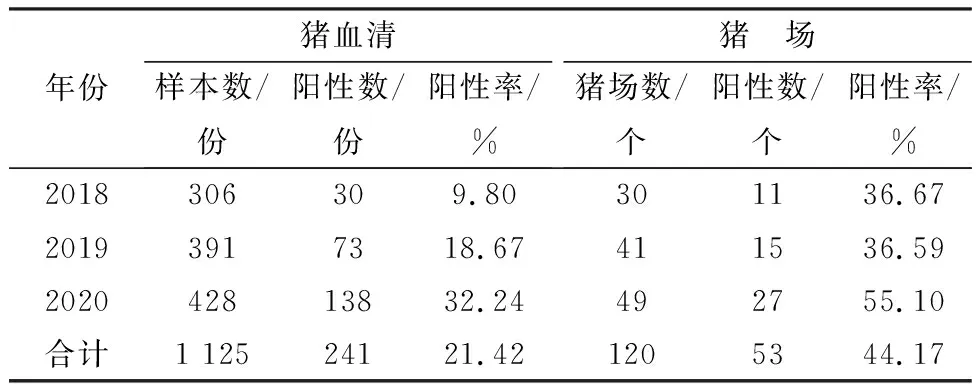
表1 2018-2020年山西省部分豬場PCV3檢測結(jié)果
2.2 PCV3 ORF2核苷酸的擴增結(jié)果用引物對PCV3陽性樣品進行PCR擴增,凝膠電泳結(jié)果顯示擴增出與預期片段大小相同的900 bp擴增條帶(圖1)。
2.3 PCV3 ORF2核苷酸序列分析山西省5株P(guān)CV3 ORF2核苷酸序列大小均為645 bp,編碼214 aa。將山西省5株P(guān)CV3 ORF2核苷酸序列與36株國內(nèi)外不同地區(qū)的PCV3 ORF2 核苷酸序列進行同源性比較,結(jié)果表明(圖2),41株P(guān)CV3 ORF2 核苷酸序列同源性為97.5%~100.0%;山西省5株P(guān)CV3 ORF2 核苷酸序列同源性為99.2%~100.0%,與36株P(guān)CV3參考株ORF2 核苷酸序列同源性為98.1%~99.8%。其中,山西省5株P(guān)CV3 ORF2 序列中PCV3-SXCZ株與 PCV3-SXYC株之間核苷酸同源性最高,為100.0%,PCV3-SXJC株與PCV3-SXXZ株之間核苷酸同源性最低,為99.2%。山西省5株P(guān)CV3 ORF2序列與國外13株P(guān)CV3參考株ORF2 序列之間,PCV3-SXJZ株與4332-7_Denmark_2017株(丹麥)ORF2 序列核苷酸同源性最高,為99.8%,PCV3-SXXZ株與PCV3-RU/SM17株(俄羅斯)ORF2 序列核苷酸同源性最低,為98.3%;與國內(nèi)23株P(guān)CV3參考株之間,PCV3-SXXZ株與PCV3/CN/Jiangxi-3/2016株(江西)及PCV3/Pig/CN/ShanXi170709株(陜西)ORF2 序列核苷酸同源性最高,為99.8%,PCV3-SXJC株與PCV3-CN/FuJian-318-2017株(福建)及PCV3-CN/FuJian-820-2016株(福建)ORF2 序列核苷酸同源性最低,為98.3%。進一步分析結(jié)果表明(圖3),山西省5株P(guān)CV3 ORF2 核苷酸序列與36株國內(nèi)外不同地區(qū)的PCV3參考株相比,僅存在幾處堿基的突變,PCV3-SXJC株有1個核苷酸的差異,即13位C→T;PCV3-SXCZ株有1個核苷酸的差異,即70位C→T,同737-8_Spain_2017(西班牙)一致。
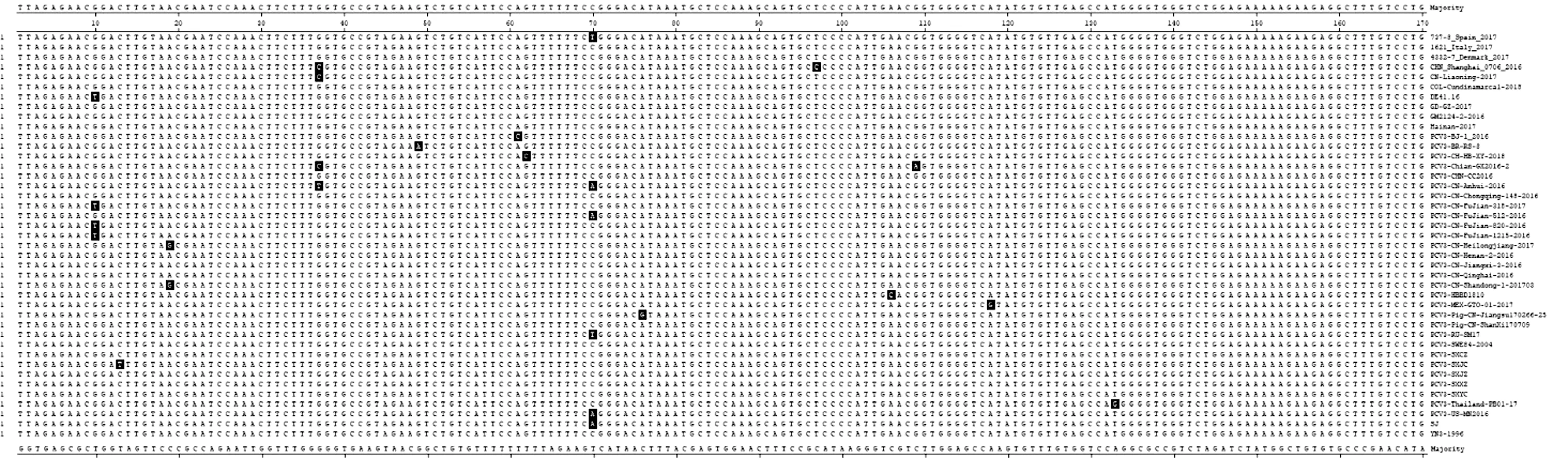
圖3 PCV3 ORF2核苷酸序列比較
2.4 PCV3 ORF2 核苷酸序列系統(tǒng)發(fā)生樹分析在分析山西省5株P(guān)CV3 ORF2序列和36株國內(nèi)外不同地區(qū)的PCV3參考株的ORF2序列核苷酸同源性結(jié)果的基礎(chǔ)上,構(gòu)建了PCV3 ORF2核苷酸序列系統(tǒng)進化樹。進化樹結(jié)果顯示(圖4),41株P(guān)CV3流行株分成2大分支,分別為3a和3b基因群,其中山西省5株P(guān)CV3流行株都屬于3b基因群。進一步分類可以看出,PCV3-SXJC株與PCV3/CN/Henan/2/2016株(河南)親緣關(guān)系較近,PCV3-SXJZ株、PCV3-SXYC株和PCV3-SXCZ株與PCV3-BJ-1_2016株(北京)、4332-7_Denmark_2017株(丹麥)和PCV3-Chian/GX2016-2株(廣西)位于同一小分支親緣關(guān)系較近,PCV3-SXXZ株與GD-GZ/2017株(廣東)、PCV3/CN/Qinghai/2016株(青海)和PCV3/CN/Jiangxi-3/2016株(江西)親緣關(guān)系較近。

圖4 PCV3 ORF2系統(tǒng)發(fā)生樹
2.5 PCV3Cap蛋白氨基酸序列分析結(jié)果顯示(圖5),41株P(guān)CV3 ORF2核苷酸序列推導的氨基酸同源性為96.7%~100.0%;山西省5株P(guān)CV3序列之間Cap蛋白氨基酸同源性為98.1%~100.0%;與36株參考株之間Cap蛋白氨基酸同源性為97.2%~100.0%。山西省5株P(guān)CV3 Cap蛋白氨基酸序列中,PCV3-SXCZ株與 PCV3-SXJZ株之間Cap蛋白氨基酸同源性最高,PCV3-SXJZ株與 PCV3-SXYC株之間Cap蛋白氨基酸同源性最高,均為100.0%,PCV3-SXXZ株與PCV3-SXJC株之間Cap蛋白氨基酸同源性最低為98.1%。山西省5株P(guān)CV3 Cap蛋白氨基酸序列與國外13株P(guān)CV3參考株之間,PCV3-SXCZ株、PCV3-SXJZ株和PCV3-SXYC株與DE41.16株(德國)、PCV3-SWE84-2004株(瑞士)Cap蛋白氨基酸同源性最高,PCV3-SXXZ株與PCV3-BR-RS-8株(巴西)、1621_Italy_2017株(意大利)、PCV3/MEX/GTO/01/2017株(墨西哥)、SJ株(韓國)、737-8_Spain_2017株(西班牙)Cap蛋白氨基酸同源性最高,PCV3-SXJC株與PCV3/Thailand/PB01/17(泰國)株Cap蛋白氨基酸同源性最高,均為100.0%,PCV3-SXXZ株與PCV3-RU/SM17株(俄羅斯)氨基酸同源性最低,PCV3-SXJC株與GM2124-2/2016株(日本)氨基酸同源性最低,均為97.7%;與國內(nèi)23株P(guān)CV3參考株之間,PCV3-SXCZ株、PCV3-SXJZ株和PCV3-SXYC株與CHN_Shanghai_0706_2016株(上海)、PCV3-CN/FuJian-512-2016株(福建)、PCV3-CN/FuJian-1215-2016株(福建)Cap蛋白氨基酸同源性最高,PCV3-SXXZ株與PCV3/CN/Jiangxi-3/2016株(江西)、PCV3/Pig/CN/Shanxi170709株(陜西)、PCV3-CHN/CC2016株(吉林長春)、PCV3/CN/Chongqing-148/2016株(重慶)Cap蛋白氨基酸同源性最高,PCV3-SXJC株與PCV3/CN/Anhui/2016株(安徽)、PCV3/CN/Henan/2/2016株(河南)、PCV3/CN/Shandong-1/201703株(山東)Cap蛋白氨基酸同源性最高為100.0%,PCV3-SXJC株與GD-GZ/2017株(廣東)Cap蛋白氨基酸同源性最低為97.2%。進一步分析顯示,山西省5株P(guān)CV3 Cap蛋白氨基酸序列較為保守,沒有出現(xiàn)不同于其他流行株的特異性突變。
3 討論
PCV3型雖然是2016年才發(fā)現(xiàn)的一個新型病毒[1-2],但實際上至少已在中國豬群中存在了近半個世紀之久[14],豬只感染后造成妊娠母豬繁殖障礙,仔豬多器官系統(tǒng)功能紊亂、先天性震顫等,病毒侵入豬的免疫系統(tǒng)后引起豬免疫力降低,極易感染其他病毒,引起混合感染,造成豬只生產(chǎn)性能降低,PSY指標下降,對中國豬業(yè)造成了嚴重的影響。近年來,隨著山西省生豬產(chǎn)業(yè)的迅猛發(fā)展,對山西地區(qū)PCV3型進行分子流行病學調(diào)研和遺傳變異分析很重要,可以及時掌握PCV3的流行動態(tài)及遺傳變異情況,基因的缺失、嵌入和重組可導致病毒毒力增強或致弱,結(jié)合臨床表現(xiàn)及早做出分析,有利于提前進行預警,對重點防控工作做出調(diào)整,及時進行應對。
2018-2020年間通過對山西省PCV3型的分子流行病學調(diào)研,發(fā)現(xiàn)PCV3在山西省各地市的豬場都有檢出,PCV3平均陽性率為21.42%,豬場平均陽性率為44.17%,說明PCV3在山西省豬場感染嚴重,已呈流行態(tài)勢,平均陽性率呈逐年上升的趨勢。郭影成等[16]報道吉林省2015-2017年P(guān)CV3型樣品陽性率和豬場陽性率都呈逐年上升的趨勢;段群棚等[17]報道2017-2019年間廣西PCV3型樣品陽性率和豬場陽性率都呈逐年上升的趨勢;李勇等[15]對華東地區(qū)豬場從2011-2017年不同季節(jié)采集的 600多份樣品進行了回溯性篩查,結(jié)果發(fā)現(xiàn)豬場一直存在PCV3型的感染,感染率呈逐年上升的趨勢,且在春冬季節(jié)相對較高,沒有呈現(xiàn)出明顯的規(guī)律性;劉燕等[19]對2017-2019年從鄭州地區(qū)豬場采集的1 175份樣品進行PCV3型檢測,結(jié)果發(fā)現(xiàn),2017-2019年鄭州地區(qū)豬場PCV3型陽性率呈現(xiàn)逐年上升的趨勢;這些報道都與本研究結(jié)果相一致。可見目前山西省PCV3型感染在豬場已呈流行態(tài)勢,形勢不容樂觀,只能憑借健全的生物安全措施加以防控,因此必須引起相關(guān)從業(yè)者的高度重視,把該病的凈化及綜合防控列入日常工作中。PCV3包含2個主要開放閱讀框(ORF)[1],其中復制酶蛋白Rep由ORF1編碼,衣殼蛋白Cap由ORF2編碼,對PCV3的ORF2序列進行遺傳變異分析意義深遠。對獲得的5份PCV3 陽性樣品的ORF2序列進行測序,并對ORF2序列和推導出的Cap蛋白氨基酸序列進行遺傳變異分析,發(fā)現(xiàn)山西省5株 PCV3的ORF2核酸序列均為645 bp,編碼214個氨基酸,5株 PCV3 ORF2之間核苷酸和Cap蛋白氨基酸同源性分別為 99.2%~100.0%和 98.1%~100.0%,與36株參考株之間核苷酸和氨基酸同源性分別為98.1%~99.8%和 97.2%~100.0%;可見PCV3之間ORF2核苷酸序列和Cap蛋白氨基酸序列同源性都很高,這與蘇戰(zhàn)強等[18]的研究結(jié)果相一致。Cap蛋白氨基酸序列沒有出現(xiàn)不同于其他毒株的特異性突變,表明PCV3在山西較為保守,尚未出現(xiàn)明顯變異。PCV3 ORF2核苷酸序列系統(tǒng)進化樹結(jié)果顯示,山西5株P(guān)CV3流行株都屬于3b基因群,表明近年來山西PCV3流行株以PCV-3b亞型為主。其中PCV3-SXJC株與PCV3/CN/Henan/2/2016株(河南)親緣關(guān)系較近,PCV3-SXJZ株、PCV3-SXYC株和PCV3-SXCZ株與PCV3-BJ-1_2016株(北京)、4332-7_Denmark_2017株(丹麥)和PCV3-Chian/GX2016-2株(廣西)位于同一小分支,親緣關(guān)系較近,PCV3-SXXZ株與GD-GZ/2017株(廣東)、PCV3/CN/Qinghai/2016株(青海)和PCV3/CN/Jiangxi-3/2016株(江西)親緣關(guān)系較近,可見山西PCV3流行株來源并不單一,因此山西各養(yǎng)豬場應該提高風險意識,在引種時加強檢疫、隔離措施等;另外應加強殺蟲滅鼠、定期監(jiān)測、自繁自養(yǎng)等生物安全措施的防控。
本研究結(jié)果為探討PCV3在山西省的流行和遺傳進化特點提供了理論參考,以期為山西省PCV3的科學防控提供依據(jù)。
