非洲豬瘟病毒p22 蛋白單克隆抗體的制備及鑒定
2023-11-23 03:26:38矯健李建達韓先杰張琳張玉玉任素芳劉飛陳智NataliiaHrabchenko張文娟于江吳家強
山東農業科學 2023年10期
矯健,李建達,韓先杰,張琳,張玉玉,任素芳,劉飛,陳智,Nataliia Hrabchenko,張文娟,于江,吳家強
(1. 青島農業大學動物醫學院,山東 青島 266109;2. 山東省農業科學院畜牧獸醫研究所/山東省畜禽疫病防治與繁育重點實驗室/農業農村部畜禽生物組學重點實驗室,山東 濟南 250100;3. 山東碧藍生物科技有限公司,山東 泰安 271400)
非洲豬瘟(African swine fever, ASF)是由非洲豬瘟病毒(African swine fever virus, ASFV)引起的一種高度傳染性、出血性疾病,臨床上以高熱、皮膚發紺、淋巴結及臟器廣泛出血為主要特征,如腎瘀斑、皮膚紅斑、肺水腫、彌散性血管內凝血等[1]。 ASFV 主要通過接觸性傳播,家豬和野豬都是其主要宿主,一旦侵入家豬,便可在豬群中快速傳播,發病率和死亡率高達100%[2-3],感染性病毒在組織中能夠存在較長時間,尤其是淋巴結和扁桃體[4]。 此外,該病毒也是唯一已知的DNA 蟲媒病毒,能夠通過軟蜱進行傳播[5]。 ASF于1921 年首次在肯尼亞報道,隨后傳入其他非洲國家[6]。 2007 年,在格魯吉亞發現了毒力更高的ASFV Ⅱ型,并迅速傳播[7]。 2018 年8 月,我國沈陽首次暴發ASF,隨后迅速蔓延到全國所有省份[8],對我國乃至全世界的養豬業造成了嚴重的經濟損失。
ASFV 是Asfarviridae 科中Asfarvirus屬的唯一成員[9],是一種有囊膜的雙鏈DNA 病毒。 ASFV粒子是具有多層結構的二十面體,直徑約為200 nm,病毒粒子結構自內向外依次為類核、核殼、內囊膜、衣殼、外囊膜[10-11]。 ASFV 基因組長度約為170~194 kb,目前已從其中鑒定出結構蛋白68種,部分蛋白的功能尚不清楚[12]。 這些結構蛋白分布在不同的區域,在病毒黏附與入侵、病毒基因組復制和轉錄、病毒包裝與出芽以及免疫逃避等病毒感染的不同方面發揮著重要的作用[5]。 由于ASFV 結構復雜繁冗,對其功能的了解也十分有限,到目前為止還沒有完全有效的ASFV 商業疫苗,因此鑒定病毒結構蛋白的免疫原性在探討病毒與宿主相互作用中至關重要[13]。
ASFV p22 蛋白由KP177R基因編碼,分子量為22 kDa。 p22 最初被描述為一種早期誘導的跨膜結構蛋白,位于病毒顆粒外部,能夠從病毒顆粒表面溶解釋放出來[14]。 然而,進一步的研究發現,p22 蛋白位于病毒顆粒的內膜和感染細胞的表面[15]。 目前,p22 蛋白在ASFV 感染致病過程中的作用仍不清楚。 本研究通過大腸桿菌原核表達系統成功表達并純化出p22 重組蛋白,通過免疫Balb/c 小鼠成功制備了其單克隆抗體(MAb),為進一步探究p22 蛋白的生物學作用奠定基礎。
1 材料與方法
1.1 細胞、菌株、載體及實驗動物
大腸桿菌Trans5α、BL21(DE3)感受態細胞購自北京全式金生物技術有限公司;骨髓瘤細胞SP2/0、pET-32a 載體均由本實驗室保存;ASFV標準陽性血清購自中國獸醫藥品監察所;SPF 級5~8 周齡Balb/c 雌性小鼠購自濟南朋悅試驗動物繁育有限公司。
1.2 主要試劑
質粒小提試劑盒、膠回收試劑盒均購自天根生化科技(北京)有限公司;限制性核酸內切酶、T4 DNA 連接酶、蛋白Marker 均購自賽默飛世爾科技公司;Ni-NTA Agarose 購自德國Qiagen 公司;單抗亞型鑒定試劑盒購自美國Southern Biotech公司;ECL 發光液、羊抗鼠HRP-IgG、羊抗豬HRPIgG 購自Beyotime 公司。
1.3 ASFV p22 蛋白的原核表達
1.3.1 重組表達質粒pET-32a-p22 的構建 根據ASFV pig/HLJ/2018 株基因序列(GenBank No.MK333180.1)的KP177R基因序列進行引物設計,正向引物(p22-F)序列:5'-CTCGAGTTATGCGTGTTTATGATTAC-3'(包含XhoⅠ酶切位點), 反向引物(p22-R) 序列: 5' - GGATCCAAAAAACAGCAGCCGCCGA-3'(包含BamHⅠ酶切位點),由生工生物工程(上海)股份有限公司合成。 以合成的密碼子優化后的p22基因序列為模板,使用引物擴增目的基因片段,PCR 反應程序為95 ℃,5 min;94 ℃,45 s,58 ℃,45 s,72℃,45 s,32 個循環;72 ℃,10 min;4 ℃結束。 將擴增產物進行1%的瓊脂糖凝膠電泳并純化回收,用T4 DNA 連接酶分別將KP177R基因與pET-32a 載體連接,將連接產物轉化到感受態細胞Trans5α 中。 挑取單克隆搖菌,菌液經PCR 鑒定為陽性后,送至生工生物工程(上海)股份有限公司測序。 測序成功后將陽性重組質粒命名為pET-32a-p22。
1.3.2 ASFV p22 重組蛋白的表達和純化 將陽性重組質粒pET-32a-p22 轉化至大腸桿菌BL21(DE3)感受態細胞,挑取單克隆至LB 培養基中37 ℃培養過夜,取1 mL 菌液接種至氨芐青霉素(ampicillin,Amp)抗性的100 mL LB 液體培養基中,37 ℃、220 r/min 搖床培養至OD600值達到0.6~0.8 時,添加終濃度為1 mmol/L 的IPTG 誘導目的蛋白表達,誘導4 h 后,收集菌體沉淀,使用裂解液( 300 mmol/L NaCl, 50 mmol/L NaH2PO4,pH=7.4)重懸菌體,進行超聲破碎,4 ℃、4 000 r/min 離心,經SDS-PAGE 電泳鑒定后使用0.45 μm 濾膜過濾,與蛋白純化填料4 ℃過夜結合。 采用親和層析的方式純化,收集洗脫產物,用于SDS-PAGE 鑒定,使用超濾管將有單一目的蛋白條帶的洗脫產物濃縮并置換到PBS中,-80 ℃保存。
1.3.3 ASFV p22 重組蛋白的Western blot 鑒定重組p22 蛋白純化后進行SDS-PAGE 電泳,隨后轉移到PVDF 膜上,經5%脫脂奶粉封閉后,以ASFV 標準陽性血清(1∶1 000)作為一抗孵育2 h。清洗后,加入HRP 標記的羊抗豬IgG 二抗(1∶8 000)孵育1 h,進行Western blot 鑒定。
1.4 ASFV p22 單克隆抗體的制備
1.4.1 Balb/c 小鼠的免疫及血清效價的測定純化后的重組p22 蛋白與等體積弗氏完全佐劑乳化后,對4 只5~8 周齡Balb/c 小鼠進行背部多點皮下注射,免疫劑量為100 μg/只,每14 d 免疫一次,共免疫3 次。 最后一次免疫7 d 后從小鼠尾靜脈取少量血,分離血清,通過間接ELISA 方法檢測血清中的抗體效價。 將純化的p22 重組蛋白以2 g/mL 的濃度包被到聚苯乙烯96 孔反應板(100 μL/孔),4 ℃放置過夜;次日用PBST 洗板3次后進行封閉(l50 μL/孔封閉液),室溫放置0.5 h;用樣品稀釋液將小鼠血清按1∶2 000,1∶4 000,1∶8 000,1∶16 000,1∶32 000,1 ∶64 000,1∶128 000……進行倍比稀釋,洗板3 次后加入稀釋好的血清并設置空白血清做陰性對照(100 μL/孔),室溫孵育1 h;洗板3 次后加入HRP 標記的鼠二抗(1∶5 000,100 μL/孔),37 ℃孵育40 min;PBST 洗板5 次并拍干,加入TMB(100 μL/孔),室溫避光顯色10 ~15 min;加入終止液(50 μL/孔),用酶標儀測定450 nm 處各孔的吸光值。
1.4.2 細胞融合及亞克隆篩選 按照標準流程取免疫后的小鼠脾細胞和骨髓瘤細胞SP2/0 融合,通過間接ELISA 方法檢測篩選陽性雜交瘤細胞,使用有限稀釋法對單克隆陽性孔進行3 次亞克隆。 篩選出穩定表達p22 的陽性孔,進行擴大培養。
1.4.3 腹水制備 將陽性雜交瘤細胞擴大培養并注射至Balb/c 小鼠的腹腔(經弗氏不完全佐劑乳化),7~10 d 后觀察小鼠腹部隆起狀況,并及時抽取腹水。 12 000 r/min 離心10 min,收集上清,-70 ℃凍存備用。
1.5 ASFV p22 單克隆抗體的鑒定
1.5.1 單克隆抗體效價鑒定 將進行不同比例稀釋后的腹水作為一抗,HRP 標記的羊抗鼠IgG作為二抗,用重組p22 蛋白建立的間接ELISA 方法進行效價的測定(方法同1.4.1)。
1.5.2 單克隆抗體亞型鑒定 用小鼠抗體亞型鑒定試劑盒對雜交瘤細胞制備的腹水抗體進行亞型測定。 準備好包被有重組p22 蛋白的板條(50 ng/孔),每個克隆收集600 μL 腹水上清,分別滴加到6 個對應的酶標孔中(100 μL/孔),37 ℃孵育1 h;PBST 洗板3 次,將稀釋好的分型二抗抗IgM、IgA、IgG1、IgG2a、IgG2b、IgG3 的抗體加入6個孔中,37 ℃孵育1 h;PBST 洗板3 次,加入TMB顯色,有信號反應的孔對應的鑒定二抗亞型即為該抗體的亞型。
1.5.3 Western blot 鑒定單克隆抗體的免疫原性純化的重組p22 蛋白經SDS-PAGE 電泳后,轉移至PVDF 膜上,使用5%脫脂奶粉封閉2 h;PBST 洗膜后,加入單克隆抗體(1∶1 000),4 ℃孵育過夜;清洗后,加入HRP 標記的羊抗鼠IgG 二抗孵育1 h,充分清洗后使用ECL 發光液進行曝光,觀察結果。
2 結果與分析
2.1 重組表達質粒pET32a-p22 的構建
用p22-F/p22-R 引物對優化的p22基因進行擴增,結果顯示,PCR 擴增后的p22基因片段約為447 bp,大小符合預期(圖1)。 通過酶切酶連將p22擴增產物連接到pET-32a 質粒中。 經測序分析后,基因測序結果完全正確。
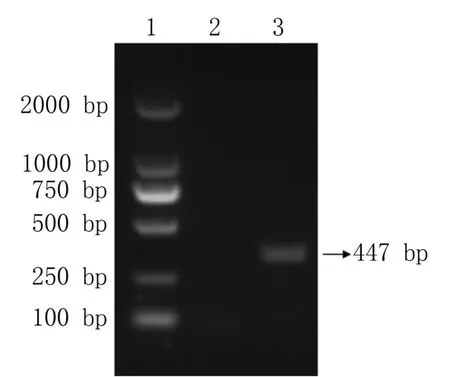
圖1 p22 基因PCR 擴增目的片段
2.2 重組p22 蛋白的表達和純化
SDS-PAGE 電泳結果顯示,與未誘導組相比,誘導組可見與預期大小一致的37 kDa 融合蛋白,經超聲破碎后可溶性分析發現,重組p22 蛋白條帶出現在超聲后上清中,表明重組p22 蛋白為可溶性表達(圖2A)。 進一步大量誘導p22 重組蛋白的表達,并與His Ni-NTA Agarose 親和層析柱結合,使用不同濃度梯度的咪唑洗脫液洗脫蛋白,收集洗脫液,隨后進行SDS-PAGE 電泳鑒定。 結果顯示,使用高濃度洗脫液時,在37 kDa 處出現單一條帶(圖2B),純化后純度大于95%。
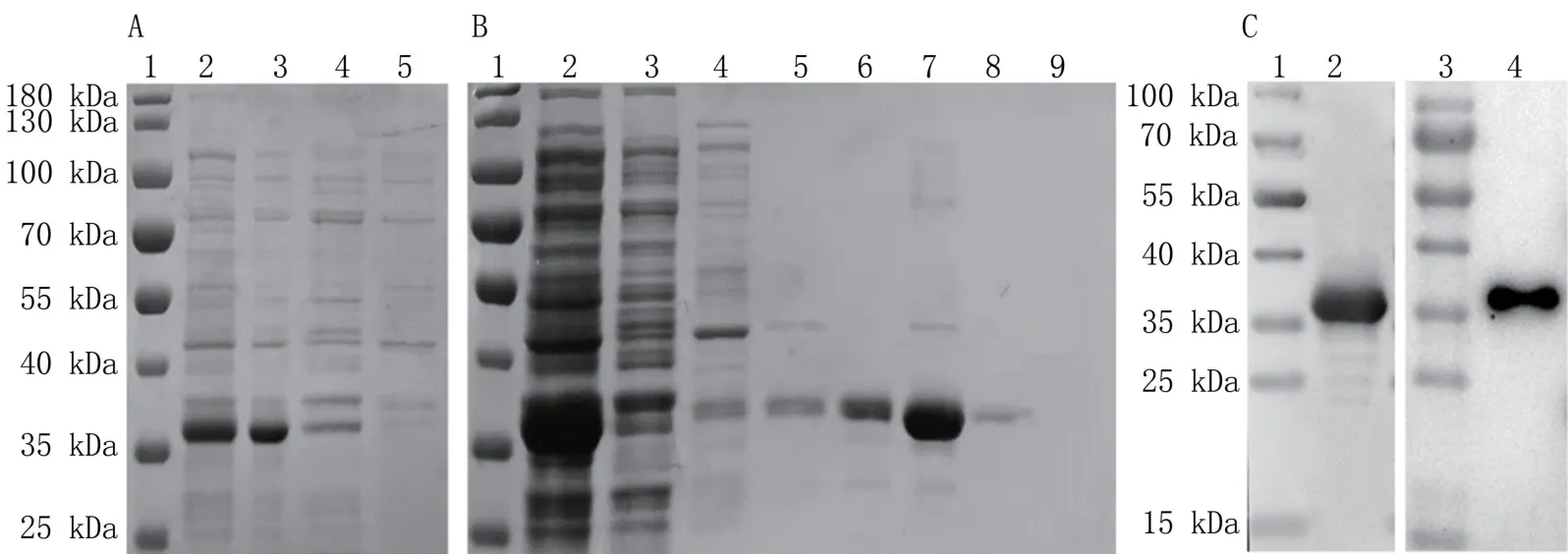
圖2 重組p22 蛋白的表達(A)、純化(B)及鑒定(C)
2.3 重組p22 蛋白的Western blot 鑒定
對重組p22 蛋白進行Western blot 鑒定,通過His 標簽抗體孵育后,在37 kDa 處檢測到嵌合His標簽的p22 蛋白;而使用ASFV 陽性血清孵育后,發現重組p22 蛋白能夠與ASFV 陽性血清發生特異性結合(圖2C),表明該抗原具有良好免疫原性。
2.4 單克隆抗體的制備及鑒定
2.4.1 血清效價的測定 三免7 d 后通過間接ELISA 方法檢測接種小鼠血清效價,測得1、2、3號小鼠的血清效價為1∶128 000,4、5 號小鼠的血清效價為1∶64 000(圖3A)。 選取1、2、3 號小鼠進行加強免疫。
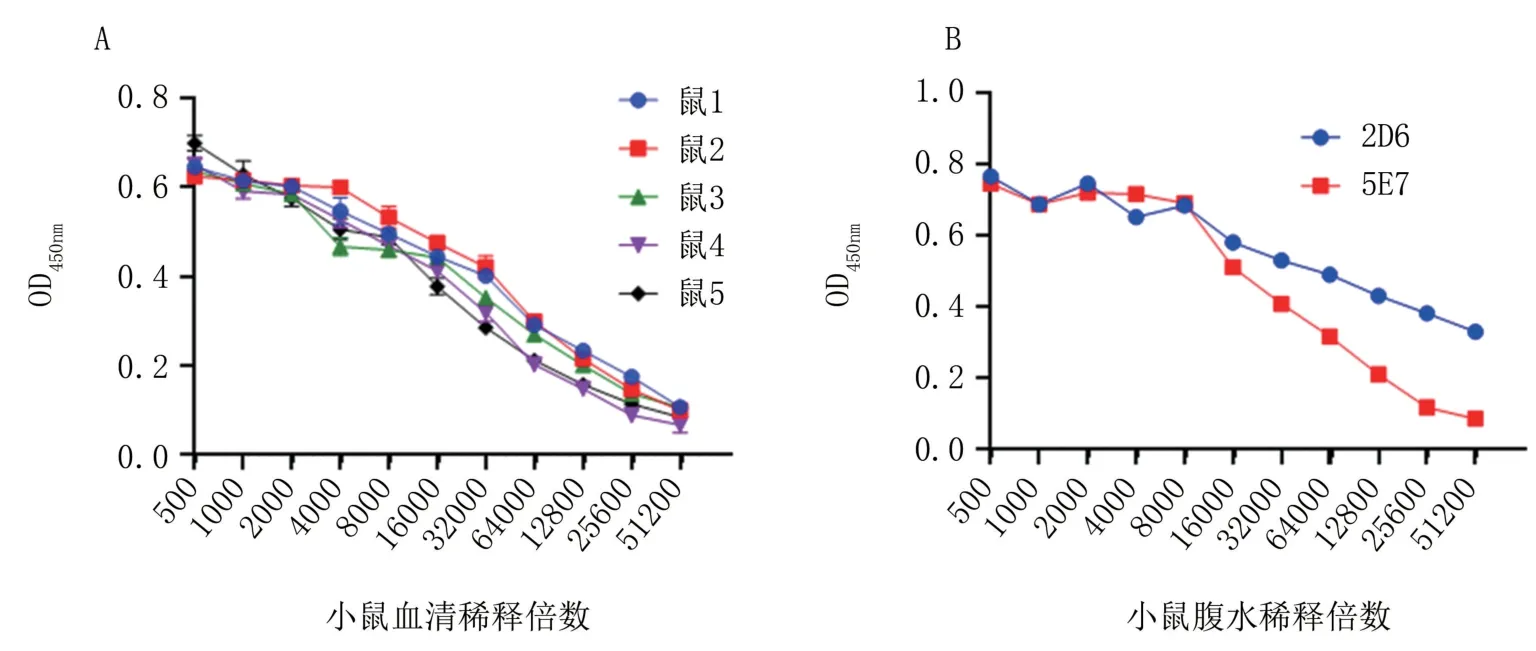
圖3 小鼠血清效價(A)及單克隆抗體效價(B)的測定
2.4.2 單克隆抗體效價測定 在骨髓瘤細胞和脾細胞融合后,進行融合篩選,對篩選的陽性孔進行3 次亞克隆后,經間接ELISA 方法檢測,最終篩選到了兩株能夠穩定分泌重組p22 MAb 的雜交瘤細胞株,通過小鼠腹腔注射兩種雜交瘤細胞株,制備了小鼠腹水,通過間接ELISA 方法測定腹水的效價分別為1∶512 000 和1∶256 000(圖3B),將獲得的單克隆抗體命名為2D6 和5E7。
2.4.3 單克隆抗體亞型鑒定 采用美國Southern Biotech 公司的單抗亞型鑒定試劑盒分別對p22 MAb 2D6 和5E7 的亞型進行檢測,結果顯示兩種MAb 的亞型均為IgG1(表1)。
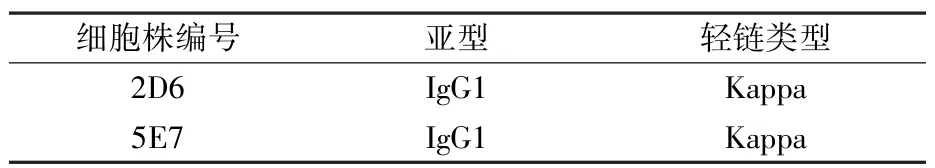
表1 單克隆抗體亞型鑒定
2.4.4 單克隆抗體的免疫反應鑒定 為了鑒定MAb 2D6 和5E7 的免疫反應性,通過Western blot檢測了其對外源性p22 蛋白的反應性。 結果顯示,兩株MAb 均能夠在37 kDa 處與p22 蛋白發生特異性反應(圖4)。
3 討論與結論
自2018 年ASF 傳入中國并迅速傳播以來,ASF 對生豬生產造成了重大的經濟威脅,目前,在中國傳播的ASFV 類型主要為一種高毒力的基因型Ⅱ毒株[16],基因型Ⅰ毒株也有報道[17]。 ASFV在環境中具有高傳染性和穩定性,可通過與受感染的豬或其產品直接接觸或通過軟蜱進行傳播[18]。 直到目前,仍然沒有針對ASFV 的有效商業疫苗,使得非洲豬瘟的預防、控制和治療變得十分艱難[19],因此,篩選ASF 免疫原性蛋白,鑒定免疫相關基因,將有助于ASF 疫苗的開發。 另外,需要對ASFV 的結構功能等基礎研究進行攻克,以期為ASF 疫苗或診斷試劑的研發提供更多理論依據。 相關研究表明,鑒定最具抗原性的病毒蛋白對改進血清學診斷試驗具有重要作用,在ASFV 血清診斷試驗中使用重組蛋白作為抗原來源可避免操縱活病毒的潛在風險[20]。 p22 蛋白為ASFV 的結構蛋白,根據序列分析比較,東歐等地區ASFV 分離株中的p22 序列保守,證明p22蛋白是抗原穩定的,因此適合用作開發新的血清診斷試驗的工具[21]。 目前對于p22 蛋白的功能研究尚不完善,其生物學功能仍待闡明。
原核表達系統具有操作簡單、表達量高的特點,本研究中所用的原核表達載體pET-32a 可在BL21(DE3)細菌中高度表達,并且可以在裂解物的上清液中檢測到大量目標蛋白,大大簡化了純化過程,同時避免了由包涵體的變性和復性可能引起的活性損失[22]。 雖然從構象結構上看,原核表達與真核表達的抗原會有差異,但不會影響抗原的線性表位[23],因此,使用原核表達系統制備單克隆抗體仍可繼續進行線性表位的篩選,為ASFV 結構功能的研究提供生物學材料。
單克隆抗體(MAb)是診斷病毒感染的關鍵試劑。 與多克隆抗體相比,MAb 在現代免疫診斷的應用中表現出特異性強、靈敏度高等優勢,在疾病預防、診斷、治療和流行病學調查等研究中發揮了巨大作用。 張馮禧等[24]通過原核表達系統獲得ASFV p30 重組蛋白并制備了MAb,效價為1∶102 400,表明該蛋白有良好的免疫原性。 齊艷麗等[25]通過原核表達系統表達純化了ASFV p54重組蛋白并制備了MAb,抗體ELISA 效價均高于1∶25 600 且特異性良好。
本研究通過大腸桿菌表達系統成功表達并純化出一種可溶性重組蛋白p22,并制備了兩株針對該蛋白的MAb。 經SDS-PAGE 和Western blot共同分析,重組蛋白大小約為37 kDa,將純化的重組蛋白做免疫原免疫小鼠,得到抗體的ELISA效價不低于1∶256 000,說明制備的MAb 具有良好的免疫原性,為p22 蛋白的結構功能等基礎研究以及ASFV 免疫類診斷試劑產品的開發提供了重要的生物材料。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中老年保健(2021年3期)2021-08-22 06:50:04
昆明醫科大學學報(2021年1期)2021-02-07 01:06:36
現代臨床醫學(2021年1期)2021-01-26 00:56:02
中華養生保健(2020年4期)2020-11-16 01:31:40
中西醫結合肝病雜志(2020年2期)2020-10-27 02:18:50
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
現代檢驗醫學雜志(2014年4期)2014-02-02 02:44:59
