植物響應低溫的生長發(fā)育及分子機制研究進展
2023-11-14 08:44:58吳宇欣蔡昌楊唐詩蓓謝裕紅王曉艷
江蘇農(nóng)業(yè)科學 2023年19期
吳宇欣, 蔡昌楊, 唐詩蓓, 謝裕紅, 王曉艷, 朱 強
(1.福建農(nóng)林大學林學院,福建福州 350000; 2.福建省將樂縣林業(yè)局,福建將樂 353300;3. 福建省三明市林業(yè)科技推廣中心,福建三明 353000)
植物在生長過程中,經(jīng)常會受到多種非生物脅迫的影響,例如冷、熱、干旱、重金屬和鹽脅迫等;這些非生物脅迫會限制植物的地理分布范圍并降低其生產(chǎn)力[1]。低溫脅迫對植物影響巨大,會破壞植物的質膜結構,降低植物光合作用能力,過量累積活性氧(ROS),使植物的生長發(fā)育出現(xiàn)遲緩甚至停滯等現(xiàn)象[2-4]。
在漫長的演變進化過程中,植物形成了一系列復雜高效的網(wǎng)絡調(diào)控機制,來應對低溫帶來的威脅。隨著分子生物學技術的不斷發(fā)展,植物應對寒冷脅迫的機制已逐漸明晰[5]。在生理生化水平上,植物生成了可溶性糖、脯氨酸、多胺類化合物等一系列滲透調(diào)節(jié)物質,來穩(wěn)定細胞膜結構和清除活性氧[6-7]。另外,大量蛋白激酶和轉錄因子在冷應激信號通路中發(fā)揮著作用[8-9]。CBF/DREB1(C-Repeat Binding Factor/Dehydration-Responsive Element-Binding Protein 1)是其中最關鍵的調(diào)節(jié)因子[10]。在冷脅迫下,CBF基因受上游的ICE1(Inducer of CBF Expression 1)誘導而大量表達,催化冷調(diào)節(jié)(COR)基因表達來調(diào)節(jié)冷應激[11]。與CBF(C-Repeat Binding Factor)依賴型信號途徑相比,非CBF依賴型信號途徑也發(fā)揮著重要作用。例如,sfr6(sensitivetofreezing6)基因在不誘導CBF基因的情況下,直接調(diào)控下游COR基因的表達[12]。同時,冷脅迫相關基因在轉錄、翻譯和翻譯后修飾方面也發(fā)揮著關鍵作用[13]。本文對近年來植物響應低溫脅迫調(diào)控機制的相關研究進行綜述,并概述目前利用基因工程手段提高經(jīng)濟作物抗寒性的研究進展,以期為更深入地研究植物低溫脅迫響應機制提供理論基礎,并為培育抗寒能力優(yōu)秀的新品種奠定一定基礎。
1 植物對低溫脅迫的生理生化響應
1.1 冷馴化現(xiàn)象
低溫脅迫包括冷害(0~20 ℃)和凍害(<0 ℃),對植物的正常生長發(fā)育具有不良影響[14]。冷害是熱帶及亞熱帶地區(qū)植物遭遇的主要脅迫方式,嚴重影響水稻、玉米、大豆等農(nóng)作物的生產(chǎn);而凍害則主要影響溫帶地區(qū)植物,對小麥、油菜等作物產(chǎn)生不利影響[15]。為了應對低溫脅迫帶來的不良影響,一些物種在不良環(huán)境中逐漸提高自身的耐凍能力,這種現(xiàn)象被稱為冷馴化[15]。植物在冷馴化過程中,會重新調(diào)節(jié)光合作用過程和碳水化合物代謝途徑,對轉錄和蛋白質進行重新調(diào)控,修復細胞損傷或死亡造成的失衡[16]。同時,植物體內(nèi)生成一些關鍵的滲透調(diào)節(jié)物質,例如可溶性糖、脯氨酸、可溶性蛋白、多胺類化合物等,用來抵抗脫水現(xiàn)象并減少冷應激對細胞的損害[17-21]。過氧化物酶(POD)、過氧化氫酶(CAT)、超氧化物歧化酶(SOD)、抗壞血酸過氧化物酶(APX)、谷胱甘肽過氧化物酶(GPX)等一些抗氧化物酶也在植物體內(nèi)大量累積,以減輕活性氧(ROS)引起的過氧化損傷[19,22]。此外,植物也通過調(diào)節(jié)一些激素代謝途徑,例如增加赤霉素、細胞分裂素、生長素、茉莉酸等一些激素的水平,來增強植物對非生物脅迫的耐受性[23-24]。
1.2 低溫對植物的生理影響
植物在受到低溫脅迫后,其體內(nèi)的各項生理生化指標和細胞器形態(tài)會發(fā)生很大改變[25]。在低溫脅迫下,植物許多細胞的生理生化功能會隨著細胞膜及脂質的變化而變化,如細胞內(nèi)離子的泄露、氨基酸的流失、各種細胞成分的結構變化,最終會導致植物壞死或死亡[26]。
首先,遭受低溫影響時,植物的表型會發(fā)生改變。當植物受到低溫脅迫時,氣孔開始閉合,致使CO2滯留在氣孔腔中,無法與外界進行交換,從而影響光合作用的進行;同時,參與蔗糖和淀粉生物合成的某些關鍵酶受到損傷,例如核酮糖-1,5-二磷酸羧化酶(RuBisCO)發(fā)生失活,直接抑制線性電子傳輸并限制光合產(chǎn)物的數(shù)量[27]。此外,低溫直接影響葉綠體的結構、功能和發(fā)育。當植物暴露在低溫下時,參與卡爾文循環(huán)(Calvin Cycle)的2種關鍵酶景天庚酮糖-1,7-二磷酸酶(SBPase)、果糖-1,6-二磷酸酶(FBPase)的活性明顯降低[3]。這些結果說明,低溫通過影響植物的呼吸作用和化合反應中關鍵酶的合成,使植物的光合作用受到限制,最終導致植物生長發(fā)育緩慢,植株矮化,葉片萎蔫,根系生長受損,作物結實率低,產(chǎn)量下降。
其次,受低溫脅迫影響期間,植物細胞中可溶性糖的水平會有所增加。可溶性糖在植物體內(nèi)起滲透保護劑的作用,當其在細胞中積累時,滲透濃度增加,致使低溫引起的細胞收縮效應減輕,從而起到保護細胞膜免受脫水和冷凍損害的作用[28]。在滲透調(diào)節(jié)方面,脯氨酸也發(fā)揮了重要作用。脯氨酸是一種在植物中廣泛存在的氨基酸,具有獨特的細胞功能,不僅參與植物的發(fā)育與代謝,還在防御和脅迫耐受性方面發(fā)揮重要作用[29]。植物處于低溫環(huán)境中會產(chǎn)生大量的脯氨酸,可以清除產(chǎn)生的過量氫離子(H+)以維持有氧呼吸的平衡,并穩(wěn)定多聚核糖體來提高蛋白質的親和性,進而提高植物抵御不良環(huán)境的能力[30]。總之,這些滲透調(diào)節(jié)物質通過維持細胞內(nèi)滲透水平的平衡,來抵抗低溫脅迫帶來的不良影響。
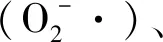
最后,低溫脅迫會導致植物細胞膜的損壞。低溫環(huán)境中,植物組織中開始形成冰晶[34]。首先是細胞外空間開始結冰導致滲透壓下降,進而使細胞脫水,最終導致細胞質膜結構異變和電解質泄漏增加[35]。隨著低溫的持續(xù)刺激,細胞最終破裂,細胞質流出體外,細胞質膜損傷,最終導致植物死亡。
2 植物低溫脅迫響應的分子機制研究
在長期的生長發(fā)育過程中,植物進化出一系列復雜的網(wǎng)絡調(diào)控機制來應對低溫脅迫,主要包括CBF依賴型信號途徑和非CBF依賴型信號途徑。
2.1 CBF依賴型機制
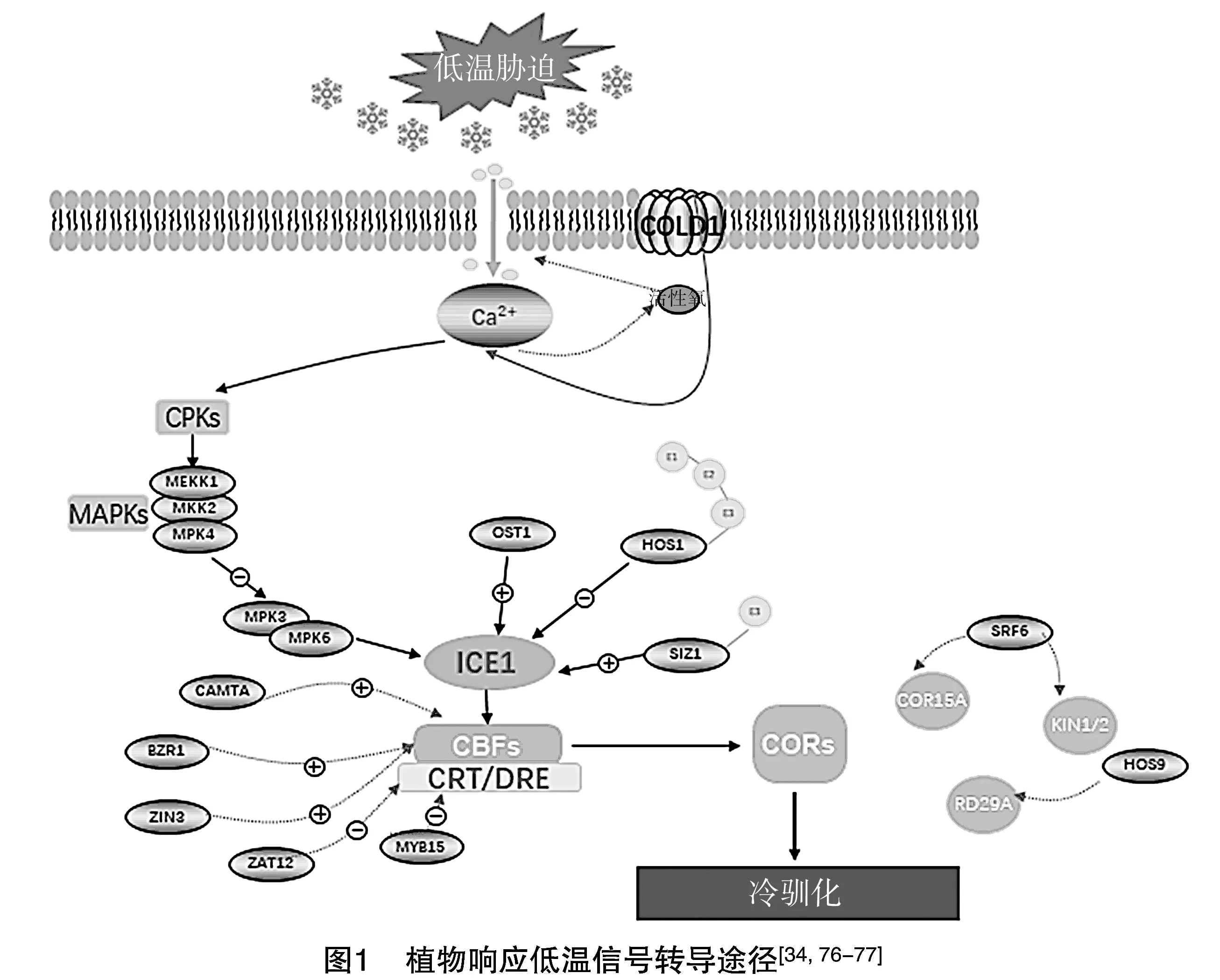
目前,已有大量研究探索了植物對寒冷的調(diào)節(jié)機制。其中,CBF途徑是植物應對低溫脅迫最關鍵的調(diào)控途徑[36]。而ICE1(Inducer of CBF Expression 1)-CBF(C-Repeat Binding Factor)-COR(Cold Regulated)信號級聯(lián)通路,是當中最重要的低溫信號通路[37]。在低溫脅迫條件下,轉錄因子ICE1與CBF基因的啟動子區(qū)域結合,激活CBFs的表達,被激活后的CBFs進一步激活下游COR基因的表達,以此增強植物的抗寒性[38]。
2.1.1 ICE1-CBF-COR調(diào)控途徑 調(diào)節(jié)低溫脅迫信號最典型的轉錄因子是C重復結合因子/脫水響應元件結合蛋白(C-Repeat Binding Factors/Dehydration Responsive Element Binding Protein,CBF/DREB),它屬于在植物生長發(fā)育及非生物脅迫反應方面發(fā)揮作用的AP2/ERF超蛋白家族ERF家族中的一個亞家族,僅包含1個AP2結構域[39]。其成員CBF1、CBF2、CBF3(DREB1B、DREB1C、DREB1A)已被證實參與低溫脅迫反應過程,當植物處于低溫條件下能夠在短時間內(nèi)被誘導[40]。研究證明,3種CBF蛋白有著非常高(86%)的序列相似性,雖然都是受冷響應的誘導,但這3種CBF蛋白還存在著功能冗余[41]。CBF1、CBF3蛋白在功能上更相似,而CBF2蛋白在表達模式上相較于其他2個CBF有所不同。在發(fā)育早期,CBF1、CBF3會在植物的根、下胚軸、子葉中特異性表達;而CBF2則是在下胚軸、子葉中表達,在根中無法表達[42]。有研究證明,在CBF2突變體中,CBF1、CBF3基因的表達量增加,表現(xiàn)出對冷凍的耐受性增加,這表明CBF2是CBF1、CBF3表達的負調(diào)節(jié)因子[43]。
CBF基因通過其AP2/ERF DNA結合域,與下游靶基因啟動子中存在的CRT/DRE順式元件(A/GCCGAC)結合,并激活下游COR基因的轉錄以增加耐寒性[44]。冷調(diào)節(jié)(COR)基因在受到調(diào)節(jié)后,會生成一些抗凍多肽、轉錄因子、蛋白激酶、與胚胎發(fā)生和脂質代謝相關的蛋白質以及與激素反應和細胞壁修飾有關的蛋白質等,這些產(chǎn)物可以幫助植物在低溫脅迫下穩(wěn)定細胞膜的結構,從而增強其耐寒性[45]。
在冷響應級聯(lián)通路的上游,存在bHLH轉錄因子家族的成員ICE1(Inducer of CBF Expression 1),它是一種MYC型堿性螺旋-環(huán)-螺旋轉錄因子,在C端含有保守的bHLH結合結構域,用以與下游的CBF3/DREB1A啟動子中的典型MYC順式元件(CANNTG)結合,正向誘導CBF3/DREB1A調(diào)節(jié)子的表達,而CBF1、CBF2基本上不受影響[46]。而ICE1的同源物ICE2與ICE1在序列上有高度一致性,編碼相同的bHLH結構域序列。ICE1、ICE2均為氣孔形成的主要調(diào)節(jié)因子,并且在調(diào)節(jié)冷反應過程中發(fā)揮重要作用[47]。目前研究表明,ICE1、ICE2存在著功能冗余,在誘導CBF基因方面具有不同的方式。ICE1、ICE2通過誘導CBF2來分別調(diào)節(jié)CBF3、CBF1的表達,降低植物的冷凍耐受性[48]。
2.1.2CBF轉錄調(diào)控的正調(diào)控作用 低溫脅迫響應過程涉及一系列轉錄途徑,一些組成型表達的轉錄因子能夠在對寒冷的反應中被激活表達。試驗證明,這些轉錄因子會正向誘導下游CBF基因的轉錄。例如,ICE1是CBFs的正調(diào)節(jié)因子,它能夠與下游基因CBF3啟動子中的MYC識別位點結合,來調(diào)控CBF3表達。ice1突變體中CBF3的表達能力降低,與野生型相比,該突變體耐寒性大大降低,不能進行冷馴化[46]。過表達ICE1則大大提高了CBF1、CBF2、CBF3、COR基因的表達,增強了植物的抗凍性[49]。參與信號傳導的鈣調(diào)蛋白結合轉錄激活因子(CAMTA)的轉錄因子家族也是CBF的正調(diào)控因子,會對溫度快速下降做出反應[13]。其具有一個特異性DNA結合活性的CG-1結構域,能夠正向調(diào)節(jié)CBF2啟動子中的CM2元件,強烈誘導CBF1、CBF2的表達[50]。油菜素唑耐受因子1(Brassina-Zole-Resistant1,BZR1)是油菜類固醇信號通路中的關鍵轉錄因子,與靶基因啟動子中的BRRE(CGTGT/CG)和E-box(CACGTG和CACTTG)基序結合,促進CBFs、WRKY6、WRKY54以及脫落酸(ABA)受體PYL6等編碼基因的表達,正向調(diào)控植物的耐寒性[51]。同時,BZR1的蛋白質水平和磷酸化狀態(tài)依賴油菜素內(nèi)脂(BR)信號傳導中上游的GSK3樣激酶BIN2(BR-Insensitive 2)的影響,在沒有BRs的情況下,具有活性的BIN2磷酸化BZR1并促進其降解,增加植物的冷凍敏感性[52]。
2.1.3CBF轉錄調(diào)控的負調(diào)控作用 除了正向調(diào)控以外,信號通路中還存在多個降低植物抗凍性的CBFs負調(diào)節(jié)因子。MYB轉錄因子家族成員MYB15被證實能夠與ICE1發(fā)生作用,并與CBF啟動子元件結合。在冷脅迫下,MYB15表達上調(diào),過表達MYB15顯示CBF基因轉錄水平降低,而在MYB15突變體中顯示CBF3、CBF1、CBF2水平增加,表明MYB15參與了CBF基因的冷調(diào)節(jié)過程,降低了植物的低溫脅迫耐受性[53]。C2H2鋅指蛋白基因ZAT12(ZincFingerTranscriptionFactor12)通過抑制15個冷響應基因和激活9個冷響應基因的表達,來調(diào)節(jié)冷馴化。ZAT12還下調(diào)CBFs基因的表達,表明它在植物適應低溫脅迫中起負調(diào)控作用[54]。乙烯信號通路中的轉錄因子EIN3(Ethylene-Insensitive3)會對耐寒性產(chǎn)生負面影響。EIN3蛋白與CBF3啟動子中特定的結合基序結合,阻止其轉錄,同時細胞分裂素信號通路中的A型調(diào)節(jié)因子ARR5、ARR7、ARR15被證實是EIN3的下游靶基因。在過表達EIN3的轉基因植物中,ARR5、ARR7、ARR15的轉錄和蛋白穩(wěn)定性受到明顯抑制,說明EIN3通過聯(lián)合乙烯和細胞分裂素信號傳導途徑,在冷應激反應中發(fā)揮拮抗作用[55]。
2.2 非CBF依賴型機制
CBF及其靶基因對于冷脅迫信號至關重要,但仍然發(fā)現(xiàn)有一些基因是不依賴CBF途徑的,它們功能的增加或缺失不會影響CBF基因功能的表達,但會使植物的抗凍能力升高或下降。例如,HY5(ElongatedHypocotyl5)是一種參與光信號傳導的bZIP類轉錄因子,當受到低溫脅迫影響時,E3泛素連接酶COP1(Constitutively Photomorphogenic 1)促進HY5的穩(wěn)定表達,通過非CBF依賴型途徑正向調(diào)節(jié)下游COR基因的表達[56]。SFR6(SensitivetoFreezing6)基因被證實通過CRT/DRE啟動子序列基序調(diào)節(jié)下游的COR基因;在冷凍敏感突變體sfr6中,COR基因包括KIN1/2、LTI78、COR15A等的表達水平顯著降低[12];而在sfr6突變體中,CBF1、CBF2、CBF3的冷誘導表達則不受影響[57]。HOS9(HighExpressionofOsmoticallyResponsiveGene9)作為一種組成表達的基因,通過冷誘導表達,會使RD29A和其他一些COR基因的轉錄水平明顯高于野生型;而在突變體hos9-1中,CBF轉錄因子基因的表達沒有受到影響,卻仍表現(xiàn)出冷凍應激的敏感性[58]。
2.3 轉錄后調(diào)控
低溫脅迫在植物中誘導廣泛的轉錄后和翻譯后修飾(PTM),對生成mRNA的質量和數(shù)量造成影響,使基因功能發(fā)生改變,最終影響植物的低溫脅迫耐受性。轉錄后調(diào)控在冷馴化過程中起著至關重要的作用。例如,STA1(Stabilized1)作為mRNA前體的剪接因子,參與mRNA的剪切以去除內(nèi)含子,使基因得以正常表達。而sta1-1突變體在冷誘導COR15A基因的剪接中存在缺陷,使得COR15A基因無法正常表達,降低了植物的耐寒性,這說明STA1的表達對植物的耐寒性起著關鍵作用[59]。核孔蛋白NUP160(Nucleoporin 160)在控制RNA核胞質轉運中發(fā)揮重要作用。擬南芥nup160突變體中CBF3的表達受到抑制,并且破壞細胞核的mRNA輸送過程,說明NUP160在植物生長、開花時間調(diào)節(jié)和低溫脅迫耐受性中起重要作用[60]。滲透反應因子LOS4(Low expression of osmotically responsive gene 4)編碼DEAD-box RNA 解旋酶,該解旋酶參與RNA的代謝過程,LOS4突變體會負向調(diào)控CBFs表達,降低RD29A和其他COR基因的表達,從而負調(diào)節(jié)植物的抗寒性,說明LOS4是CBF基因的正調(diào)節(jié)因子,并且在基因調(diào)控和植物耐寒性方面發(fā)揮關鍵作用[61]。
2.4 翻譯后調(diào)控
在翻譯后修飾(PTM)過程中,幾個基因與ICE、CBF、COR基因相互作用,以改變其活性、構象、定位和穩(wěn)定性。磷酸化、泛素化和SUMO偶聯(lián)是植物調(diào)節(jié)低溫脅迫反應途徑的主要PTM[62]。植物受到冷脅迫時,ICE1受到翻譯后修飾,進而影響CBF或其他下游基因的表達。
2.4.1 磷酸化修飾 磷酸化在植物冷適應中起重要作用,是一種可逆的蛋白質修飾,對激酶和磷酸酶具有高度依賴性[11]。最常見的磷酸酶OST1(Open Stomata 1)/SnRK2.6/SRK2E是 ABA 信號傳導中的絲氨酸/蘇氨酸 (Ser/Thr)蛋白激酶,可以磷酸化ICE1基因,幫助其增加轉錄穩(wěn)定性。同時,OST1蛋白還通過干擾HOS1蛋白與ICE1蛋白的結合,來抑制HOS1介導的ICE1降解,雙重作用下增強CBF基因的活性,來緩解冷應激效應[63]。除OST1外,絲裂原活化蛋白激酶(Mitogen-Activated Protein Kinase,MAPK)級聯(lián)也通過磷酸化作用參與植物低溫脅迫反應。MAPK是一組能被細胞因子、神經(jīng)遞質、激素、細胞應激及細胞黏附等細胞外刺激激活的絲氨酸-蘇氨酸蛋白激酶。其級聯(lián)過程由MAPK激酶激酶(MAP Kinase Kinase Kinase,MAPKKK)、MAPK激酶(MAP Kinase Kinase,MAPKK)和MAPK組成[64]。在植物感知外界信號刺激后,首先MAPKKKs磷酸化并激活MAPKKs,激活的MAPKKs隨后磷酸化MAPKs,最后活化的MAPKs磷酸化特定的下游底物,如轉錄因子、激酶或其他酶等,共同調(diào)控植物的生長和發(fā)育、生物和非生物脅迫反應以及植物激素信號轉導等過程[65]。響應寒冷的級聯(lián)包括MEKK1、MKK2、MPK4和(或)MPK6激酶[66]。受到冷脅迫后MEKK1被上調(diào),MEKK1激活下游的MKK2,MKK2的直接下游靶標MPK4被MKK2特異性磷酸化和激活,同時MPK3、MPK6也因冷處理而被激活[67]。活化的MPK3/MPK6級聯(lián)通路通過在Ser94、Thr366、Ser403位點處磷酸化而促進ICE1的降解,從而減少CBF基因的轉錄來實現(xiàn)負調(diào)控,而MEKK1-MKK2-MPK4級聯(lián)則是正向調(diào)節(jié)冷反應,并會對MPK3、MPK6激酶活性造成抑制[68]。
2.4.2 泛素化和SUMO化修飾 泛素化是指泛素分子在一系列特異性酶作用下,將細胞內(nèi)的蛋白質進行分類,選中特定的靶蛋白,對其進行特異性修飾的過程[69]。泛素化修飾涉及泛素激活酶E1、泛素結合酶E2、泛素連接酶E3所連接的一系列酶促反應。其中,E3-泛素連接酶通過與靶分子相互作用并為泛素化反應提供支架而起最重要的作用[70]。HOS1(High Expression of Osmotically Responsive Gene 1)是一種功能性無名指蛋白,具有泛素E3連接酶活性,包含一個變異型RING指結構域,靶向ICE1進行泛素化介導的蛋白質降解,負調(diào)控CBF的表達,最終降低植物的耐寒性[71]。
另外一種蛋白翻譯后的修飾方式為SUMO化修飾。SUMO分子通過在E1活化酶、E2結合酶、E3連接酶級聯(lián)與靶蛋白偶聯(lián),調(diào)控靶蛋白的結構與功能[72]。而SUMO修飾蛋白SENPs與SUMO分子共同調(diào)節(jié)受體蛋白的SUMO化狀態(tài),特異性地對底物蛋白進行去SUMO化修飾,進而改變細胞功能[73]。目前,SUMO化修飾在控制細胞周期、維持基因組完整性、控制亞細胞運輸和調(diào)控轉錄機制等方面發(fā)揮著重要作用[74]。SIZ1(SUMO ligase)是一種SUMO E3連接酶,介導與ICE1的偶聯(lián),在低溫下增強ICE1的穩(wěn)定性,有助于誘導CBF3/DREB1A的表達,正向調(diào)節(jié)植物的耐寒性[75]。
基于以上對植物響應低溫脅迫信號相關研究進行的綜述,歸納出植物響應低溫脅迫信號轉導途徑的分子機制模式圖(圖1)。
3 利用基因工程改良植物抗冷性的相關研究進展
受溫室效應的影響,極端天氣頻頻發(fā)生。在農(nóng)業(yè)生產(chǎn)過程中,農(nóng)林類作物常常遭受低溫侵襲,生理發(fā)育過程遭到破壞,作物生長不良,最終造成結實率低、產(chǎn)量下降甚至植株死亡。隨著全球人口的不斷增加,人們對各種作物產(chǎn)品特別是糧食的需求越來越大[78]。利用基因工程手段改良植物的抗逆性,并培育抗寒能力優(yōu)越的品種,收獲更多的作物產(chǎn)量,對農(nóng)業(yè)發(fā)展及社會經(jīng)濟增長具有重要意義。
研究表明,在水稻中過表達bZIP73、bZIP71,二者形成異二聚體,可以抑制ABA生物合成,促進從花藥到花粉的可溶性糖轉運,從而提高水稻在生育期的抗寒性,最終提高結實率和籽粒產(chǎn)量[79]。過表達DREB1A的轉基因水稻,可以增加脯氨酸和各種糖類等滲透保護劑的含量,提高水稻對干旱、高鹽、低溫脅迫的耐受性[80]。水稻中表達R2R3-MYB轉錄因子家族中的OsMYB2則增強植物對鹽、低溫、脫水脅迫的耐受性,并提高種子萌發(fā)率,積累更多的可溶性糖和游離脯氨酸,以促進滲透調(diào)節(jié),提高水稻抗寒性[81]。過表達NAC轉錄因子家族成員SNAC2基因,則可以幫助水稻增強細胞膜的穩(wěn)定性,提高對滲透脅迫的耐受性,并生成一系列過氧化物酶、鳥氨酸氨基轉移酶、賴氨酸酮戊二酸還原酶、重金屬相關蛋白、鈉/氫交換劑、熱休克蛋白、GDSL樣脂肪酶等產(chǎn)物,最終使植株出現(xiàn)對寒冷、鹽、脫水脅迫耐受性顯著提高的表型[82]。研究表明,核小GTP酶RAN2在植物體內(nèi)主要起介導細胞核中GTP和細胞質中GDP交換的作用,因此在水稻中過表達RAN2基因,可以促進植物細胞核內(nèi)微管蛋白的正常輸出,從而維持穩(wěn)定的細胞分裂,最終提高水稻的抗寒性[83]。
研究表明,在玉米中過表達ZmbZIP4,與野生型相比,轉基因植物的側根數(shù)量明顯增加,植物的主根變得更長,其根系得到改善,并且生成了許多應激反應基因和脫落酸合成相關基因來增加植物抵抗非生物脅迫的能力[84]。過表達ZmMYB31基因則提高了玉米體內(nèi)相關冷應激基因的表達,并且降低了在冷脅迫中造成的離子外滲、ROS含量、低溫光抑制現(xiàn)象,從而在寒冷和過氧化物脅迫中發(fā)揮正向調(diào)控作用[85]。ZmICE1基因能夠抑制Glu/Asn生物合成基因天冬酰胺合成酶的表達,來減少ROS的產(chǎn)生,并可以直接調(diào)控DREB1的表達,且ZmICE1啟動子與正調(diào)節(jié)因子ZmMYB39結合,均可顯著增加玉米的抗寒性;過表達ZmICE1對玉米產(chǎn)量相關性狀沒有明顯的負面影響,表明過表達ZmICE1基因既可培育耐寒玉米新品種,又不會對玉米產(chǎn)量造成影響[86]。
有關轉基因小麥和大麥的研究進展表明,從小麥的HD-ZipⅠ基因中克隆出HDZI-3、HDZI-4啟動子,成功優(yōu)化轉基因小麥和大麥中TaCBF5L、TaDREB3基因在非生物脅迫下的表達,可以提高轉基因植物的抗寒性,并減少轉基因對植物發(fā)育和籽粒產(chǎn)量的負面影響[87]。克隆出水稻的WRKY71基因啟動子和小麥的Cor39基因啟動子優(yōu)化TaDREB3的表達,構建大麥轉基因株系;與野生型相比,過表達TaDREB3明顯提高了大麥的抗凍性,并提高了冷響應基因的表達量,同時沒有對植物特性和籽粒產(chǎn)量產(chǎn)生負面影響[88]。從小麥中分離出DREB/CBF基因TaRAP2.1L,作為一種脅迫響應型轉錄抑制因子,生成的TaRAP2.1L突變體植株可以激活DREB/CBF下游基因的表達,從而增強小麥抗凍和抗干旱能力,同時沒有對小麥的生長和生產(chǎn)造成負面影響[89]。
在番茄中過表達冷調(diào)節(jié)基因LeCOR413PM2(Cold-Regulated413-PlasmaMembrane2),可減輕植物細胞膜的損傷,降低植物體內(nèi)ROS的積累和PSⅡ的光抑制現(xiàn)象,而且可以維持體內(nèi)抗氧化酶的高活性和滲透調(diào)節(jié)物質的含量,從而提高轉基因番茄植株的耐寒性[90]。HY5-MYB15-CBFs轉錄級聯(lián)反應已被研究證實在番茄的冷反應中起重要作用,當在番茄中過表達HY5或MYB15基因時,均能激活CBF1、CBF2、CBF3的表達,從而增強番茄作物的耐寒性[91]。在番茄中過表達油菜類固醇受體SlBRI1(BrassinosteroidInsensitive1),則可以降低MDA、ROS的積累,提高SOD、POD、CAT的活性。并且在過表達SlBRI1的轉基因植株中,ICE1、CBFs相關基因表達量明顯增高,說明SlBRI1通過影響ICE1-CBF-COR途徑的轉錄水平,并正向影響ROS清除系統(tǒng)、光系統(tǒng)的光抑制以及植物激素的生物合成和信號傳導,來增加番茄對寒冷壓力的耐受性[92]。而在番茄中過表達SlDREB3,可以幫助番茄在冷應激下減輕ROS積累和因冷凍引起的細胞損傷,與此同時,還可通過激活SlLEA基因的表達,提高轉基因品系對低溫脅迫的耐受性[93]。
4 總結與展望
低溫脅迫作為一種主要的非生物脅迫,對植物影響巨大,是影響全球農(nóng)林作物生長及生產(chǎn)的主要因素之一;對相關農(nóng)作物進行耐低溫能力研究,對于糧食生產(chǎn)及社會的可持續(xù)發(fā)展具有重要意義。到目前為止,植物的耐寒機制研究已經(jīng)取得重大突破,其中ICE1-CBF-COR信號級聯(lián)通路在植物抗寒方面起著至關重要的作用。本綜述總結了植物在耐寒性方面所發(fā)揮的多重調(diào)控作用,包括CBF依賴型信號途徑和非CBF依賴型信號途徑,并著重介紹了關鍵轉錄因子CBF在植物感知低溫信號過程中發(fā)揮的功能及其依賴于CBF信號通路對于植物平衡生長、發(fā)育和對低溫脅迫響應方面的所發(fā)揮的作用。目前,基因工程技術已廣泛應用于改良植物性狀,將外源基因通過轉化插入植物的基因組中,定向改變植物的功能性狀,同時克服了傳統(tǒng)育種的周期長、效率低等弊端[94]。本文歸納了多個農(nóng)業(yè)作物物種(水稻、小麥、番茄等)利用基因工程技術在抗寒性方面已經(jīng)取得的成就,旨在為今后改良植物抗性研究提供一定基礎。
由于低溫脅迫影響植物從體外到細胞的各個方面,形成了錯綜復雜的信號網(wǎng)絡,需要從細胞、蛋白、代謝等水平對其進行研究,因此仍需要進一步研究揭示冷馴化過程中植物通過感知環(huán)境變化來調(diào)整自身發(fā)育的機制。隨著溫室效應的逐年增強,極端環(huán)境出現(xiàn)的頻率越來越高,因此對植物進行更多的抗寒性研究,提高植物抗寒冷的能力,進而培育出抗凍能力優(yōu)越的植物新品種和種質資源,仍具有非常重要的意義。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
中國生殖健康(2019年3期)2019-02-01 06:12:26
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
海軍航空大學學報(2015年3期)2015-11-11 17:20:00
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
