雙組分調節系統對肺炎克雷伯菌多黏菌素耐藥機制影響的研究進展
2023-11-09 07:02:58楊文麗王東亮馮鈞帥齊保立史惠文
數理醫藥學雜志 2023年10期
楊文麗,王東亮,馮鈞帥,陳 莉,齊保立,史惠文,袁 媛
1. 甘肅中醫藥大學第一臨床醫學院(蘭州 730000)
2. 甘肅省人民醫院重癥醫學科(蘭州 730000)
肺炎克雷伯菌(Klebsiella pneumoniae, KP)是一種重要的人類病原體,被歸類為一種常見的機會性醫院相關細菌,涉及多種感染,包括尿路感染、肺部感染、肝膿腫和手術傷口感染[1]。在過去十年中,全球范圍內耐多藥KP 分離株的報告顯著增加。
多黏菌素(多黏菌素B 和黏菌素)是于1947年從多黏菌芽孢桿菌中分離出的一種具有抗革蘭氏陰性菌活性的多陽離子抗菌肽[2]。在20 世紀80 年代,由于多黏菌素的神經毒性和腎毒性,其應用受到限制。21 世紀初,隨著多重耐藥(multi-drug resistance, MDR)微生物數量的增加,尤其是碳青霉烯耐藥菌的出現,多黏菌素再次受到臨床的關注[3]。多黏菌素是治療KP 感染的最后手段藥物,然而臨床中出現了不同程度的耐碳青霉烯類肺炎克雷伯菌(carbapenem-resistantKlebsiella pneumoniae, CRKP)耐多黏菌素的情況。2022 年中國細菌耐藥監測報告數據顯示,CRKP 對多黏菌素B 的耐藥率為7.9%,對黏菌素的耐藥率為8.2%[4]。研究顯示,雙組分調節系統(two-component regulatory systems, TCSs)可通過感知環境變化調節KP 對多黏菌素耐藥的修飾[5]。KP 可通過TCSs 修飾脂多糖(lipopolysaccharide,LPS)、mgrB 負反饋調節因子、外排泵等介導多黏菌素耐藥。
1 細菌雙組分調節信號轉導的基礎
典型的TCSs 由一對蛋白質組成,包括一個傳感器組氨酸激酶(histidine kinase, HK)和一個反應調節因子(response regulator, RR)。可變HK n 端結構域感知環境變化,通過結合細胞外配體或通過其他構象變化,三磷酸腺苷(adenosine triphosphate, ATP)水解觸發保守的c 端組氨酸殘基的自磷酸化。HK 結合的磷酸被轉移到RR 保守的n 端結構域的天冬氨酸殘基上,RR 是一種位于細胞質中的同型二聚體蛋白,激活RR 的可變c 端輸出結構域,允許其通過靶向DNA 來調節基因組表達。通過磷酸化的信息流,細菌能夠有效感知其周圍環境的變化(營養、pH 值、滲透壓、抗生素等),并以一種允許對動態環境進行快速反應的方式協調基因表達[5]。
2 TCSs對多黏菌素耐藥的影響機制
細菌細胞外膜(outer membrane, OM)的陰離子性質是由LPS 的存在所決定的,它包含一個帶負電荷的脂質A 部分。LPS 是多黏菌素的初始靶點,多黏菌素可與LPS 的脂質A 區域的磷酸基相互作用[2,6]。一旦附著,多黏菌素就會通過取代二價陽離子Ca2+和Mg2+來破壞OM,導致細胞死亡。
當暴露在陽離子抗菌肽,如多黏菌素B和黏菌素下,細菌會通過改變LPS 保護自己免受不利環境的刺激。暴露在陽離子抗菌肽下激活了細菌的TCSs,從而導致磷酸乙醇胺(phosphoethanolamine, PEtN)和4-氨基阿拉伯糖(4-aminoarabinose, L-Ara4N)陽離子基團的合成和轉移,對LPS 的脂質A 進行共價修飾[7]。這些LPS 共價修飾可以中和OM 上的負電荷,使OM 帶正電荷,阻礙多黏菌素與OM 的結合,導致抗生素作用的減少或消失,從而形成多黏菌素耐藥[8]。
3 PhoP/PhoQ和PmrA/PmrB對肺炎克雷伯菌的耐藥機制
在介導KP 對多黏菌素耐藥中最典型的兩個TCSs 是PhoP/PhoQ 和PmrA/PmrB。研 究 發 現,在暴露于多黏菌素的KP 中,PhoP/PhoQ 和PmrA/PmrB 系統表達上調[9]。Naha 等[10]的研究顯示,在多黏菌素耐藥的KP 大多數菌株的TCSs 基因表達上調,包括PhoP(4~6 倍)、PhoQ(5~9 倍)、PmrA(4~13 倍)和PmrB(6~11 倍)。
PhoP/PhoQ 和PmrA/PmrB 的激活是由環境刺激和TCSs 內的特定突變觸發的,這些突變導致它們的構成激活和隨后的過表達[11-12]。
3.1 PhoP/PhoQ系統
PhoP/PhoQ 系統是KP 中研究最廣泛的TCSs 系統,其在許多致病性和非致病性細菌中較為保守。
在各種致病菌中,PhoP/PhoQ 具有感知宿主細胞內信號并在感染期間調節細菌生活方式的能力[13]。反應調節因子PhoP 在許多革蘭氏陰性菌中高度保守,傳感器激酶PhoQ 位于細胞內膜(intracellular membrane, IM),可通過自磷酸化對幾種環境變化作出反應,包括低pH 值、低Mg2+濃度和抗生素的存在,然后磷酸基團被轉移到反應調節因子PhoP,它激活了參與LPS 修飾的下游基因的表達,通過遺傳調控對OM 進行修飾[14-17]。
PhoP/PhoQ 系統的激活,主要受到mgrB基因突變的負反饋系統調節作用的影響,PhoP/PhoQ系統通過PmrD 間接激活PmrA/PmrB 系統來促進多黏菌素抗性[18-20]。mgrB的插入失活與PhoP/PhoQ 系統和pmrHFIJKLM 操縱子的過表達有關,PhoP/PhoQ 系統一旦被激活,磷酸化的PmrA 就會結合到pmrCAB 操縱子和pmrHFIJKLM 操縱子的啟動子區域,增加RNA 聚合酶的識別和結合,并導致操縱子的上調,介導PEtN 與L-Ara4N 的合成和轉移到脂質A[21]。
多黏菌素對易感表型抗性的逆轉與PhoP和PhoQ基因的非同義突變相關。在智利的一項研究中,ST25 中PhoP 的104 位氨基酸替換、ST1161 中PhoQ 的A351D 氨基酸替換在黏菌素異質性耐藥分離株中被檢測到[22]。另外,PhoP基因的L26Q 突變可導致黏菌素耐藥[23-24]。羅馬尼亞的一項研究提示PhoP基因的L4F 突變、PhoQ基因的L26Q、Q426L、L224Q、Q317K 突變也會增加黏菌素的抗性[25]。此外,Halaby 等[26]在黏菌素異質性耐藥(colistin heteroresistant, CST-HR)和產生超廣譜β-內酰胺酶(extended-spectrum betalactamase, ESBL)的KP 分離物中發現了PhoQ 中的A21S 替代,使黏菌素最低抑制濃度(minimum inhibitory concentration, MIC) 從2 μg/mL 增 加 到16 μg/mL,表明該氨基酸替代在CST-HR 的表型中發揮了重要作用。
mgrB基因編碼一個小的跨膜蛋白是PhoP/PhoQ 系統的負調控因子。mgrB的突變導致PhoP/PhoQ 雙組分系統上調,進而導致pmrCAB和pmrHFIJKLM 操縱子的過表達[27-29]。pmrCAB和pmrHFIJKLM 操縱子的上調歸因于磷酸化的PmrA,PmrA 被PmrD 激活,而PmrD 反過來又被mgrB破壞導致PhoP 磷酸化激活[9,30]。此外,激活的PhoP 也可直接激活KP 中的pmrHFIJKLM操 縱 子[31]。與TCSs 突 變(PmrA/PmrB 或PhoP/PhoQ)相比,mgrB突變/失活導致KP 對多黏菌素耐藥的作用更大[24]。這一結果表明,mgrB在KP 的多黏菌素耐藥性中起著重要作用。印度的一項研究發現,IS903 樣和ISKpn26 樣插入元件造成的mgrB的破壞導致KP 對黏菌素敏感性降低[32]。
有研究稱IS5 樣插入元件是KP 中mgrB破壞的主要元素,但ISKpn14(IS1 家族成員)的插入也偶有報道[33-34]。Morales-León 等[22]的研究發現IS5 樣和IS1 樣轉座子插入序列使mgrB失活,降低了mgrB的mRNA 水平,從而影響多黏菌素的抗性。然而,沙特阿拉伯的一項研究發現存 在ISKpn14、ISKpn28、IS903 導 致mgrB基 因破壞,而ISKpn14 是主要的IS,59%的分離株存在mgrB基因破壞是由ISKpn14 介導的[23]。在羅馬尼亞的一項研究中,ISL3(ISKpn25)和IS5(ISKpn26)在不同的核苷酸位置破壞mgrB基因,增加CPKP 的黏菌素抗性[25]。另外,生物信息學工具預測mgrB基因M27K 突變,提示同樣是有害的。相比之下,Liu 等[35]研究發現,MgrB 蛋白的表達不受M27K 突變的影響。
在Naha 等[10]的研究中,mgrB中的氨基酸替換(C28G,V32G)導致突變是有害的,影響主要功能域,可能調節蛋白活性,有助于黏菌素抗性的增加。mgrB的失活也被證明可以通過抑制宿主防御反應的啟動和限制多種抗菌肽的作用來增強KP 分離株的毒力[36]。
3.2 PmrA/PmrB系統
KP 的多黏菌素耐藥也歸因于PmrA/PmrB基因的突變,該基因是另一個編碼控制L-Ara4N 和PEtN 合成的雙組分調節系統[22,37-38]。
在大腸桿菌和腸道沙門氏菌中,PmrB 作為一種傳感器細胞質膜結合激酶,被高價鐵(Fe3+)和酸性環境(pH=5.5)激活。激活后,它通過磷酸化激活PmrA[39]。在體內,PmrA/PmrB 參與感知環境,幫助細菌在體內侵襲上皮細胞、在細胞內生長、細胞間擴散、誘導巨噬細胞凋亡,并且與細菌毒力表達相關[40]。PmrA或PmrB基因的突變通常會導致PmrA 的結構性激活,可以上調pmrCAB 操縱子和pmrHFIJKLM 操縱子,pmrCAB操縱子參與PEtN 對脂質A 修飾,而pmrHFIJKLM操縱子編碼的酶負責合成和將L-Ara4N 轉移到脂質A[40]。總的來說,PmrA 的這種構成性激活導致了更多正電荷的LPS,從而降低了帶正電荷的多黏菌素的親和力。
PmrB基因的突變已在具有耐藥性或敏感性降低的KP 分離株中被廣泛描述。一項研究顯示,CRKP 中PmrB基因P95L、T157P、R256G、V352E 突變與黏菌素的耐藥性增加相關[25]。此外,Jayol 等[41]描述了6 個黏菌素耐藥菌株的PmrB 中一個單一的T157P 氨基酸取代,而這種取代會破壞PmrB 蛋白的α-螺旋二級結構,并連續激活PmrA,從而產生PmrC 的過表達,最終導致黏菌素抗性。另一項研究在ST25 和ST11 中也發現了PmrB 的相關氨基酸(分別是P95L 氨基酸和A256G 氨基酸)取代導致KP 對黏菌素的耐藥性增加[22]。而Liu 等[35]的研究顯示,PmrB 中的氨基取代D313N 使黏菌素MIC 提高到8 mg/L,比原菌株高16 倍,而PmrB 中的M285L 和PmrA 中的S204L 并沒有增加黏菌素對原菌株的MIC 值。
Wand 等[42]的研究在PmrA中只發現了G53C突變導致黏菌素MIC 增加到64 mg/L。目前,關于PmrA基因突變導致多黏菌素耐藥性增加的研究相對較少。
PmrA/PmrB 和PhoP/PhoQ 的突變和構成性激活可能共同導致pmrCAB 操縱子和pmrHFIJKLM操縱子的激活,從而導致多黏菌素耐藥性[2,8]。與多黏菌素抗性相關的PhoP/PhoQ 和PmrA/PmrB 調控關系見圖1。
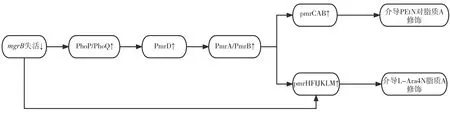
圖1 與多黏菌素抗性相關的PhoP/PhoQ和PmrA/PmrB調控圖Figure 1. Regulatory plots of PhoP/PhoQ and PmrA/PmrB associated with polymyxin resistance
4 其它TCSs對肺炎克雷伯菌的耐藥機制
進化實驗表明,KP 中多黏菌素耐藥性也可能由于CrrB基因的突變而出現[43]。CrrA/CrrB 雙組分系統通過PmrAB 調控網絡參與了對pmrHFIJKLM 和CrrC 操縱子的控制,而CrrB 調控pmrHFIJKLM 操縱子的表達是通過CrrC 操縱子介導的[44]。研究發現,臨床CrrB 突變導致L-Ara4N和PEtN 同時添加到脂質A 中,誘導更高的多黏菌素抗性[45]。Naha 等[10]的研究也提示,CrrA/CrrB突變導致KP 對黏菌素產生耐藥性。在羅馬尼亞的一項研究中,CrrB基因罕見的P151S 替換,使異質性耐藥KP 的黏菌素MIC >64 mg/L[25]。另一項研究已證實GrrB基因突變可誘導KP 對黏菌素的耐藥性升高,且該研究發現CrrB基因Q10L、Y31H、W140R、N141I 和S195N 錯義突變均會引起KP 對黏菌素的MIC 值增加64~1024 倍[44]。類似地,Jayol 等[46]報道了一個具有細微差異的突變,CrrB中的P151L 賦予了黏菌素耐藥性,而Pitt 等[47]證實了同一基因中的P158R 替換。此外,CrrA/CrrB 可激活PmrA,而CrrB的功能獲得突變可激活導致黏菌素耐藥性的基因表達,而不受PmrA/PmrB 系統的影響[48-49]。
KP 編碼另外兩種TCSs 為CpxAR 和PhoBR,它們抑制KpnO 孔蛋白,降低了對多種抗生素的敏感性,包括氯霉素、阿米卡星、萘啶酸和四環素[50-51]。兩個TCSs 都由于OM 在應激狀態下而激活。此外,在KP 中CpxA/CpxR 可同時上調三個MDR RND 家族外排泵以響應OM 應激[50]。
TCSs 對多種細菌的生存發揮著關鍵作用,它為細菌提供了感知和反應周圍環境的不可或缺的工具。鑒于TCSs 在細菌穩態中的重要作用以及可通過它們激活的多種抗生素耐藥性機制,TCSs是開發新的抗菌療法的一個較好靶點[52-53]。使用預測軟件來識別結合一個或多個TCSs 以使其無效的假定化合物的能力,將是藥物靶向TCSs 的一個重要工具[54]。例如PmrB基因,71%的耐藥性非同義突變發生在Hamp(存在于組氨酸激酶、腺苷酸環化酶、甲基接受蛋白和磷酸酶)連接子和DHp(二聚化和組氨酸磷酸轉移)結構域。這些結果增強了研究人員對支持多黏菌素耐藥性的調控機制的理解,并可能有助于開發新的策略來減少耐藥性的出現[55]。此外,有研究顯示,與對碳青霉烯類抗生素敏感的菌株相比,CRKP 中OmpK35/36 孔蛋白減少與EnvZ-OmpR、PhoPQ 和BaeSR 等TCSs 的下調有關[56]。
5 結語
耐多藥KP 是一項全球性的公共衛生問題。本文通過闡述TCSs 導致KP 對多黏菌素耐藥的機制,將TCSs 設想為未來可能的抗菌靶點,為規劃抗擊KP 耐藥性的策略提供依據,拓寬耐藥菌研究的視野,從而為未來的研究提供更多思路,并為臨床上更加有效的治療耐多藥KP 感染提供參考。
猜你喜歡
工業設計(2022年8期)2022-09-09 07:43:20
保健醫苑(2022年5期)2022-06-10 07:46:38
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
軍民兩用技術與產品(2021年10期)2021-03-16 06:05:30
北京測繪(2020年12期)2020-12-29 01:33:58
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
家庭影院技術(2017年9期)2017-09-26 03:41:45
