氧化處理對黑珍珠和赤霞珠葡萄酒顏色及花色苷穩定性的影響
2023-11-08 06:10:38程思琦李生保曲措洛松次仁茶靜國游義琳黃衛東戰吉宬
食品與發酵工業 2023年20期
程思琦,李生保,曲措,洛松次仁,茶靜國,游義琳,黃衛東,戰吉宬*
1(中國農業大學 食品科學與營養工程學院,北京,100083)2(北京市葡萄科學與釀酒技術重點實驗室,北京,100083) 3(西藏芒康藏東珍寶酒業有限公司,西藏 昌都,401121)
顏色是決定葡萄酒品質的重要因素之一,可以反應葡萄原料和葡萄酒的狀況,對品嘗者的感官偏好有重要影響。花色素及其衍生物是葡萄酒呈色的主要物質,其核心結構是3,5,7 -三羥基-2-苯基苯并吡喃陽離子,花色素由于缺乏電子而不穩定,導致其在自然界中通常以糖基化的花色苷形式存在[1]。
我國釀酒葡萄品種多樣,主要以赤霞珠、霞多麗等歐亞種葡萄(Vitisvinifera)為主。黑珍珠(又名藏葡萄)是西藏芒康地區廣泛種植的特色葡萄品種,由法國傳教士于18世紀中葉引入該地區,經過100多年的自然馴化,已十分適應當地的風土條件。黑珍珠葡萄產量高,抗性強,不易感染真菌病害,用其為原料釀制的葡萄酒具有顏色絢麗、呈紫紅色的特點。但在研究過程中發現,黑珍珠葡萄酒的顏色在陳釀過程中容易氧化降解,對其色澤品質有較大影響。我們推測黑珍珠葡萄酒顏色容易氧化降解與其花色苷組成有關。
不同品種的葡萄花色苷組成不同,歐亞葡萄品種中5-O-葡萄糖苷轉移酶失活[2],無法合成3,5-O-雙糖花色苷,葡萄酒顏色主要由單糖花色苷提供;但該酶在美洲葡萄(Vitislabrusca)、山葡萄(Vitisamurensis)[3]、河岸葡萄(Vitisriparia)[4]、圓葉葡萄(Vitisrotundifolia)[5]、刺葡萄Vitisdavidii[6]等非歐亞種葡萄中成功表達,因此山葡萄V.amurensis[3]等通常含有較多的雙糖花色苷。李貝貝等[7]通過微衛星技術分析認為西藏黑珍珠可能為‘巴柯’(歐美雜交種,原產地法國,由‘Folle Blanche’與‘Grand Glabre’雜交)的同物異名品種。作為歐美雜交種,黑珍珠葡萄中可能含有大量的雙糖花色苷。近年來關于雙糖花色苷的研究主要集中在檢測不同品種葡萄及葡萄酒花色苷含量的差異[8-10],以及探究模擬體系中雙糖苷的輔色效率[11]和聚合能力[12]方面,我國鮮有研究者關注雙糖花色苷對紅葡萄酒顏色穩定性的影響。
因此,本研究利用CIELab參數法對比在與氧氣接觸過程中黑珍珠葡萄酒與赤霞珠葡萄酒的顏色變化情況,并利用高效液相色譜-質譜聯用技術(ultra-high performance liquid chromatography-mass spectrometry,UPLC-MS/MS)分析葡萄酒花色苷的組成及含量,以期進一步探究雙糖花色苷在葡萄酒中的穩定性,并為黑珍珠葡萄酒的開發利用提供參考。
1 材料與方法
1.1 材料與試劑
葡萄酒樣品由西藏芒康藏東珍寶酒業有限公司提供,2021年西藏鹽井產區黑珍珠原酒[酒精度(15.54±0.17)%,殘糖(2.77±0.03) g/L]和赤霞珠原酒[酒精度(15.7±0.43)%,殘糖(1.84±0.07) g/L]。
花青素-3-O-葡萄糖苷(色譜純),成都曼斯特生物技術有限公司;乙腈(色譜純),阿拉丁生物技術公司;濃HCl、氯化鈉、醋酸、醋酸鈉,均為分析純,上海麥克林生化科技有限公司。
1.2 儀器與設備
V1800可見分光光度計,尤尼科(上海)儀器有限公司;ACQUITY UPLC I-class/Xevo TQS高效液相色譜質譜聯用美國,Waters公司。
1.3 實驗方法
1.3.1 葡萄酒氧化
將150 mL黑珍珠和赤霞珠葡萄酒分別置于200 mL 錐形瓶中,覆蓋3層紗布,置于20 ℃培養箱控溫保藏,進行氧化。每天取樣,分析測定,每組酒樣設置3組平行。
1.3.2 色度和色差測定
參照李運奎等[13]的方法進行測定。
1.3.3 CIELab參數的測定
參照PéREZ-CABALLERO等[14]方法進行測定。酒樣經0.45 μm濾膜過濾,置于0.2 cm光程石英比色皿中(用蒸餾水作為參比),測定供試酒樣在450、520、570、630 nm的吸光度,計算出三色值X、Y、Z值,繼而得CIELab參數L*、a*、b*。
1.3.4 總花色苷的測定
采用pH示差法對葡萄酒中總花色苷的含量進行測定[15]。
1.3.5 單體花色苷的測定
翻開迪慶工行的史冊,幾十年間發生的變化都歷歷在目,它見證了改革開放以來工行發展的艱辛歷程,這是一段值得銘記的歷史。
采用UPLC-MS/MS進行葡萄酒花色苷的檢測,BEH C18色譜柱(100 mm×2.1 mm,1.7 μm),柱溫40 ℃,進樣體積5 μL,流速0.2 mL/min。流動相A:1%甲酸的水,流動相B:1%甲酸的乙腈。洗脫梯度:0~0.5 min 5% B;0.5~10 min 5%~23% B,10~13 min 46% B,13~13.5 min 5% B;13.5~15 min 5% B。
質譜采用電噴霧離子源(ESI),正離子模式,離子掃描范圍:100~1 000m/z;霧化器壓力:35 psi;干燥氣流速:10 L/min;干燥氣溫度:325 ℃。葡萄酒樣品過0.22 μm濾膜后直接上樣。花色苷的定性定量分析參照中國農業大學葡萄酒研究中心建立的“葡萄與葡萄酒花色苷HPLC-UV-MS/MS指紋譜庫”[16]。
1.4 數據處理
采用Microsoft Office Excel 2003統計數據,SPSS 24.0進行顯著性數據分析,Origin 2021軟件繪圖,Adobe Photoshop軟件進行顏色匹配。
2 結果與分析
2.1 飽和度和色差值的變化
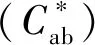
a-飽和度;b-色差值圖1 黑珍珠和赤霞珠葡萄酒暴露在氧氣條件下6 d色度和色差值的變化Fig.1 Changes in saturation and chromatic aberration of Black Pearl and Cabernet Sauvignon wine exposed to oxygen for 6 days注:不同小寫字母代表差異顯著(P<0.05)(下同)。
2.2 CIELab參數的變化
在CIELab體系中,顏色參數(L*)代表物質明暗程度,L*數值越大,表示顏色亮度越高,顏色越淺。圖2顯示,黑珍珠葡萄酒的亮度值低于赤霞珠,在接觸氧氣的過程中,黑珍珠的亮度值逐漸上升,顏色變淺;而赤霞珠的亮度值逐漸下降,顏色加深。

a-L*,亮度;b-a*,紅色色調;c-b*,黃色色調圖2 黑珍珠和赤霞珠葡萄酒暴露在氧氣條件下6 d CIELab參數的變化Fig.2 CIELab changes in Black Pearl and Cabernet Sauvignon wine exposed to oxygen for 6 days
b*代表葡萄酒的黃藍程度,b*>0時,顏色偏黃,b*<0時,顏色偏藍。黑珍珠葡萄原酒的黃色色調值高于赤霞珠葡萄酒,在接觸氧氣0~6 d的過程中,黑珍珠葡萄酒的黃色色調值顯著上升,上升64.15%;而赤霞珠葡萄酒的黃色色調值變化不大。
2.3 總花色苷含量的變化
黑珍珠和赤霞珠葡萄酒在與氧氣接觸的過程中總花色苷的變化情況如圖3所示,隨接觸時間的延長,黑珍珠葡萄酒的總花色苷含量持續下降,從初始的297.49 mg/L下降至158.86 mg/L,下降了46.51%;赤霞珠葡萄酒的總花色苷含量變化不大,從初始的142.42 mg/L下降至135.64 mg/L,下降了4.76%。氧氣對黑珍珠葡萄酒總花色苷的降解作用顯著大于赤霞珠葡萄酒。
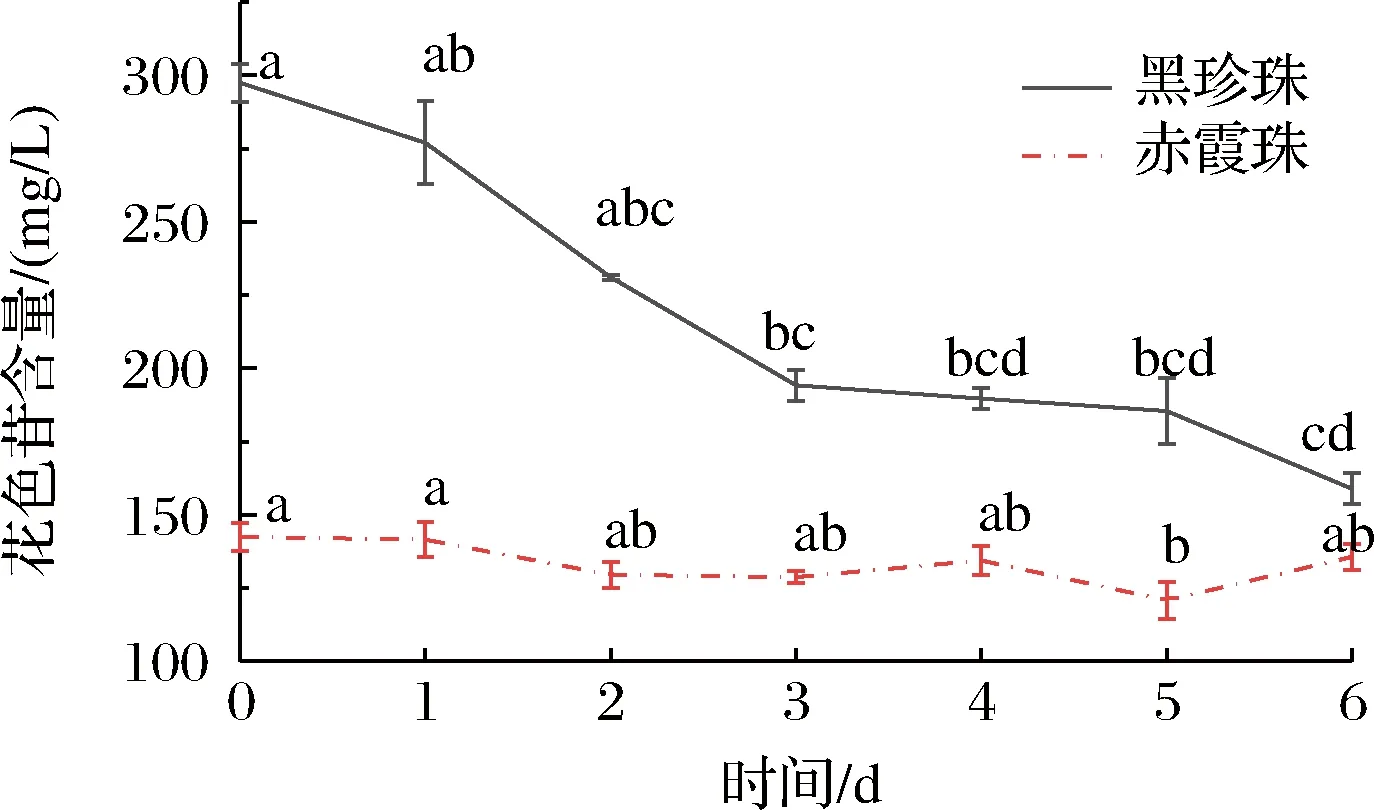
圖3 黑珍珠和赤霞珠葡萄酒暴露在氧氣條件下6 d總花色苷含量的變化Fig.3 The changes of total anthocyanins in Black Pearl and Cabernet Sauvignon during the 6 days oxygen explosion
2.4 赤霞珠和黑珍珠葡萄酒中花色苷的組成
在黑珍珠和赤霞珠葡萄酒中共檢測到28種花色苷,其中黑珍珠葡萄酒中檢測到21種花色苷,包括5種未酰化的雙糖花色苷,7種酰化的雙糖花色苷,5種未酰化的單糖花色苷,4種酰化的單糖花色苷;赤霞珠葡萄酒檢測到19種花色苷,包括3種未酰化的雙糖花色苷,5種未酰化的單糖花色苷,11種酰化的單糖花色苷,未檢測到酰化的雙糖苷。
赤霞珠葡萄酒的顏色主要由單糖花色苷提供,占花色苷總量的99.37%,但也檢測到了少量的甲基花青素-3,5-O-葡萄糖苷(0.35 mg/L)和二甲基花翠素-3,5-O-葡萄糖苷(1.89 mg/L),此前,歐亞種葡萄中含有少量的雙糖花色苷的現象已被研究者證實[18]。黑珍珠葡萄酒中花色苷總量更高,花色苷組成主要以雙糖花色苷為主(646.89 mg/L),占花色苷總量的85.47%,尤其以甲基花青素-3,5-O-葡萄糖苷(322.97 mg/L)和二甲基花翠素-3,5-O-葡萄糖苷(153.21 mg/L)含量豐富。這主要與葡萄的品種有關,黑珍珠葡萄為歐美雜交種[7],美洲葡萄和歐美雜交種葡萄酒通常含有較高比例的雙糖花色苷[4-5]。
由表1和圖4可知,赤霞珠葡萄酒中含有較高比例的酰化單糖花色苷,特別是二甲花翠素-3-O-(6-O-乙酰化)-葡萄糖苷(123.91 mg/L),花色苷的酰化比例高達36.31%,而黑珍珠葡萄酒中的花色苷主要以非酰化形式存在,僅含有6.91%的酰化雙糖花色苷和0.76%的酰化單糖花色苷,其中主要以二甲基花翠素3-O-(6-O-香豆酰化)-葡萄糖苷-5-O-葡萄糖苷(30.77 mg/L)為主。這主要是因為美洲種和歐美雜交種的葡萄中大部分酰基轉移酶失活,造成葡萄酒中花色苷的酰化程度一般低于赤霞珠等歐亞種葡萄[19]。
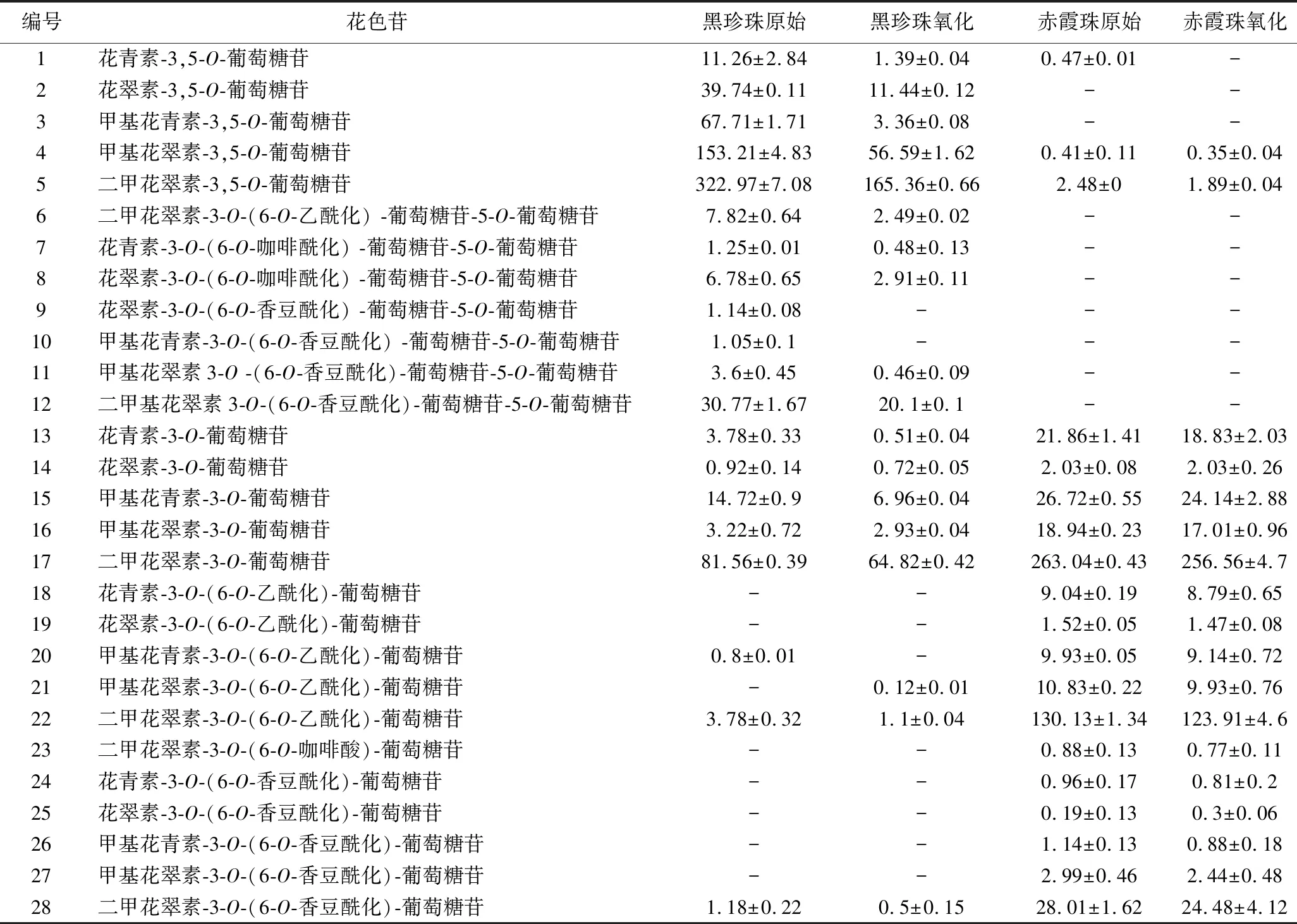
表1 葡萄酒中單體花色苷的組成情況 單位:mg/L
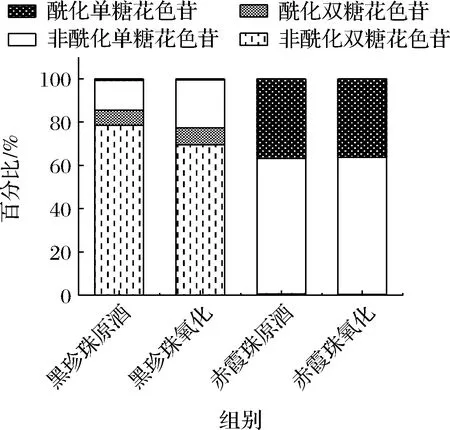
圖4 黑珍珠與赤霞珠葡萄酒酰化花色苷的比例Fig.4 The proportion of acetylated anthocyanins in Black Pearl and Cabernet Sauvignon
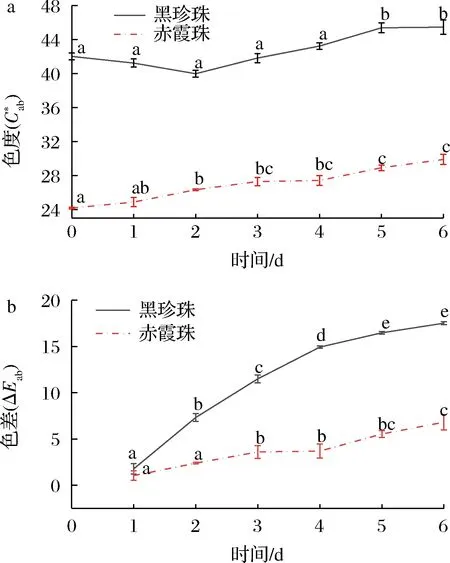
葡萄酒與氧氣接觸后,單體花色苷的變化情況與圖3總花色苷的變化趨勢大體一致,都呈現下降趨勢。氧化6 d后,與赤霞珠葡萄酒相比(總花色苷下降5.24%,雙糖花色苷下降33.3%,單糖花色苷下降5.06%),黑珍珠葡萄酒的花色苷總量減少54.78%,其中雙糖花色苷降低59.10%,單糖花色苷降低29.39%,花色苷的降解率明顯高于赤霞珠葡萄酒。黑珍珠和赤霞珠葡萄酒中雙糖花色苷的降解率都顯著高于單糖花色苷,這與葡萄酒中雙糖花色苷易氧化降解并產生棕色物質[20-21]的報道一致,因此黑珍珠葡萄酒中高比例的雙糖花色苷是導致氧化過程中花色苷含量迅速下降的重要原因。此外,酰化率較低也是黑珍珠顏色不穩定的原因之一,經過氧化黑珍珠葡萄酒未酰化雙糖花色苷的占比由78.56%下降至69.59%,而酰化雙糖花色苷的占比略有上升,由6.91%上升至7.73%。
3 討論
取代基、糖苷化和酰化等結構是影響單體花色苷穩定性的重要因素。其中,雙糖苷取代可以抑制查爾酮形式花色苷開環裂解成醛和酚酸,因此,模擬葡萄酒[22]和花色苷的提取液[23]中雙糖花色苷的穩定性高于單糖花色苷,然而在葡萄酒中雙糖花色苷容易與酒中的物質發生褐化反應,加速葡萄酒顏色的老化,但該雙糖花色苷氧化物的具體結構尚不清晰[5, 21]。
在葡萄酒與氧氣的接觸過程中,單體花色苷易與葡萄酒中的黃烷-3-醇等多酚類物質反應,形成聚合花色苷[12, 24],除Portisin型以外的吡喃花色苷為橙紅色,非吡喃聚合花色苷大多表現出紫紅色的色調[25],穩定性均顯著高于相應的單體花色苷。本研究中,赤霞珠的總花色苷下降,但紅色色調上升,可能正是因為生成了聚合花色苷。然而,黑珍珠葡萄酒中存在的大量雙糖花色苷由于5號碳位被糖苷取代,不僅無法環化形成吡喃花色苷,也極大的降低了非吡喃花色苷的聚合效率[26],因此,雙糖花色苷在葡萄酒中難以形成穩定的聚合花色苷也是其穩定性弱于單糖花色苷的重要原因。然而,根據本試驗結果,單糖花色苷在黑珍珠葡萄酒中的降解率也高于赤霞珠葡萄酒,這可能是因為雙糖花色苷的降解產物具有強還原性[27],加速了單糖花色苷的氧化。
酰化也是影響花色苷穩定性的重要因素,酰化花色苷可以形成分子內輔色(intramolecular copigmentation)和自締合(self-association),保護花色苷的烊鹽離子免受親核攻擊[28]。由于葡萄品種和葡萄生長環境等因素的影響,歐亞種葡萄中花色苷的酰化率分布在0%~53.5%[29-30]。LAGO-VANZELA等[31]發現巴西雜交葡萄品種Violeta含有大量香豆酰化雙糖花色苷(非酰化雙糖花色苷1 146.45 mg/L和對香豆酰化雙糖花色苷408.87 mg/L,酰化率26.24%),在25 ℃和35 ℃下陳釀120 d,表現出較好的顏色穩定性[31]。本研究發現,赤霞珠葡萄酒中花色苷的總酰化率高達36.8%,而黑珍珠葡萄酒只有7.67%,因此花色苷的酰化率較低可能也是導致黑珍珠葡萄酒顏色不穩定的因素之一。
針對非歐亞種葡萄雙糖花色苷含量高,顏色不穩定的問題,主要的改良思路有:a)與歐亞種葡萄雜交,降低5-O-葡萄糖苷酶的表達量,從而減少雜交葡萄中雙糖花色苷的比例[32];b)修飾雙糖花色苷的結構,添加黃烷-3-醇或酚酸,與雙糖花色苷形成輔色[11]和酰化作用[31, 33],在結構上增強其穩定性。然而,采用雜交育種的方法周期長,對葡萄酒花色苷組成的改良作用有限;而采用化學方法修飾,目前僅在模擬體系中進行試驗,尚未應用于葡萄酒體系中,關于提高雙糖苷穩定性的方法仍有待進一步探索。
4 結論
本研究對氧化過程中葡萄酒的顏色和花色苷組成進行分析,結果表明,與赤霞珠葡萄酒相比,黑珍珠葡萄酒的顏色衰退迅速,在較短時間內葡萄酒紅色色調減弱,黃色色調上升,色差值變化速度明顯快于赤霞珠葡萄酒。這可能是因為赤霞珠葡萄酒中花色苷主要以單糖苷取代的形式存在,且36.8%的花色苷發生酰化作用;而黑珍珠葡萄酒中單糖花色苷僅占14.53%,葡萄酒顏色主要由未酰化的雙糖花色苷提供,鑒于未酰化雙糖花色苷易氧化的特點,導致黑珍珠葡萄酒花色苷的降解和顏色的衰退。本研究為葡萄酒中雙糖花色苷易發生褐化反應而降低葡萄酒的陳釀潛力,提供了進一步的試驗依據;同時也為黑珍珠葡萄酒的開發和顏色品質的改良奠定了基礎。
