鄂西南五種野生莢蒾屬植物種群結構與動態特征研究
2023-11-01 04:51:20趙奐敦李春輝艾訓儒薛衛星徐來仙向欽周云
廣西植物 2023年9期
趙奐敦 李春輝 艾訓儒 薛衛星 徐來仙 向欽 周云






摘?要:為探明鄂西南地區野生莢蒾屬(Viburnum)植物的種群數量特征及區域分布狀況,并揭示宜昌莢蒾(V. erosum)、樺葉莢蒾(V. betulifolium)、合軸莢蒾(V. sympodiale)、茶莢蒾(V. setigerum)、莢蒾(V. dilatatum)5種莢蒾屬植物種群的生存現狀及發展趨勢,在鄂西南金子山國有林場、木林子國家級自然保護區及七姊妹山國家級自然保護區,共布設27 hm2固定動態監測樣地對莢蒾屬植物進行調查,通過對5種莢蒾屬植物種群進行年齡結構、動態量化分析和靜態生命表以及相關曲線等研究,探討莢蒾屬植物種群結構與動態特征及未來發展潛力。結果表明:(1)鄂西南莢蒾屬植物在分布區域及種群數量大小上均存在明顯差異。(2)種群結構與動態分析顯示,5種莢蒾屬植物種群年齡結構呈現為金字塔型,幼齡階段種群個體數量較多,具有較強的增長潛力,但對外界干擾均具有較強的敏感性。(3)靜態生命表顯示,5種莢蒾屬植物存活量均隨著齡級的增加而單調遞減;消失率與死亡率曲線變化趨勢相似,但不同植物消失率與死亡率曲線波動具有差異性;種群存活曲線均趨近于Deevey-Ⅱ型。(4)4個生存函數曲線表明,5種莢蒾屬植物種群均表現出前期大量減少、中期趨于穩定、后期衰退的特點。綜上認為,鄂西南莢蒾屬植物資源豐富,對其屬植物的合理開發具有一定的科學性和可行性,幼齡階段時可適當進行人工疏伐,提高保存率;中齡穩定階段時可篩選觀賞性狀優勢種進行適當開發利用;部分物種則需要加強保護,以維持該地區生物多樣性。
關鍵詞: 莢蒾屬, 種群結構, 動態量化分析, 靜態生命表, 生存函數
中圖分類號:Q948
文獻標識碼:A
文章編號:1000-3142(2023)09-1747-12
收稿日期:2022-11-14
基金項目:中國生態環境部生物多樣性調查評估項目(2019HJ2096001006); 湖北省林業科學研究院委托項目(H20064)。
第一作者: 趙奐敦(1996-),碩士研究生,主要從事森林生態系統結構與功能研究,(E-mail)2429989761@qq.com。
*通信作者:李春輝,高級農藝師,主要從事植物生態及生物資源開發利用研究,(E-mail)1106125155@qq.com。
Population structure and dynamic characteristics of five
wild Viburnum plants in Southwest Hubei, China
ZHAO Huandun1, LI Chunhui2*, AI Xunru1, XUE Weixing1,
XU Laixian1, XIANG Qin1, ZHOU Yun1
( 1. School of Forestry and Horticulture, Hubei Minzu University, Enshi 445000, Hubei, China; 2. Agriculture and Rural
Affairs Bureau of Enshi Tujia and Miao Autonomous Prefecture, Enshi 445000, Hubei, China )
Abstract:To explore the population quantitative characteristics and regional distribution of wild Viburnum plants in Southwest Hubei, and reveal the survival status and development trend of the following five Viburnum species: V. erosum, V. betulifolium, V. sympodiale, V. setigerum and V. dilatatum. A total of 27 hm2 dynamic monitoring stands were set up in Jinzi Mountains National Forest Farm, Mulinzi National Nature Reserve and Qizimei Mountains National Nature Reserve in Southwest Hubei. The age structure, dynamic quantitative analysis and static life table of five Viburnum species were used to explore the population structure, dynamic characteristics and future development potential of Viburnum plants. The results were as follows: (1) The distribution area and population size of Viburnum plants in Southwest Hubei were significantly different. (2) The age structure of the five Viburnum species displayed as the pyramidal type summarized from population structure and dynamic analysis. The number of individuals of each population was large at the young stage (89.48% of V. erosum in Jinzi Mountains, 76.38% of V. betulifolium, 67.24% of V. erosum in Mulinzi, 62.11% of V. dilatatum, 56.53% of V. sympodiale, 58.44% of V. setigerum), and only a small proportion of individuals (0.18% of V. erosum in Jinzi Mountains, 0.85% of V. betulifolium, 3.29% of V. erosum in Mulinzi, 3.97% of V. dilatatum, 8.37% of V. sympodiale, 2.79% of V. setigerum) at the old stage. The young stage plants of five Viburnum species all had strong growth potentials but were sensitive to external disturbances. (3) The survival quantity of the five Viburnum species was decreasing monotonically with increasing age class presented by static life tables. Except for V. setigerum, the life expectancy reached peak at the young age, all the other Viburnum species populations reached the peak at the middle age. The change trend of mortality rate and vanish rate were similar, but the fluctuation of vanish rate and mortality rate curves were different among different species. The survival curves were tended to be Deevey-Ⅱ type. (4) The four survival function curves showed that all five Viburnum species populations exhibited a substantial decrease at the early stage, stabilization at the middle stage, and decline at the late stage. To sum up, plant resources of Viburnum species are abundant in Southwest Hubei. Therefore, it is scientific and feasible to exploit Viburnum plants rationally. At the young stage, artificial thinning can be carried out appropriately to improve the preserving rate. During the stable middle stage, the dominant species which have good ornamental characters can be selected for appropriate development and utilization. However, some species need to be protected to maintain biodiversity in this area.
Key words: Viburnum, population structure, dynamic quantitative analysis, static life table, survival function
種群是物種和植物群落的基本單位,對種群的重要屬性即種群結構與動態進行研究也一直是種群生態學研究的主要內容(Condit et al., 2000;Manabe et al., 2000;牛翠娟等,2015;劉浩棟等,2020)。種群結構與動態既能反映種群在特定階段的數量分布和配置情況,又能揭示種群與環境之間的關系、種群動態變化及預測未來發展趨勢,對植物資源的保護和利用具有重要價值(金慧等,2017;何斌等,2020)。種群統計是研究種群結構與動態的基本方法,主要包括種群年齡結構、動態量化分析、靜態生命表、存活曲線、生存函數分析等核心內容(王卓等,2009)。其中,種群年齡結構及動態量化分析不僅可以揭示種群在各齡級階段的現存數量、種群的更新繁殖能力及種群未來發展的穩定性,還能在一定程度上反映種群在所處環境中的適應能力及種群在該群落中的地位和作用(劉普幸,2011;拓鋒等,2021)。種群動態量化分析是對種群結構動態分析中等級歸類劃分粗放等缺點進行彌補(陳曉德,1998)。靜態生命表可以直觀地展現種群的存活率、死亡率及生命期望等重要種群特征,是研究種群結構與動態變化的主要手段,其與生存函數結合對種群進行分析,對揭示種群的生命過程、生存現狀及種群在一定條件下生存、繁衍及變化的可能性具有重要意義(羅西等,2021;拓鋒等,2021)。因此,綜合利用各種群結構與動態的研究方法,更有助于人們深入了解種群的現狀、動態特征及演替規律,預測種群未來發展趨勢,對種群的保護與利用具有重要指導意義(解婷婷等,2014)。
莢蒾屬(Viburnum)植物隸屬于五福花科(Adoxaceae),全球約200種,是亞熱帶森林生態系統重要的組成部分,也是極具開發價值的觀賞灌木屬,在國際上被譽為“萬能綠化灌木”。湖北省分布有莢蒾屬植物43種,僅鄂西南恩施州就有28種(含22種6變種)(艾訓儒等,2018)。因為該區域最具代表性的植被類型為亞熱帶常綠落葉闊葉混交林,所以目前有關群落中植物種群結構與動態的研究主要集中在優勢種(姚蘭等,2017;朱強等,2019;王進等,2020)、瀕危植物(楊心兵等,2000;林勇等,2017;羅西等,2021)等相關研究,而極少涉及在群落中較為劣勢卻具有較高觀賞價值的莢蒾屬植物。據報道,國內約90%的莢蒾屬植物資源仍處于野生狀態(呂文君等,2019),并且對該屬的研究主要集中在常見種的種子休眠與萌發特性(肖月娥等,2007;楊軼華等,2015)、扦插繁殖(黃增艷和宋蕾,2007)、化學成分及元素含量測定(白長財等,2011;孫曉蕾等,2020)等方面,因此查清莢蒾屬植物資源的野生分布現狀,探究優勢莢蒾屬植物的生存現狀與更新潛力,這對莢蒾屬植物原生種種質資源的調查、保護及開發栽培利用等具有重要意義。
本研究以鄂西南金子山國有林場、木林子國家級自然保護區及七姊妹山國家級自然保護區內的莢蒾屬植物種群為研究對象,基于莢蒾屬植物資源野外調查數據,篩選出5個種群個體數量較為優勢的莢蒾屬植物為代表進行種群結構與動態特征分析。通過對5種優勢莢蒾屬植物的種群個體數量特征、分布現狀、種群結構與動態特征、野外生存現狀、自然更新與發展趨勢等進行分析討論,以期為國內莢蒾屬植物資源的開發利用等提供一定理論基礎,同時為鄂西南地區生物多樣性的保護與維持提供一定科學依據。
1?研究區概況
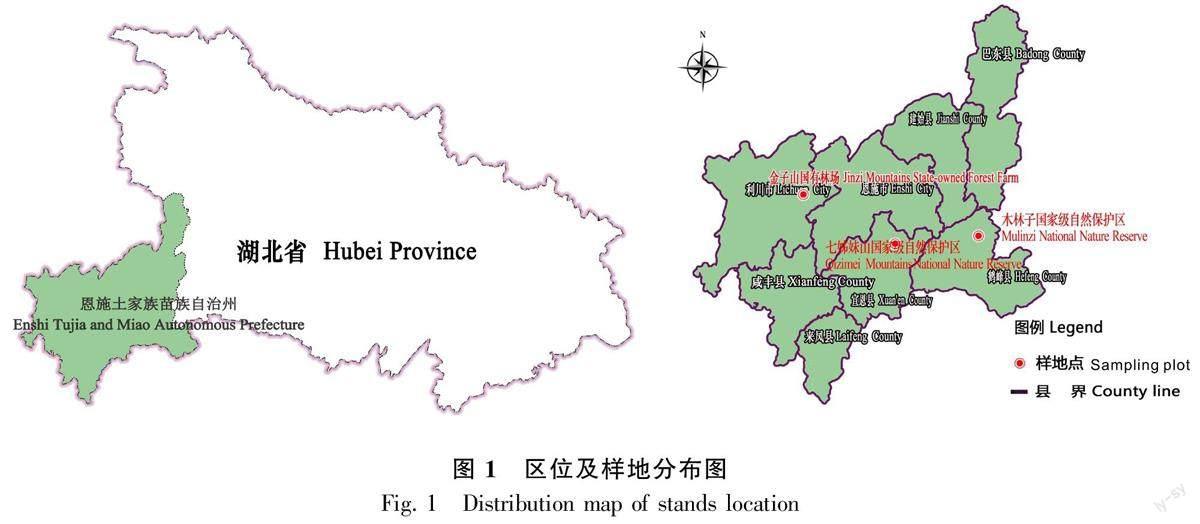
鄂西南地處云貴高原與東部低山丘陵的過渡區域,與重慶、湖南相鄰,主要組成部分是恩施土家族苗族自治州,地勢起伏大,地形復雜多樣,主要為喀斯特地貌,境內小氣候豐富,主要為亞熱帶大陸性季風濕潤氣候,具有豐富的野生動植物資源。研究樣地位于鄂西南金子山國有林場、木林子國家級自然保護區和七姊妹山國家級自然保護區的常綠落葉闊葉混交林內,總面積為27 hm2,土壤類型主要為棕壤或黃棕壤,年均氣溫為12.7 ~ 15.5 ℃,年均降水量為1 475.0 ~ 1 667.5 mm,年日照時數為1 212~1 300 h,無霜期約為4個月(王進等,2020);樣地中木植被有明顯垂直分層,群落物種組成多樣(姚蘭等,2017;朱強等,2019)(表1,圖1)。
2?研究方法
2.1 樣地建設及野外調查
2014年4月至2016年9月,通過實地調查,在金子山國有林場、木林子國家級自然保護區、七姊妹山國家級自然保護區中選擇植被保存較為完整的天然林,利用全站儀按照CTFS(Center for Tropical Forest Science)樣地建設標準(Condit, 1998)布設固定監測樣地,面積共27 hm2,包含675個20 m × 20 m的固定樣地,其中木林子大樣地為15 hm2,包含375個20 m × 20 m的固定樣地;金子山及七姊妹山大樣地均為6 hm2,分別包含150個20 m × 20 m的固定樣地(王進等,2020)。野外調查以20 m × 20 m的固定樣地為基本單元,用紅色油漆對樣地中所有胸徑(DBH)≥ 1 cm的木本植物個體在距離地面1.3 m處的樹干上進行標記,并掛上特制的具有唯一編號的鋁牌;進行物種學名(科、屬、種)鑒定,測定并記錄符合要求的木本植物個體的胸徑(DBH)、冠幅、樹高(H)及生長狀況等信息;根據設定的坐標原點,記錄樣地中所有木本植物個體的相對坐標值(X,Y)。基于野外調查數據,對所有樣地中的莢蒾屬植物進行統計,并分析各莢蒾屬植物種群個體數量及在各樣地中的分布情況。結合樣地及各莢蒾屬植物種群個體數量等實際情況,挑選5個種群個體數量較為優勢的莢蒾屬植物進行種群結構與動態特征分析。
2.2 種群結構劃分
采用徑級代替齡級的方法,結合樣地內5種莢蒾屬植物的徑級特征,以5 mm為步長,按照莢蒾屬植物胸徑將大小級劃分為9個徑級:Ⅰ級,1 cm≤ DBH<1.5 cm;Ⅱ級,1.5 cm≤DBH<2 cm,Ⅲ級,2 cm≤DBH<2.5 cm;Ⅳ級,2.5 cm≤DBH<3 cm;Ⅴ級,3 cm≤DBH<3.5 cm;Ⅵ級,3.5 cm≤DBH<4 cm;Ⅶ級,4 cm≤DBH<4.5 cm;Ⅷ級,4.5 cm≤DBH<5 cm;Ⅸ級,DBH≥5 cm。根據徑級將所有個體劃分為幼齡(Ⅰ級,Ⅱ級)、低齡(Ⅲ級,Ⅳ級)、中齡(Ⅴ級,Ⅵ級)和高齡(Ⅶ級以上)4個階段。采用陳曉德(1998)的種群結構動態量化分析方法,對5種莢蒾屬植物種群結構進行定量分析。其中,Vpi和V′pi分別為不考慮和考慮外部干擾因素下整個種群結構的數量變化動態指數,P極大(Pmax)為種群隨機干擾風險極大值。Vn、Vpi、V′pi取正、0、負值時分別反映種群個體數量的增長、穩定、衰退的結構動態關系。
2.3 種群靜態生命表及存活曲線
靜態生命表及存活曲線在研究種群的結構與動態特征中具有重要作用,能直觀地展現種群在不同徑級的實際生存個體數、死亡數量及存活趨勢,由于所研究的莢蒾屬植物均為野生種群,為避免生命表中出現死亡率為負的情況,因此采用勻滑技術對數據進行處理(江洪,1992;吳承禎等,2000)。將5種莢蒾屬植物種群全部個體按照大小級編制靜態生命表,并以大小級為橫坐標,繪制5種莢蒾屬植物的死亡率曲線、消失率曲線及存活曲線;采用Hett和Louckst(1976)提出的指數函數模型Nx=N0e-bx(代表Deevey-Ⅱ型曲線)與冪函數模型Nx=N0x-b (代表Deevey-Ⅲ型曲線) 對種群存活曲線進行檢驗。引入生存函數S(i)、i表示大小級、累積死亡率函數F(i)、死亡密度函數f(ti)和危險率函數λ(ti)ti表示種群生存時間4個生存分析函數對5種莢蒾屬植物種群結構進行分析,hi為區間長度(李金昕等,2016)。
2.4 數據處理
采用Excel對文中的數據進行處理及分析,使用Origin 2021軟件繪制死亡率與消失率曲線、存活曲線及生存函數曲線,并用SPSS軟件對各種群大小級進行Kolomogorov-Smirnov檢驗(簡稱KS檢驗)。
3?結果與分析
3.1 莢蒾屬植物種群數量特征及分布現狀
據調查所得(表2),鄂西南3個區域莢蒾屬植物共13種,分別為巴東莢蒾(Viburnum henryi)、茶莢蒾(V. setigerum)、樺葉莢蒾(V. betulifolium)、莢蒾(V. dilatatum)、宜昌莢蒾(V. erosum)、水紅木(V. cylindricum)、合軸莢蒾(V. sympodiale)、球核莢蒾(V. propinquum)、短序莢蒾(V. brachybotryum)、粉團(V. plicatum)、蝴蝶戲珠花(V. plicatum)、少花莢蒾(V. oliganthum)和陜西莢蒾(V. schensianum),各莢蒾屬植物的分布狀況及種群個體數量存在顯著差異。其中,合軸莢蒾分布在金子山、木林子樣地,球核莢蒾分布在金子山、七姊妹山樣地,少花莢蒾和陜西莢蒾僅分布在金子山樣地,短序莢蒾和粉團僅分布在木林子樣地,蝴蝶戲珠花僅分布在七姊妹山樣地,其余莢蒾屬植物則在3個調查樣地均有分布。樣地中莢蒾屬植物共有8 198株,個體總數為金子山 > 木林子 > 七姊妹山,其中種群個體數量較為優勢的有樺葉莢蒾、茶莢蒾、莢蒾、宜昌莢蒾、合軸莢蒾等,其余莢蒾屬植物種群個體數量均比較少,因此本研究以這5種優勢種為主要研究對象。
3.2 5種莢蒾屬植物種群結構特征及動態變化分析
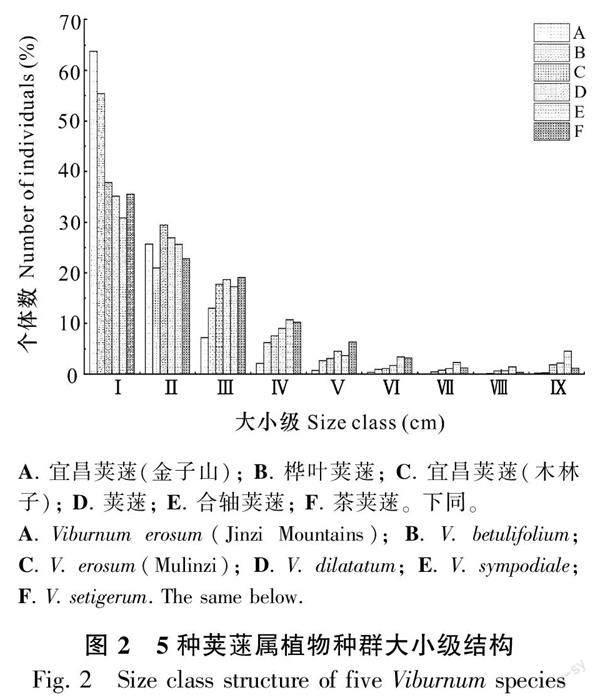
對3個區域中各莢蒾屬植物種群大小級個體數量進行分析,結果見圖2。由圖2可知,5種莢蒾屬植物種群個體數量均隨大小級的增加而總體呈現下降趨勢。5種莢蒾屬植物種群在幼齡階段的個體數量占比均較大,但相較于其他區域中莢蒾屬植物種群各大小級個體數量分布情況,金子山宜昌莢蒾、樺葉莢蒾種群在幼齡階段個體數量變化更明顯,說明在該樣地中宜昌莢蒾和樺葉莢蒾種群在幼齡階段對環境的適應能力較弱。除了木林子樣地中合軸莢蒾種群個體數量在高齡階段占比大于中齡階段且種群大小級結構呈現為不規則的金字塔型以外,其他莢蒾屬植物種群在高齡階段的個體數量占比均最小且種群大小級結構均呈現為高度規則的金字塔型。由此可知,5種莢蒾屬植物種群均具有大量的幼齡樹,種群的增長狀態穩定,表明鄂西南地區5種莢蒾屬植物種群具有長期穩定的自我發展能力及維持持續繁衍更新的能力。對各莢蒾屬植物種群的大小級進行精確性KS檢驗得到,宜昌莢蒾(金子山)、樺葉莢蒾、宜昌莢蒾(木林子)、莢蒾、合軸莢蒾及茶莢蒾的KS值分別為0.160、0.429、0.466、0.537、0.405和0.748,不同種群呈現為正態分布,顯著性值均大于0.05。
對3個區域中各莢蒾屬植物進行動態量化分析,結果見表3。由表3可知,5種莢蒾屬植物種群均在Ⅸ級出現負值的情況,呈現衰退狀態,但5種莢蒾屬植物種群數量動態變化指數Vpi(不考慮外部干擾)和V′pi(考慮外部干擾)均顯示Vpi>V′pi>0,說明5種莢蒾屬植物種群總體屬于增長型,這與種群大小級結構分析結果一致。木林子的莢蒾、合軸莢蒾種群及七姊妹山的茶莢蒾種群V′pi指數均不大于1,說明在考慮外界干擾情況下,這3個莢蒾屬植物種群的動態變化趨勢處于增長型與穩定型之間,各種群動態變化指數值由大到小為茶莢蒾、莢蒾、合軸莢蒾種群;金子山宜昌莢蒾、樺葉莢蒾種群及木林子宜昌莢蒾種群V′pi指數均大于1。種群結構對隨機干擾的敏感性指數(Pmax)均大于0,說明5種莢蒾屬植物種群穩定性受到外界環境干擾的敏感性強,并且敏感性最高的是金子山宜昌莢蒾,最低的是木林子合軸莢蒾。
3.3 5種莢蒾植物種群靜態生命表及存活曲線
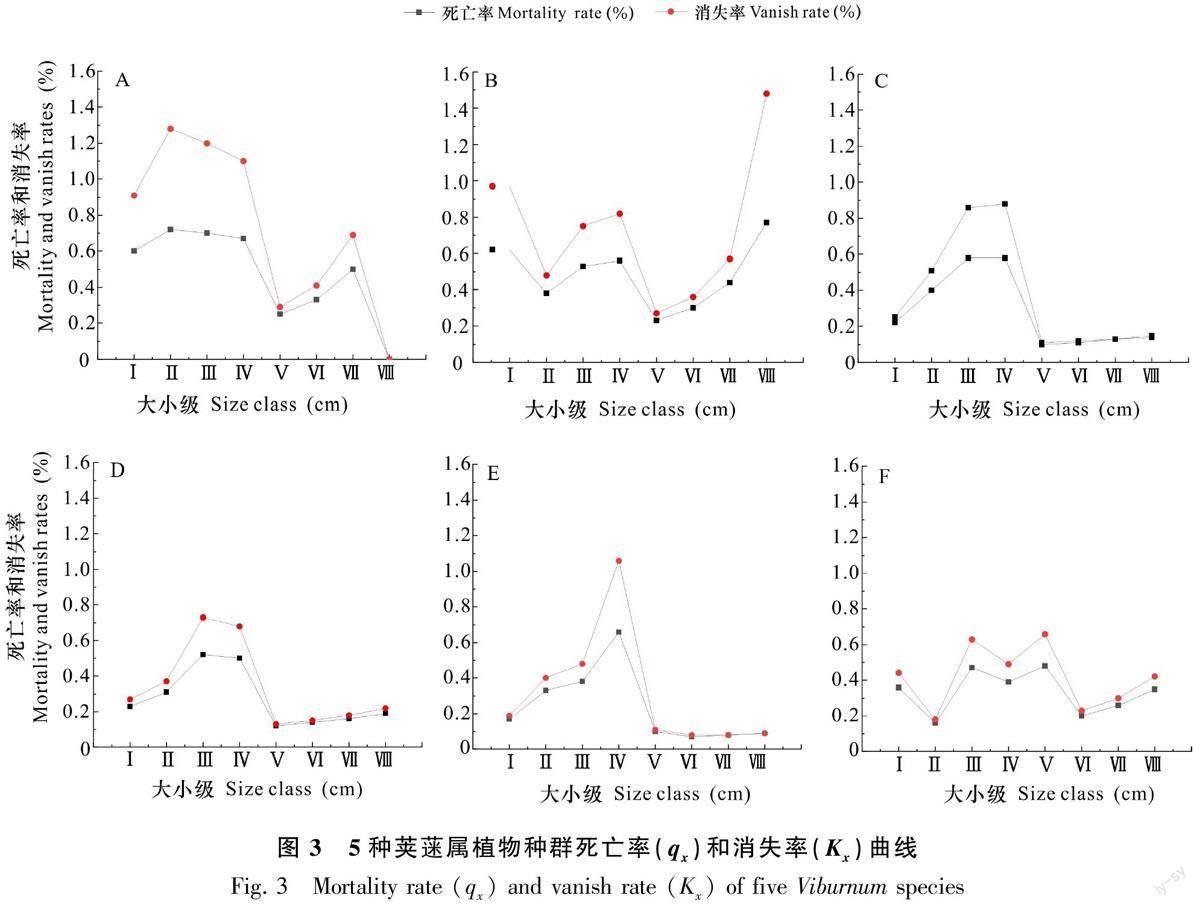
對3個區域中5種莢蒾屬植物種群的數據進一步整理并計算得到種群靜態生命表,結果見表4。由表4可知,隨著大小級的增加,5種莢蒾屬植物種群的標準化存活個數(lx)逐漸減小,符合物種的生物學特性。種群個體的期望壽命(ex)除七姊妹山茶莢蒾種群外均在Ⅴ級處出現最高峰值, 其中合軸莢蒾種群的期望壽命值最大,且5種莢蒾屬植物種群的期望壽命值均在Ⅷ級時偏低,說明在Ⅷ級時各種群已處于生理衰退期。5種莢蒾屬植物種群消失率(Kx)與死亡率(qx)曲線變化趨勢一致(圖3),但不同莢蒾屬植物種群的死亡率與存活率曲線波動具有較大差異。金子山宜昌莢蒾、樺葉莢蒾種群的死亡率與存活率分別在Ⅱ級、Ⅰ級時段達到第一次峰值,其中,宜昌莢蒾種群死亡率與消失率在Ⅰ~Ⅴ級呈現先上升后下降的趨勢,而樺葉莢蒾種群死亡率與消失率在Ⅰ~Ⅴ級呈現先降后升的趨勢,但2種莢蒾屬植物種群的期望壽命值總體均呈現上升的趨勢。木林子3種莢蒾屬植物的死亡率與存活率則在Ⅰ級和Ⅱ級達到第一次峰值,3種莢蒾屬植物種群死亡率與消失率在Ⅰ~Ⅴ級呈現先升后降的趨勢,期望壽命值則先減后增;在Ⅴ級后,3種莢蒾屬植物種群死亡率與消失率變化較為緩慢,但期望壽命值下降明顯。七姊妹山樣地中茶莢蒾種群死亡率與消失率在Ⅰ~Ⅶ級的變化較為平緩,期望壽命值在Ⅱ級時達到峰值,之后整體呈下降趨勢。綜上認為,5種莢蒾屬植物在幼齡、低齡階段種群易受到競爭、環境篩選等作用的影響,但在中齡階段后,除茶莢蒾以外,其他莢蒾屬植物受到的環境篩選和競爭壓力相對減弱,環境適應能力增強,并且合軸莢蒾、莢蒾在樣地中有更好的生存能力。
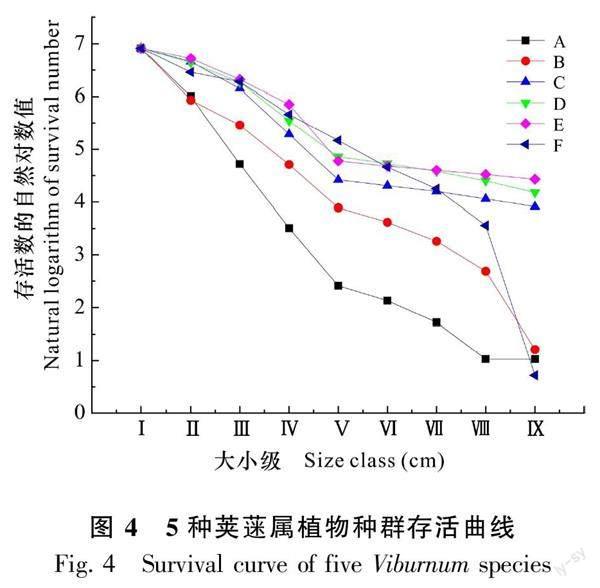
不同樣地的莢蒾屬植物種群的存活曲線趨勢變化不一致,結果見圖4。由圖4可知,其中木林子3種莢蒾屬植物種群及金子山宜昌莢蒾種群的變化趨勢整體呈現為“凸-凹-凸”型,但七姊妹山茶莢蒾種群則在整體呈現出“凹-凸-凹”的變化趨勢,而金子山樺葉莢蒾種群則呈現為“凹-凸-凹-凸-凹”的變化趨勢。總體而言,5種莢蒾屬植物種群的存活曲線介于“Deevey-Ⅱ”和“Deevey-Ⅲ”型之間。結合Helt和Louckst提出的指數函數和對數函數,對5種莢蒾屬植物種群存活曲線進行檢驗(表5),對比擬合的兩種模型的R2,各莢蒾屬植物種群的指數模型R2值均大于冪函數模型,且P值均小于0.001,結合靜態生命表和存活曲線分析認為,可以明顯得出5種莢蒾屬植物種群存活曲線與“Deevey-Ⅱ”型相近,種群存活曲線的變化趨勢和模型檢驗的結果保持一致,說明各莢蒾屬植物種群結構總體處于穩定狀態,但不同種群存在一定差異性。
3.4 5種莢蒾屬植物種群生存分析
基于種群靜態生命表,對5種莢蒾屬植物種群的生存分析結果(圖5)顯示,各莢蒾屬植物種群累積死亡率隨大小級單調遞增,生存率隨大小級單調遞減,并且均表現為幼齡、低齡階段變化幅度大,在中齡及高齡階段變化幅度小,說明天然條件下5種莢蒾屬植物幼齡階段死亡率都較高,這與靜態生命表的分析結果一致。木林子3種莢蒾屬植物種群及七姊妹山樣地茶莢蒾種群的生存率與累積死亡率均在Ⅱ級時達到平衡,而金子山宜昌莢蒾、樺葉莢蒾種群的累積死亡率則一直高于生存率,這說明金子山宜昌莢蒾、樺葉莢蒾種群在幼齡階段之前已經進入衰退,木林子及七姊妹山莢蒾屬植物種群進入衰退期的時間則在幼齡階段時期。5種莢蒾屬植物的死亡密度曲線均隨著大小級的增加而呈現波動下降的趨勢且在幼齡及低齡階段的值相對較高,說明5種莢蒾屬植物種群個體數量在幼齡及低齡階段大量銳減。5種莢蒾屬植物種群的危險率在Ⅰ~Ⅴ級隨大小的增長而單調遞增,進一步說明5種莢蒾屬植物種群個體數量在幼齡及低齡階段隨著齡級的增加而減少,之后危險率變化趨勢則隨著大小級的增加而逐漸趨于平緩。4個生存函數曲線表明,5種莢蒾屬植物種群均表現為前期個體數大量減少,中期趨于穩定,后期衰退的狀態。
4?討論
4.1 樣地中莢蒾屬植物種群數量特征及分布現狀
種群數量特征可以表征種群個體數量及分配狀況,對于揭示群落的形成和維持機制具有重要意義(費世民等,2008)。對鄂西南3個區域莢蒾屬植物個體數量對比分析得到,單位面積內莢蒾屬植物個體數量排序依次為金子山 > 七姊妹山 > 木林子。3個區域中莢蒾屬植物均為灌木或小喬木,在群落中基本處于中下層,各區域地理位置及氣候相近,受水熱條件差異的影響較小,但群落演替階段存在差異性,群落演替程度為木林子 > 七姊妹山 > 金子山(艾訓儒等,2018;王進等,2020;李瑋宜等,2021),這說明各區域莢蒾屬植物個體數量受到群落演替階段的影響。從鄂西南各莢蒾屬植物種群個體數量來看,單位面積內種群個體數量大小為樺葉莢蒾 > 茶莢蒾 > 莢蒾 > 宜昌莢蒾 > 合軸莢蒾等。由此可見,不同莢蒾屬植物在同一地區的種群大小存在明顯差異,這可能與不同種群對環境適應能力、繁殖及生存能力的差異性有關。對5種莢蒾屬植物優勢種群的分布情況進行分析,樺葉莢蒾主要分布于金子山,茶莢蒾主要分布于七姊妹山,莢蒾與合軸莢蒾主要分布在木林子,宜昌莢蒾則在金子山與木林子均有分布。一般而言,同屬物種具有相似的進化特征和生態適應性,導致其對環境資源的利用也可能具有相似性,因此它們可能會面臨更強烈的種間競爭,進而影響共存(Ackerly & Donoghue,1998)。綜上所述,鄂西南野生莢蒾屬植物資源豐富,但各莢蒾屬植物種群個體數量及分布情況均存在明顯的差異性。因此,對于種群較為優勢的物種,可考慮進行開發利用,并加強對其自身生物學性狀、競爭能力及環境影響因素等的進一步研究;而較為劣勢或稀少的種群則需加強其野生種質資源的保護。
4.2 5種莢蒾屬植物種群結構特征
種群的徑級結構可以替代種群年齡結構來反映種群發展對生境的適應與種群的生存狀態(陳育等,2014)。本研究對3個區域中5種莢蒾屬植物的種群結構與動態特征分析顯示,各莢蒾屬植物種群的個體數量均隨著大小級的增大而逐漸減小,總體均呈現為金字塔結構,這與李功麟等(2021)對固沙灌木種群年齡結構的研究結果相似,種群幼齡階段數量多,可以適應當地生境,通過大量繁殖幼苗的策略來維持種群穩定性,但高齡階段種群個體數量相對較少,反映各莢蒾屬植物種群在高齡階段的植株生存較為困難的現象。對各種群的大小級進行KS檢驗,得出各種群均呈現正態分布。種群中個體對環境的適應能力可以通過種群數量動態來反映(Bierzychudek, 2014)。本研究對各莢蒾屬植物種群動態量化分析顯示,5種莢蒾屬植物都對外部環境具有較高的敏感性,表現為種群整體均呈現為穩定增長、后期衰退的趨勢。結合樣地中實際調查情況,莢蒾屬植物在森林群落中主要位于中下層的位置,在群落中屬于相對劣勢的物種,光照等環境因素也在一定程度上影響其種群增長的穩定性,同層種間資源競爭壓力大,并且隨著幼齡個體的逐步增長,需要從環境中獲得更多的資源,因此種內競爭也逐漸加劇。在不考慮和考慮外部干擾時,各莢蒾屬植物種群增長潛力均為宜昌莢蒾(金子山) > 樺葉莢蒾 > 宜昌莢蒾(木林子)> 茶莢蒾 > 莢蒾 > 合軸莢蒾,但在考慮外部干擾時,宜昌莢蒾及樺葉莢蒾種群具有更大的增長潛力;其他莢蒾屬植物的動態變化為處于增長型和穩定型之間的過渡階段,這進一步反映各莢蒾屬植物種群增長及動態變化趨勢的差異性可能是受到其生境及植物自身性狀的影響。
4.3 5種莢蒾屬植物種群動態變化趨勢
靜態生命表、存活曲線、生存函數分析等不僅能反映種群的生存狀況及環境對其的影響程度,還可以預測種群未來的發展趨勢(陳曉德,1998)。本研究對3個區域內莢蒾屬植物種群數量進行統計,并編制種群靜態生命表以及相關曲線等來分析種群的生存狀態及發展趨勢。5種莢蒾屬植物種群存活曲線均趨近于Deevey-Ⅱ曲線,說明各莢蒾屬植物種群具有相對穩定的發展趨勢,存活量隨大小級的增加而減少。七姊妹山茶莢蒾種群的期望壽命值在Ⅱ級時出現最高峰,其他莢蒾屬植物種群的期望壽命值均在Ⅴ級處出現最高峰且合軸莢蒾種群期望壽命值最大,說明七姊妹山茶莢蒾種群在幼齡階段時能夠更好地適應生存環境;而其他莢蒾屬植物則在中齡階段時對環境的適應能力較好,其中合軸莢蒾的適應能力最強,但隨著大小級的增加,各莢蒾屬植物種群在高齡階段均處于生理衰退期。這可能與七姊妹山樣地處于干擾后的中期階段且樣地中主要物種的大徑級個體在空間分布上表現出特定的生境偏好有關(姚良錦等,2017)。
5種莢蒾屬植物種群的死亡率與存活率曲線研究結果顯示,各莢蒾屬植物種群的死亡率與存活率曲線變化趨勢較為接近,但不同莢蒾屬植物種群的死亡率與存活率曲線波動差異較為明顯,這可能與環境的篩選作用及各莢蒾屬植物的資源競爭力有關。各莢蒾屬植物種群的死亡率與存活率均在幼齡或低齡階段達到第一次峰值,由于各莢蒾屬植物種群在幼齡和低齡個體密度大,隨著生長過程中對資源和空間的需求增加,種內及種間競爭能力增強,因此由密度制約效應引發的種群自疏和他疏作用導致種群有較高的死亡率(韓文軒和方精云,2008;何斌等,2020)。
生存函數能夠更直觀、具體地說明種群的結構和動態變化(李文英等,2018)。本研究生存函數分析結果與靜態生命表分析結果一致,5種莢蒾屬植物種群均表現為前期幼齡及低齡階段個體數大量減少,中期趨于穩定,后期衰退的狀態,這說明在天然更新下,雖然各莢蒾屬植物種群在幼齡階段死亡率較高,種群個體數量急劇下降,但大量的幼齡及低齡樹依舊對種群中齡階段具有較強的貢獻,表明通過環境篩選及密度制約而存活的樹種對環境具有較強的適應及資源競爭能力,有助于維持種群的平穩發展,而在高齡階段,各莢蒾屬植物則進入生理衰退時期(李文英等,2018)。
目前,國內關于野生莢蒾屬植物野外資源的研究較少,對莢蒾屬植物的認知嚴重不足,雖然本研究掌握了鄂西南部分地區莢蒾屬植物的基礎數據,并揭示了部分較為優勢的莢蒾屬植物種群結構與動態特征,但各莢蒾屬植物的功能性狀、繁殖更新能力、種內及種間關系等還有待進一步研究。因此,建議查明該地區其他莢蒾屬植物種群較為弱勢的原因及其種群結構與動態變化,并加強對該地區各莢蒾屬植物的監測與系統性研究,明確各莢蒾屬植物自身生物學特性及空間分布特征,為維護鄂西南地區的生物多樣性提供幫助,也為國內其他地區的莢蒾屬植物資源的研究及開發利用提供科學依據。
5?結論
鄂西南地區是中國莢蒾屬植物分布的主要區域,莢蒾屬植物受到物種自身性狀、種間關系及群落演替階段的影響,在分布情況及種群個體數量方面都具有明顯差異。研究區內個體數量較多的5種莢蒾屬植物為樺葉莢蒾、宜昌莢蒾、莢蒾、茶莢蒾和合軸莢蒾;5種莢蒾屬植物種群大小級結構呈現為金字塔型,種群增長穩定,幼齡個體數量多,自我更新能力較強,具有較大的開發潛力,但種群又易受到外界環境干擾、物種的更新能力及資源競爭力等因素的影響,尤其是各種群在高齡階段時均受到較強的影響。因此,考慮該屬植物對環境資源需求的前提下,在對鄂西南莢蒾屬植物資源保護利用過程中,幼齡階段時可適當進行人工疏伐,提高保存率;中齡穩定階段時可篩選性狀優勢種進行適當開發利用。
參考文獻:
AI XR, HUANG S, YAO L, et al., 2018. Flora of Enshi [M]. Beijing: Science Press, 4: 103-116.[艾訓儒, 黃升, 姚蘭, 等, 2018. 《恩施植物志》 [M]. 北京: 科學出版社, 4: 103-116.]
ACKERLY DD, DONOGHUE MJ, et al., 1998. Leaf size, sapling allometry, and Corners Rules: phylogeny and correlated evolution in maples [J]. Am Nat, 152(6): 767.
BAI CC, HAN L, ZHAO YS, 2011. Determination of 12 inorganic elements in Viburnum betulifolium of fruit [J]. Asia-Pacific Trad Med, 7(10): 18-19.[白長財, 韓璐, 趙云生, 2011. 樺葉莢蒾果實中12種無機元素含量測定 [J]. 亞太傳統醫藥, 7(10): 18-19.]
BIERZYCHUDEK P, 2014. Plant biodiversity and population dynamics [M]//MONSON RK. Ecology and the Environment. New York: Springer, 8: 29-65.
CONDIT R, ASHTON PS, BAKER P, et al., 2000. Spatial patterns in the distribution of tropical tree species [J]. Science, 288(5470): 1414-1418.
CONDIT R, 1998.Tropical forest census plots: methods and results from Barro Colorado Island, Panama and a comparison with other plots[M]. Berlin: Springer: 23-55.
CHEN XD, 1998. A study on the method of quantitative analysis for population and community structural dynamics [J]. Acta Ecol Sin, 18(2): 104-107.[陳曉德, 1998. 植物種群與群落結構動態量化分析方法研究 [J]. 生態學報, 18(2): 104-107.]
CHEN Y, YANG J, ZHANG PJ, et al., 2014. Population structure and spatial point pattern of Helianthemum soongoricum in West Ordos, Inner Mongolia, China [J]. J Desert Res, 34(1): 75-82.[陳育, 楊劼, 張璞進, 等,2014. 西鄂爾多斯半日花(Helianthemum soongoricum)種群結構和點格局分析 [J]. 中國沙漠, 34(1): 75-82.]
FEI SM, HE YP, CHEN XM, et al., 2008. Quantitative features of populations of Pinus tabulaeformis and P. armandii regenerated following water damage at Qinling Mountain, China [J]. Chin J Plant Ecol, 32(1): 95-105.[費世民, 何亞平, 陳秀明, 等, 2008. 秦嶺水災跡地油松和華山松更新種群數量特征 [J]. 植物生態學報, 32(1): 95-105.]
HE B, LI Q, CHEN QL, et al., 2020. Quantitative characteristics and population dynamics of Pinus massoniana plantation in northwest Guizhou Province[J]. J Centr S Univ For Technol, 40(11): 129-137.[何斌, 李青, 陳群利, 等, 2020. 貴州省西北部馬尾松人工林種群數量特征與動態 [J]. 中南林業科技大學學報, 40(11): 129-137.]
HETT JM, LOUCKS OL, 1976. Age structure models of balsam fir and eastern hemlock [J]. J Ecol, 3(64): 1029-1044.
HAN WX, FANG JY, 2008. Self-thinning law in plant population : -3/2 vs -4/3? [J]. J Peiking Univ(Nat Sci Ed), 44(4): 661-668.[韓文軒, 方精云, 2008. 植物種群的自然稀疏規律—— -3/2還是-4/3? [J]. 北京大學學報(自然科學版), 44(4): 661-668.]
JIN H, ZHAO Y, YIN H, et al., 2017. Quautitative characteristics and dynamic analysis of the endangered species Rhododendron chrysamium population in Changbai Mountain [J]. Chin J Ecol, 36(10): 3123-3130.[金慧, 趙瑩, 尹航, 等, 2017. 長白山瀕危植物牛皮杜鵑(Rhododendron chrysanthum)種群數量特征與動態分析 [J]. 生態學雜志, 36(11): 3123-3130.]
JIANG H,1992. Picea asperata population ecology [M]. Beijing: China Foresty Press: 11-12.[江洪, 1992. 云杉種群生態學 [M]. 北京: 中國林業出版社: 11-12.]
LIU HD, CHEN Q, XU ZY, et al., 2020. Natural population structure and spatial distribution pattern of rare and endangered species of Dacrydium pectinatum [J]. Acta Ecol Sin, 40(9): 2985-2995.[劉浩棟, 陳巧, 徐志揚, 等, 2020. 珍稀瀕危樹種陸均松天然種群結構與空間分布格局 [J]. 生態學報, 40(9): 2985-2995.]
LIU PX, 2011. Study on population structure and dynamics of Populus euphratica in the middle and lower reaches of the Shule River Basin Oasis, Hexi Corridor [J]. J Nat Resour, 26(3): 429-439.[劉普幸, 2011. 疏勒河中下游綠洲胡楊種群結構與動態研究 [J]. 自然資源學報, 26(3): 429-439.]
LUO X, GUO QJ, YAO L, et al., 2021. Characteristics of natural population structure of endangered plant Liriodendron chinense [J]. J Centr S Univ For Technol, 41(7): 115-123.[羅西, 郭秋菊, 姚蘭, 等, 2021. 瀕危植物鵝掌楸的天然種群結構特征 [J]. 中南林業科技大學學報, 41(7): 115-123.]
LIN Y, AI XR, YAO L, et al., 2017. Population structure and dynamics of Metasequoia glyptostroboides parent trees [J]. Chin J Ecol, 36(6): 1531-1538.[林勇, 艾訓儒, 姚蘭, 等, 2017. 水杉原生母樹種群結構與動態 [J]. 生態學雜志, 36(6): 1531-1538.]
L WJ, LIU HT, XIA BS, et al., 2019. Viburnum resources and their application to landscaping [J]. World For Res, 32(3): 36-41.[呂文君, 劉宏濤, 夏伯順, 等, 2019. 莢蒾屬植物資源及其園林應用 [J]. 世界林業研究, 32(3): 36-41.]
LI JX, WU DJ, ZHANG SP, et al., 2016. Life table and dynamic analysis of Fagus hayatae population in Micangshan Nature Reserve, Sichuan Province, China [J]. Bull Bot Res, 36(1): 68-74.[李金昕, 吳定軍, 章世鵬, 等, 2016. 四川米倉山自然保護區臺灣水青岡種群生命表及動態分析 [J]. 植物研究, 36(1): 68-74.]
LI GL, ZHANG DH, ZHANG ZS, et al., 2021. Population dynamics of main sand-fixing shrubs in Gurbantunggut Desert [J]. J Desert Res, 41(2): 129-137.[李功麟, 張定海, 張志山, 等, 2021. 古爾班通古特沙漠沙丘主要灌木的種群數量動態 [J]. 中國沙漠, 41(2): 129-137.]
LI WY, LI X, GAN XH, 2018. Population structure and dynamics of endangered plant Tetracentron sinense [J]. Subtrop Plant Sci, 47(3): 222-228.[李文英, 李欣, 甘小洪, 2018. 瀕危植物水青樹的種群結構與數量動態 [J]. 亞熱帶植物科學, 47(3): 222-228.]
LI WY, AI XR, YAO L, et al., 2021. Biodiversity evaluation of three national nature reserves in Southwest Hubei [J]. Hubei Fore Sci Technol, 50(2):18-22. [李瑋宜, 艾訓儒, 姚蘭, 等, 2021. 鄂西南3個國家級自然保護區生物多樣性評價 [J].湖北林業科技, 50(2):18-22.]
MANABE T, NISHIMURA N, MIURA M, et al., 2000. Population structure and spatial patterns for trees in a temperate old-growth evergreen broad-leaved forest in Japan [J]. Plant Ecol, 151(2): 181-197.
NIU CJ, LOU AR, SUN RY,et al., 2015. Foundations in ecology [M]. Beijing: Higher Education Press: 64-65.[牛翠娟, 婁安如, 孫儒泳, 等, 2015. 基礎生態學(第3版) [M]. 北京: 高等教育出版社: 64-65.]
SUN XL, ZHANG SY, CHEN WH, et al., 2020. Chemical constituents research progress of the genus Viburnum [J]. Chin J Mod Appl Pharm, 37(15): 1907-1913.[孫曉蕾, 張世堯, 陳文華, 等, 2020. 莢蒾屬植物化學成分的研究進展 [J]. 中國現代應用藥學, 37(15): 1907-1913.]
HUANG ZY, SONG L, 2007. The softwood-cutting propagation of Viburnum × carlcehalum [J]. Hubei Agris Sci, 46(6): 950-952.[黃增艷, 宋蕾, 2007. 紅蕾雪球莢蒾嫩枝扦插的繁殖技術 [J]. 湖北農業科學, 46(6): 950-952.]
TA F, LIU XD, HUANG DL, et al., 2021. Quantitative dynamics of Picea crassifolia population in Dayekou basin of Qilian Mountains [J]. Acta Ecol Sin, 41(17): 6871-6882.[拓鋒, 劉賢德, 黃冬柳, 等, 2021. 祁連山大野口流域青海云杉種群數量動態 [J]. 生態學報, 41(17): 6871-6882.]
WANG Z, HUANG RF, WANG LH, et al., 2009. Analysis on life table of natural Sabina vulgaris population in MU Us Sandy Land [J]. J Desert Res, 29(1): 118-124.[王卓, 黃榮鳳, 王林和, 等, 2009. 毛烏素沙地天然臭柏種群生命表分析 [J]. 中國沙漠, 29(1): 118-124.]
WANG J, YAO L, AI XR, et al., 2020. Structure and dynamic characteristics of Betula luminifera populations in different regions of Southwest Hubei Province, China [J]. Chin J Appl Ecol, 31(2): 357-365.[王進, 姚蘭, 艾訓儒, 等, 2020. 鄂西南不同區域亮葉樺種群結構與動態特征 [J]. 應用生態學報, 31(2): 357-365.]
WU CZ, HONG W, XIE JS, et al., 2000. Life table analysis of Tsuga longibracteata population [J]. Chin J Appl Ecol, 11(3): 333-336.[吳承禎, 洪偉, 謝金壽, 等, 2000. 珍稀瀕危植物長苞鐵杉種群生命表分析 [J]. 應用生態學報, 11(3): 333-336.]
XIE TT, SU PX, ZHOU ZJ, et al., 2014. Structure and dynamic characteristics of Calligonum mongolicum population in the desertoasis ecotone [J]. Acta Ecol Sin, 34(15): 4272-4279.[解婷婷, 蘇培璽, 周紫鵑, 等, 2014. 荒漠綠洲過渡帶沙拐棗種群結構及動態特征 [J]. 生態學報, 34(15): 4272-4279.]
XIAO YE, ZHOU XY, ZHANG XQ, et al., 2007. Advance in research on dormancy and germination of Viburnum seed [J]. Seed, 26(6): 56-59.[肖月娥, 周翔宇, 張憲權, 等, 2007. 莢蒾屬(Viburnum)種子休眠與萌發特性研究進展 [J]. 種子, 26(6): 56-59.]
YAO L, AI XR, YI YM, et al., 2017. Structure and dynamics of dominant populations in the mixed forest of subtropical evergreen and deciduous broad-leaved tree species in the Southwest of Hubei Province [J]. Sci Silv Sin, 53(2): 10-18.[姚蘭, 艾訓儒, 易詠梅, 等, 2017. 鄂西南亞熱帶常綠落葉闊葉混交林優勢種群的結構與動態特征 [J]. 林業科學, 53(2): 10-18.]
YANG XB, LIU SX, YANG FS, 2000. A study on the structure of Davidia involucrata var. vilmoriniana population in Houhe, Hubei Province [J]. J Biol, 17(1): 16-18. [楊心兵, 劉勝祥, 楊福生, 2000. 湖北省后河自然保護區光葉珙桐種群結構的研究 [J]. 生物學雜志, 17(1): 16-18.]
YANG YH, GONG W, LI H, et al., 2015, Study on seed germination characteristics of 4 species of Viburnum [J]. Territ Nat Res Stud, (1): 88-90.[楊軼華, 宮偉, 李虹, 等, 2015. 4種莢蒾種子萌發特性研究 [J]. 國土與自然資源研究, (1): 88-90.]
YAO LJ, YAO L, YI YM, et al., 2017. Species composition and communty structure of a 6-ha subtropical evergreen and deciduous broad-leaved mixed forest dynamics plot in the Qizimei Mountains, Hubei Province [J]. Biodivers Sci, 25(3): 275-284.[姚良錦, 姚蘭, 易詠梅, 等, 2017. 湖北七姊妹山亞熱帶常綠落葉闊葉混交林的物種組成和群落結構 [J]. 生物多樣性, 25(3): 275-284.]
ZHU Q, AI XR, YAO L, et al., 2019. Structure and dynamics of Carpinus fargesiana population in southwest Hubei Province [J]. J Centr S Univ For Technol, 39(8): 93-100.[朱強, 艾訓儒, 姚蘭, 等, 2019. 鄂西南川陜鵝耳櫪種群結構與動態 [J]. 中南林業科技大學學報, 39(8): 93-100.]
(責任編輯?李?莉?王登惠)
