采用腦網絡分析的應激誘發腦-機接口運動感知與反饋激活時序規律研究
2023-10-29 10:06:48張凱徐光華李文平李輝杜成航韓丞丞張四聰謝杰仁
西安交通大學學報 2023年10期
關鍵詞:實驗
張凱,徐光華,2,李文平,李輝,杜成航,韓丞丞,張四聰,謝杰仁
(1. 西安交通大學機械工程學院,710049,西安;2. 西安交通大學機械制造系統工程國家重點實驗室,710054,西安)
腦-機接口作為一種新型的人機交互技術,可以在不依賴于腦的正常輸出通路(外圍神經和肌肉組織)的情況下,使人類大腦直接與計算機或其他外部設備進行通信,進而控制外部設備進行相應的任務并提供反饋[1]。其中,作為其重要工程的應用,運動誘發腦-機接口為腦損傷閉環神經協同康復提供了一種可行的技術手段,具體通過刺激范式有效誘發受試者的運動中樞使其產生神經響應信號,再通過信號處理技術對腦電信號進行實時解碼,然后將解碼得到的運動意圖轉化為外部設備的控制指令,輔助受試者完成運動輔助和康復訓練[2-4]。該過程有效模擬了運動行為控制的中樞響應過程,實現了神經沖動沿著運動感知環路—神經中樞—運動控制環路—受控肢體的傳導,并利用物理接口和雙向通信技術從大腦外部構建了一條閉環回路,進而實現運動控制環路的重建[5-6]。
基于腦-機接口技術的神經環路重建有效性受到多種因素的影響,其中,運動感知與控制的時序性對于接口性能與應用效能具有重要的影響。Hebb法則與脈沖時間依賴突觸可塑性規律(spike-timing dependent plasticity, STDP)表明:活動時序依賴的突觸可塑性滿足確定性的時間窗口,直接揭示了有順序的激活突觸前和突觸后神經元是修飾環路的關鍵,從微觀層面揭示了時序性對神經環路重塑的重要規律[7-9]。然而,傳統的誘發范式僅利用依賴主觀認知與弱視覺誘發信息來引導受試者產生自主想象[10-12],其不僅易受到生理狀態變化的影響,且難以保證運動感知與控制的同步性,易導致運動誘發腦-機接口的中樞響應過程在不同試次、不同時間具有高度變異性,直接造成整個交互過程中,運動感知神經與運動控制神經的激活過程難以保證同步性和有效時序性(如圖1所示),無法滿足感知反饋同步性和時效性要求,與STDP所揭示的規律相悖。由此,限制了運動誘發腦-機接口的應用性能,難以保證臨床應用中的康復訓練與治療效果。
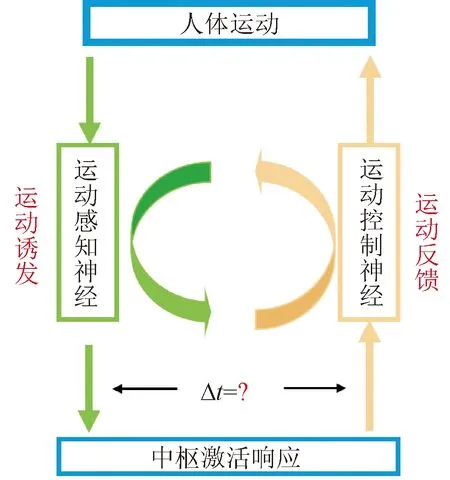
圖1 傳統運動誘發腦-機接口感知與反饋激活示意Fig.1 The diagram of perception and feedback activation of traditional brain-computer interface based on motor activation
探索運動感知環路與控制環路激活的強匹配誘發模式,對于增強閉環腦-機交互過程的同步性和時序性具有重要啟示。中樞激活響應效果往往取決于刺激范式的設計,作為一種典型的刺激模式,應激反應可以定義為各種刺激物(應激源)作用于個體而引起的非特異性反應。特定條件下的應激反應有助于提升自主注意力水平,促進情緒和思維的正向調整,并提升交感神經的激活程度,引起腦電信號功率的增強[13-14]。文獻 [15]研究表明,特異性的重心失衡應激刺激會引起運動中樞的激活,且該激活方法具有更強的激活程度和更有序的激活效果。因此,應激狀態的出發時刻引起受試者的必然性運動響應,為腦-機交互中運動感知環路與反饋控制環路的有序激活規律研究提供了一種可行的方法。同時,研究發現,大腦的信息加工和處理過程可以通過特定腦區之間信號同步強化效果來進行表征,其中,研究驅動源腦區和神經有向連接的相關規律對于理解大腦活動和神經調控的時序規律有著重要的作用[16]。腦功能網絡,是一種將圖論和復雜網絡理論應用于大腦活動定量分析的研究,作為一種描述大腦皮層不同功能區相互作用一種方法,可以有效幫助研究人員進一步了解腦內信息的加工機制和作用規律[17]。
因此,本文利用應激誘發腦-機接口技術與腦功能網絡分析手段,探索了運動激活響應的時序作用規律,研究了閉環腦機時序同步強化方法,為時序同步的腦-機接口設計提供有效理論與技術基礎。
1 應激誘發的時序激活響應檢測平臺設計與搭建
1.1 應激狀態觸發與運動響應關聯分析
應激誘發運動響應通常產生于受試者從非應激態轉變為應激態的瞬時時刻,此時刻對應的應激參數定義為應激觸發點(用失衡角度θ表示)。重心失衡作為一種典型的運動應激刺激,往往伴隨著前庭神經的激活,其作用過程可以描述為:前庭感受器受到偏離重力方向的信息時,神經中樞通過調節四肢的肌張力或控制肢體運動來調整身體的姿態和體位,以維持身體的平衡。如圖2所示,基于之前研究的重心失衡應激腦-機接口模型[15],受試者會利用肢體進行支撐動作,來防止重心的完全失衡,而這種運動本質上受到本能行為和自主意識的共同支配,因此具有確定的運動激活性。因此,這種運動響應的強關聯模式,有望提供一種可控型意念誘發模式。

(a)重心失衡應激平臺
(b)非應激態

(c)應激態
1.2 運動感知與反饋控制激活的時序信息檢測平臺設計
在運動調控的過程中,有效刺激的產生往往伴隨著感知神經的激活,運動動作的發生表征了運動控制神經的激活。對于自然的正常神經環路,運動感知與控制環路的激活幾乎是瞬時同步的,而腦-機接口構建的人工神經環路中,兩者的激活關系往往不同步,且缺乏有效的激活監測手段。
在真實運動任務下,真實運動的感知與控制反饋近乎同步,因此受試者在應激觸發點位置產生的真實運動會引起運動感知與反饋神經同時激活。對于腦-機交互任務,利用外界刺激誘發運動中樞,通過提取腦電激活特征來實施運動控制。由此可以類比得到,有效刺激感知輸入的時刻為運動感知環路的激活時刻,腦電激活特征的啟動時刻為運動控制環路的激活時刻。
由于受試者存在反應延遲及腦電的演變過程,腦-機接口任務中的感知激活與控制激活往往存在一定的時延。因此,通過記錄受試者在應激刺激下應激觸發點與腦電特征響應點對應的時刻,有望得到腦-機交互過程中運動感知與控制反饋環路激活的相對時序關系。為此,設計了如圖3所示的應激觸發點與運動誘發激活檢測的多源信息采集系統。

圖3 多源生理信息采集系統Fig.3 The acquisition system for multi-source physiological information
陀螺儀位姿傳感器通過對重心偏移的角度進行采集以捕捉應激點對應的角度和時刻參數,薄膜壓力傳感器通過對接觸力變化進行監測判斷肢體真實運動產生的時刻,腦電信號則是從中樞層面反映運動激活響應的動態變化。
為了減小測量過程的誤差,對不同傳感器之間時序標定進行統一標準化設置,以保證多源信息采集與任務狀態的變化一致性。同時,為保證隨動座椅與傳感器之間的實時通信,采用TCP/IP與串口通信完成系統控制的信號的實時處理,其具體框架如圖4所示。
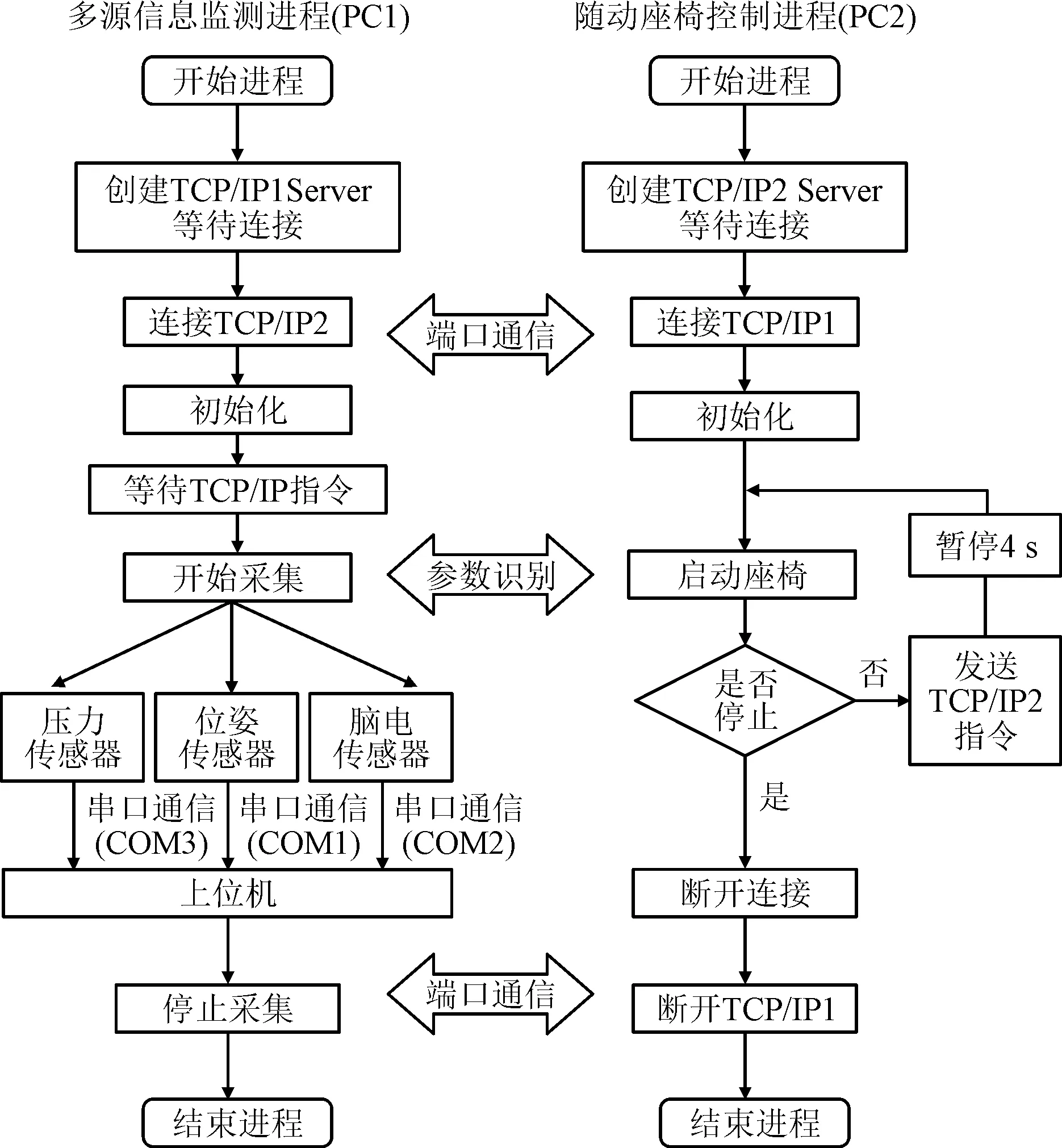
圖4 多源信息采集與運動誘發平臺軟件系統 Fig.4 The multi-source information acquisition and motion induction platform software system
隨動座椅與多源信號采集系統采用TCP/IP進行交互通信、接收信號、處理進程并記錄受試者生理信號隨座椅運動的動態過程。在真實運動或意念激活的信號檢測過程中:利用真實運動的接觸力變化對應旋轉座椅角度變化,可以得到反映感知神經激活的應激點參數;分析計算腦電信號的激活時刻,可以求得運動控制神經的激活時刻。
2 腦-機接口運動感知與反饋激活檢測方法設計
2.1 運動應激觸發點檢測實驗設計
為求解應激觸發點對應的刺激參數,設計了如圖5所示的實驗。

圖5 應激誘發-運動執行反饋實驗方案示意 Fig.5 The diagram of stress response induction and motor execution feedback
在運動執行實驗過程中,前2 s為運動準備階段,受試者在無物理限位的隨動座椅上保持靜止。從第2 s起,座椅隨機向左/右方向以10 rad/s的速度勻速旋轉。期間,要求受試者在無支撐接觸力的條件下,盡量保持身體平衡,直到座椅偏轉到失衡的臨界點,本能反應驅使受試者利用手部支撐座椅扶手,以保持身體平衡,該動作發生過程被布置于支撐點的壓力薄膜傳感器所記錄。在整個過程中,陀螺儀全程記錄不同時刻對應的偏移角度。
通過記錄支撐接觸力產生時刻的對應的偏移角度,即可確定引起應激狀態觸發的失衡角度θ,即應激點。由于本文的研究對象為左右手運動意念誘發任務,因此通過多次重復左右側運動偏轉實驗任務,將疊加平均后的結果定義為單側手的應激點。
2.2 腦電激活響應檢測實驗設計
相比真實運動,腦-機交互過程中感知神經的激活通常為外界刺激輸入的時刻,控制神經環路激活的判據通常為腦電激活特征的產生時刻,而這種模式下的感知與激活存在一定的時延。
因此,記錄應激點產生時刻與腦電特征啟動時刻的時延,為量化運動感知與反饋的激活同步關系提供了一種可行的方案。為此,根據運動執行實驗得到的應激點參數,設計應激刺激運動誘發實驗方案,如圖6所示。
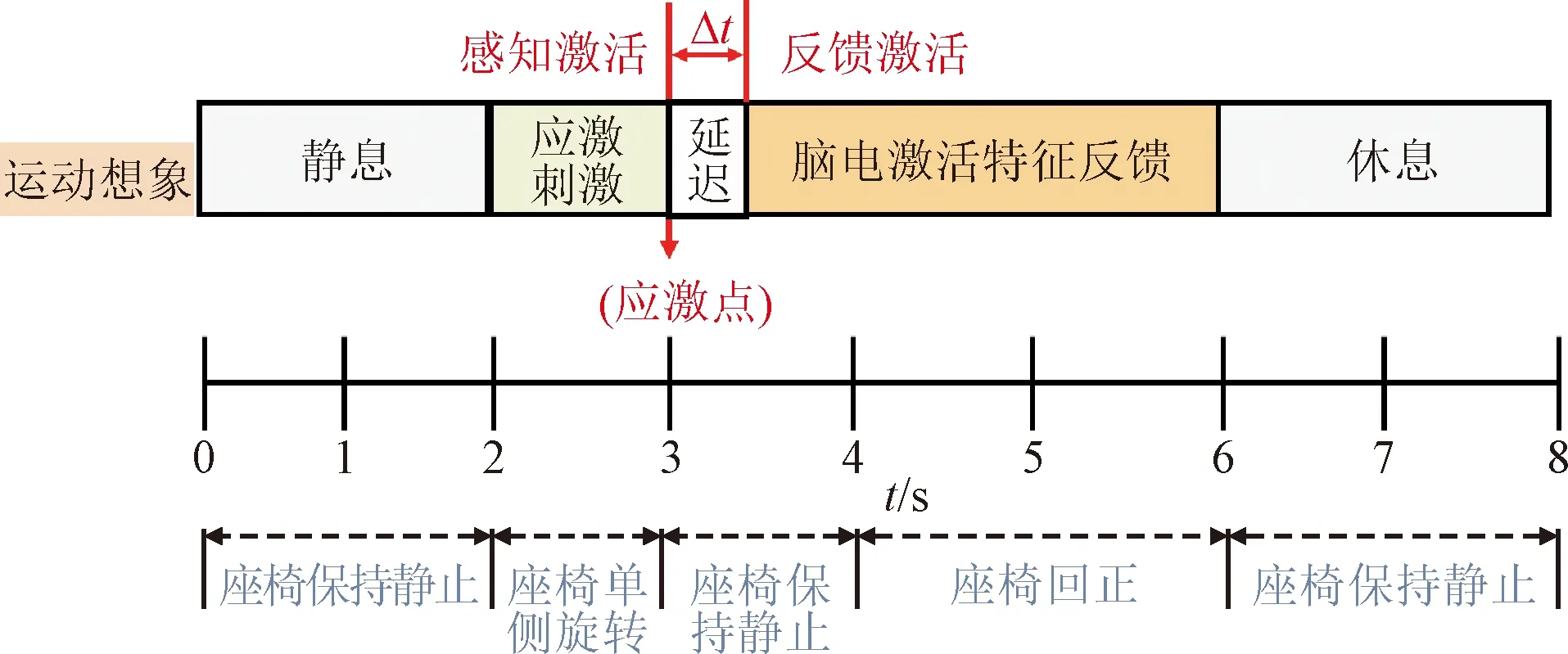
圖6 應激誘發-運動激活反饋實驗方案示意 Fig.6 The diagram of stress response induction and motor activation feedback
首先,利用運動執行實驗所求解的應激點參數對隨動座椅的偏移角速度、位移進行設置,具體通過調整隨動座椅的偏轉角速度ω;然后,從第2 s起,座椅啟動并在1 s內偏轉到應激觸發點的位置,此時采用安全限位裝置代替受試者的運動支撐,以保證受試者在實驗全程保持靜止;從應激點后,受試者執行自主意念誘發任務,直到第6 s,單次實驗結束。
2.3 實驗條件及腦電數據采集
為了探索時序依賴的腦電響應規律,構建感知同步運動誘發接口,招募了12名受試者(男7女5),身體健康無精神疾病,雙眼矯正視力正常,平均年齡(24±2.5)歲。所有受試者都被告知實驗的全部程序,并簽署知情同意協議。
實驗使用奧地利g.tec公司生產的g.Nautilus高精度無線生物電信號采集分析系統,可以用于運動狀態下的腦電信號采集,其內置阻抗檢測功能可測量電極與頭皮接觸電阻,精度達 kΩ 級,并具有硬件帶通濾波和陷波濾波功能。選取運動區域的相關電位活動作為分析對象,根據10/10 標準電極布置法則,在受試者頭部運動皮層相關區域布置電極,其分別位于FC3、C3、CP3、Cz、FC4、C4、CP4、CPz,參考電極位于右耳耳垂(A2),接地電極(GND)位于前額(FPz)。同時,保證所有阻抗保持在5 kΩ以下。腦電圖信號的采樣率設置為500 Hz,采用50 Hz陷波濾波器和3~100 Hz帶通濾波器濾波。在進行信號分析之前,對每個受試者不同任務的腦電信號進行疊加平均,以盡可能減少噪聲和異常值對分析結果的干擾。同時,去掉誤差較大和不可用的試次,以保證后續分析結果的準確性。
轉椅采用數字伺服控制器(Syntron-543AA0),精度為0.1°,配備三軸加速度計(witt-jy901),以記錄實際位移運動學。搭載壓力薄膜傳感器(IMS-S08A),最大量程可達40 kg。所有的運動參數(旋轉角度/角速度)都可以手動指定。
實驗時,每個受試者要依次參與3類實驗:運動執行,應激刺激運動誘發(VSMI)與無應激運動想象(MI)實驗。其中,VSMI實驗中提供應激刺激和視覺提示,MI實驗中僅供座椅旋轉的視覺動畫作為提示。運動執行實驗一共分為3組,每組完成 10次重復實驗,每組間隔2 min。兩種運動誘發實驗各需要完成6組實驗,每組開展 10 次重復實驗,每組間隔3 min。當受試者在參與腦電采集實驗過程中,應盡量避免肢體或者頭部運動、吞咽口水、咬牙。實驗過程中,實時記錄腦電數據、壓力數據、位姿數據、時標信息。
3 基于腦網絡分析的時序依賴規律量化方法研究
由于EEG能夠直接測量神經活動且具有高時間分辨率,探索基于EEG的跨腦區功能連接對理解大腦運動功能的調控機制具有重要的作用。研究表明,特定腦區信號之間的同步強化增強作用可以通過腦功能性連接進行表征[18],而這種連接關系的動態特征能夠通過多項指標進行定量評估。在腦電信號分析的領域,研究人員提出了多種算法來定義信號之間的功能連接指標,常見的如皮爾遜相關系數、波譜相干、互信息、相滯指數等[19-20],其中,有向傳遞函數是一種用多變量自回歸模型來表征腦電信號功能性連接強弱的方法,并遵循格蘭杰因果關系多變量有效測量規律[21]。其主要通過計算單通道輸入信息和輸出信息的信息量的關系,來衡量單通道對其他通道的信息貢獻量關系,進而定性判斷不同腦區響應之間相互聯系與影響。本節通過引入腦網絡分析方法,利用MATLAB的econnectome工具箱,對應激點誘發下中樞響應時序演化規律進行研究,以揭示應激調控的內在作用機制,驗證所提腦-機接口方法對于中樞響應同步性增強的有益效果。
首先,利用多變量的自回歸模型(multivariate autoregressive, MVAR)來描述一個N通道的EEG信號[22],X(n)=[x1(n),x2(n),…,xN(n)]。利用靜息態和激活態的腦電信號計算激活比E
(1)
(2)
式中:E為激活能量比;A為激活態腦電信號的能量;R為靜息態腦電信號的能量;m為運動激活啟動時間點;n為單試次總時間點;r=1,2,3,…,p為MVAR的階數;Ar為系數矩陣;w(n)為高斯白噪聲。
然后,將所求系數矩陣與信號表示投影到頻域,得到
W(f)=Ar(f)X(f)
(3)
(4)
計算系統傳遞矩陣D(f)
(5)
式中D(f)元素Dij表示系統中第j個輸入與第i個輸出之間的關系。
將EEG中第j通道對第i通道的影響關系定義為
(6)

由此,計算單通道的傳遞函數與所有相關通道的元素平方和,即可得到有向傳遞函數的表達式
(7)

(8)

有向傳遞函數對通道之間信息交流進行了定量的評估,為了進一步挖掘運動激活響應過程腦網絡的動態屬性,本文采用圖分析的方法計算響應的加權網絡性質,旨在實現對大腦激活狀態時序規律的捕捉。作為一種定量評估腦網絡局部信息處理的效率的指標,聚類系數常常被用來表征網絡功能分化能力以及評估網絡集群程度[23],而運動皮層腦區的激活程度,往往也伴隨著局部信息處理效率的增加而增加。因此,選取聚類系數C作為腦激活狀態動態評估的指標之一,計算式為
C=
(9)
式中:i、j、h分別為腦網絡中的任意節點;ωij為節點i和節點j之間的連通性強度;z為網絡節點數;Z為網絡節點的集合。
特征路徑長度L是一種評估網絡功能整合能力的常用指標[24],其定義為網絡中任意兩個節點之間的平均值。特征路徑長度越短,網絡功能整合能力就越強,網絡連通性越好。特征路徑長度計算式為
(10)

全局網絡效率EG作為一種表征網絡功能集成屬性的指標[25],數值大小與路徑長度呈反比,可以更加直觀地反映腦網絡全局信息的傳輸能力,計算式為
(11)
作為描述大腦不同腦區之間局部信息傳輸能力的重要指標[26],局部效率EL可以表征網絡功能分離的程度,進而對不同區域活動的規律進行評價,計算式為

(12)
4 實驗與結果
腦網絡功能連接能夠有效反映運動信息加工處理過程和不同腦區的協同工作機制,以進一步揭示運動行為控制任務下的大腦的工作機制。為此,采用腦網絡分析方法進一步探索運動激活響應的動態演化過程。
雖然本文僅對8通道腦電信號進行腦網絡分析,但該布置對負責運動皮層的主要通道(FC3、C3、CP3、Cz、FC4、C4、CP4)進行了覆蓋,且能保證對運動誘發任務中對側運動皮層之間的信息交互規律捕捉,因此具有可行性。同時,由于應激刺激對于其他皮層的誘發響應規律尚不明確,為了避免其他腦區激活響應對運動皮層的分析帶來干擾,故未擴大通道的選擇范圍。
4.1 腦功能網絡連接分析
腦功能連接反映中樞激活的主要表現形式,可以描述為:核心激活區將作為質心樞紐,與其他相關區域建立連接,協同完成信息加工。在由左右手運動誘發/執行所引起的運動激活過程中,靜息態下的兩側腦網絡呈現對稱態,單側手運動激活態下的網絡呈現偏側化形態,通常表現為對側運動皮層激活,與同側區域建立有向連接。
為了評估本小節所提出方法對運動刺激的激活響應規律,本文通過對應激刺激運動誘發過程的網絡連接的動態過程進行可視化,用以分析運動響應的感知同步關系,評估結果如圖7所示。

(a)左手運動誘發腦網絡激活變化規律

(b)右手運動誘發腦網絡激活變化規律

(c)左手運動想象腦網絡激活變化規律

(d)左手運動想象腦網絡激活變化規律
對同一任務下的腦電信號進行疊加平均,提取μ頻段(8~12 Hz)和β頻段(13~30 Hz)的腦電信號作為分析對象。如圖7(a)所示:在運動準備階段,左右兩側枕葉皮層產生激活,網絡結構呈現對稱狀態;應激刺激開始,旋轉座椅向左側偏轉,運動皮層網絡結構由對稱態變為偏側化,右側運動皮層的CP4、C4通道作為質心通道,控制信息流向其他區域流出,完成對運動信息的協同處理;達到應激點后,停止外源刺激,應激響應占據主導思維活動,此時腦網絡連接密度增加,皮層激活逐漸達到峰值;隨著激活過程的結束,腦網絡由偏側化恢復到對稱態。圖7(b)反映了右手運動激活的腦網絡變化規律,激活質心為左側腦區的C3、CP3通道,演化過程與7(a)所示規律具有一致性。由此可以說明,偏側化重心失衡刺激會誘導受試者產生與運動誘發/運動執行相同的腦網絡模式,當失衡點達到應激觸發點后,該運動激活引起的網絡連接和信息交流會更加密集。由此可以說明,應激觸發不僅與傳統運動中樞激活具有一致性,而且能有效強化運動腦區之間的信息交流和傳遞。
4.2 基于腦網絡動力學特性的響應規律表征
腦網絡的拓撲結構可以直觀反映中樞響應的演化過程,而網絡參數的量化可以實現對中樞響應時序關系的表征。因此,通過量化腦網絡的聚類系數、特征路徑長度、全局效率與局部效率在誘發任務的動態參數變化來表征運動刺激與中樞激活程度的時序變化規律,具體結果如圖8所示。

(a)聚類系數
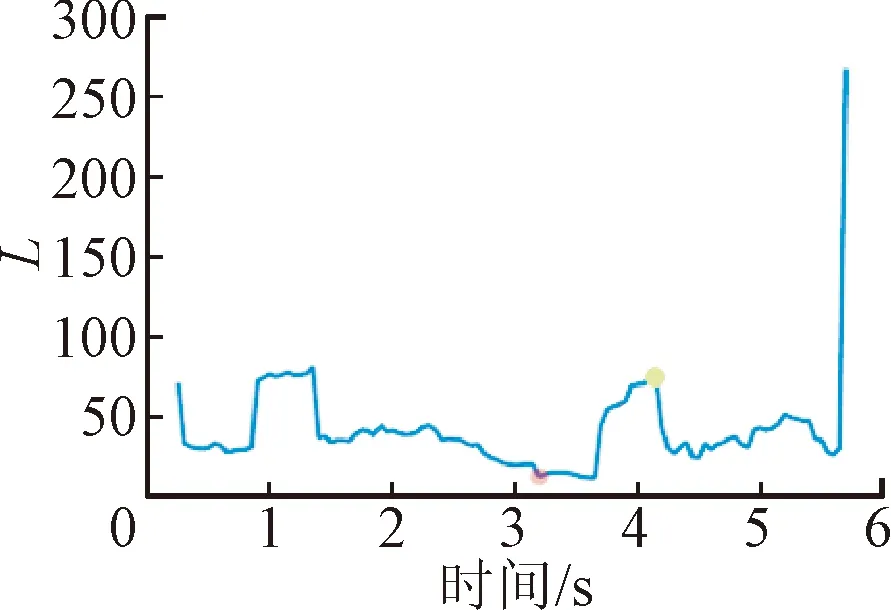
(b)特征路徑長度

(c)全局效率

(d)局部效率
圖8給出了1名受試者所有數據疊加平均后的網絡參數變化。可以看出,在運動準備階段(0~2 s),腦網絡的聚類程度和信息處理效率較低;隨著應激刺激的啟動(2~3 s),C、EG、EL曲線先下降而后急劇上升,表明大腦在受到應激事件后,先表現出一定延遲,然后運動皮層之間信息加工在短時間內迅速增加;達到應激點后(3~4 s),運動激活主導此刻的思維活動,該時間段內的C、EG、EL曲線小幅度下降后又持續攀升,并達到峰值,峰值時刻代表了運動皮層激活最強的狀態;運動誘發任務結束后(4~6 s),隨動座椅回正,大腦信息加工效率逐漸從峰值降低,隨著受試者回到靜息態,恢復到與運動準備階段相同的水平。不同于上述3個參數,特征路徑長度表征了網絡功能整合能力,長度越短,整合能力越強。雖然變化規律與上述3個參數相反,但揭示的規律具有一致性。
4.3 感知反饋激活時序規律的量化統計分析
本小節對12名受試者的應激點參數、激活響應時延以及激活最大持續時間進行統計。應激點參數和運動響應的激活時延由量化分析方法[15]得到,最大激活持續時間通過腦網絡動力學參數變化規律得到,即記錄激活啟動點(圖8中紅色標識點)與響應峰值(圖8中綠色標識點)對應的時間區間。對疊加平均后的信號進行統計分析,結果如表1所示。
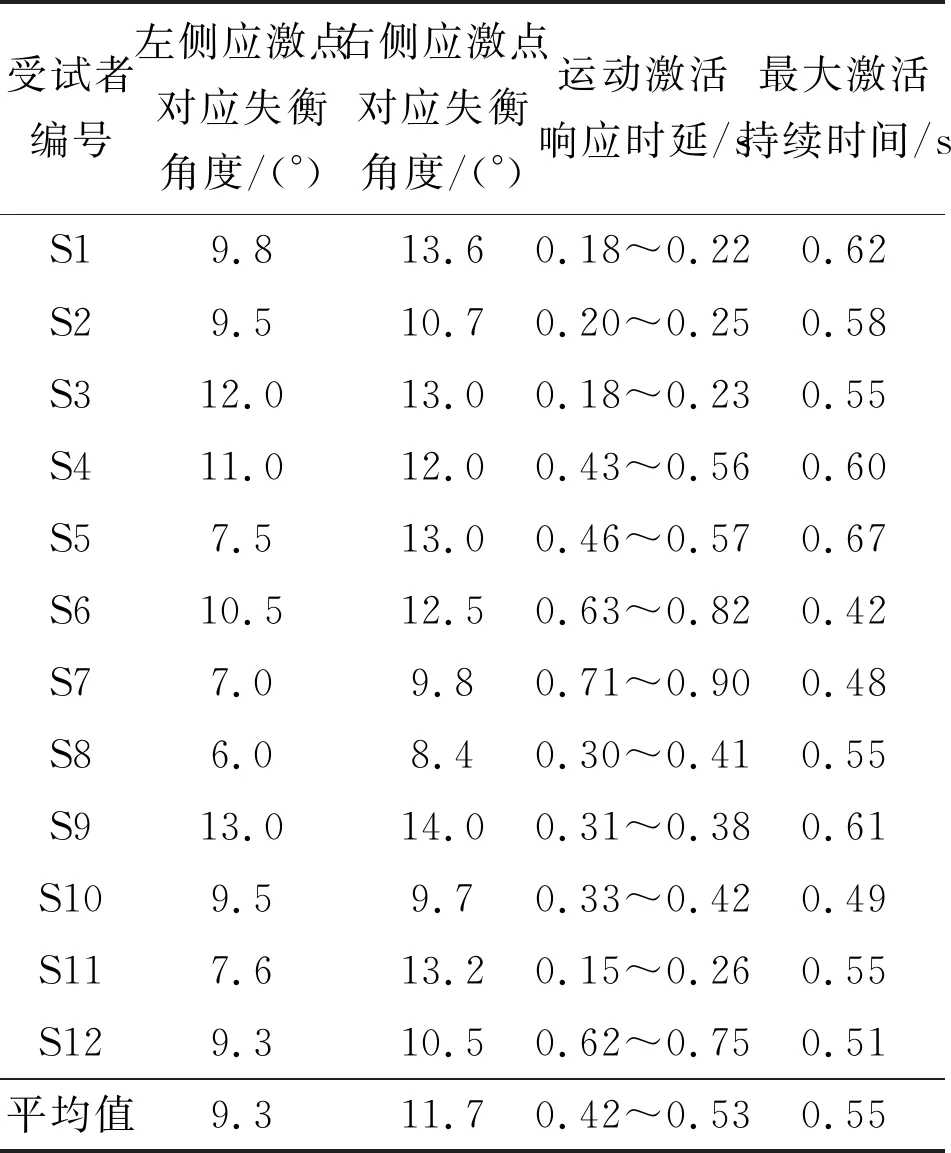
表1 應激刺激中樞響應參數
由統計結果可知,同一受試者的左右側應激點都具有差異,表明了受試者在失衡刺激下左右側運動響應水平具有一定的差別,且右側身體平衡控制能力普遍優于左側平衡的控制,這可能取決于個體的利手特性。雖然本實驗未將利手因素作為受試者篩選的條件,但是由于人群中右利手的人數普遍大于左利手人數,所以受試者群體的右側平衡控制能力優于左側平衡控制能力,也符合右利手在人群中數量占優的規律。不同受試者自主意念誘發的響應時延規律具有差異性,即感知與反饋響應的時延為0.15~0.90 s不等,但最大激活響應時間的持續時間具有一致性,為400~600 ms。不難理解,由于受到個體差異的影響,不同受試者對于應激的響應水平與腦-機交互的操作能力都具有一定的差異性,這也決定了外部“神經搭橋”感知-激活的延遲特異性。不過,有效的激活響應量化結果可以指導腦-機接口外源刺激、系統響應,輔助訓練參數的更新優化,以達到誘發可控、時序同步的交互效果。
4.4 應激觸發激活響應同步增強效果驗證
為了驗證應激觸發點誘發對于閉環調控同步增強的效果,本小節分析對比應激觸發運動誘發與無應激運動想象實驗結果。實驗通過有應激和無應激的外界刺激誘發受試者產生中樞的激活響應,利用不同誘發模式下感知反饋激活響應時延,驗證應激觸發的同步增強效果,分析結果如圖9所示。

圖9 不同誘發模式下感知反饋激活響應時延Fig.9 The delay for motor perception and feedback activation in different evoked pattern
由分析結果可知,所有受試者在應激觸發點后的1 s內都產生了運動激活響應,12名受試者的感知反饋平均時延為0.629 8 s,而傳統的運動想象平均響應時間長達1.782 9 s。除受試者S6以外,兩種范式對于其他所有受試者的響應都具有顯著性差異(p<0.01),且應激刺激的誘發響應同步性在所有受試者之間都高于傳統的運動想象,由此證明了應激刺激對于運動激活響應的有序性調控具有積極的影響。受試者S6在應激誘發任務中產生的效果沒有優于傳統想象,可能的原因是由于受試者尚未在訓練過程中適應失衡應激的刺激模式,進而導致其無法利用應激刺激完成主觀運動意念的激活。也可能是在失衡刺激中產生了身體運動,進而給腦電信號帶來了較大的串擾。
圖9表明,應激運動誘發相較于傳統的運動想象具有更強的誘發效果,結合上文中提到應激刺激的作用效果,其可能原因是:在重心失衡應激刺激下,受試者的自主神經運動控制會對重心失衡的狀態產生應激反應,進而導致其前庭神經以及負責運動感知和決策的前額葉在應激刺激過程中產生激活,而伴隨著受試者主觀運動意圖的響應,運動皮層隨之激活,在前額葉的激活前提下,該運動激活效果可能具有疊加效果,進而產生誘發增強的效果。
5 結 論
本文面向運動誘發腦-機交互過程中運動感知環路與反饋環路激活的時序規律展開研究,通過應激運動誘發腦-機接口平臺,開展重心失衡任務下的運動誘發激活時序檢測方法,研究基于腦功能網絡的激活時序量化規律,揭示了應激刺激腦-機交互過程中,運動感知激活與控制激活的時序規律和腦網絡動態演化過程。
腦功能網絡分析結果表明,應激刺激運動誘發的網絡動態演化規律與傳統運動激活響應的演化規律具有一致性,且在達到應激觸發點后,運動中樞的網絡連接密度和信息交流會變得更加密集,其動力學特性表征的腦激活響應更加強烈。通過檢測應激觸發點與腦電激活響應點,可以實現腦-機交互任務中,感知神經與運動控制神經激活時延的有效量化。在線實驗結果表明,負責運動控制的腦電特征一般會出現于運動感知激活后的0.15~0.90 s內,具體時延在不同受試者之間具有差異性,而其最大激活響應的持續時間為400~600 ms,在不同受試者之間具有一致性。同時,基于應激觸發的運動意念誘發模型,可以有效提升感知環路與控制環路神經激活的同步性,且該同步性的增強效果具有顯著性。
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
