柏木無性系和家系含碳量的早期評價與優良品系選擇*
2023-10-27 09:49:40邱勇斌鄭成忠王文月金國慶周志春
林業科學 2023年9期
楊 濤 邱勇斌 沈 漢 鄭成忠 張 振 王文月 金國慶 周志春
(1. 中國林業科學研究院亞熱帶林業研究所 浙江省林木育種重點實驗室 杭州 311400;2. 華中農業大學 武漢 430070;3. 浙江省開化縣林場 開化 324300)
全球森林儲存著近3千億t碳,林木是森林生態系統中最重要的碳匯,在維持全球碳平衡、減緩CO2濃度上升等方面起著重要作用(郭金堂等,2016)。森林的適應性取決于林木基因型對未來氣候變化的反應,對林木生產力的形成和固碳能力具有重要影響,國內外學者研究輻射松(Pinus radiata)、火炬松(Pinus taeda)、落葉松(Larix gmelinii)、冷杉(Abies fabri)和白樺(Betula platyphylla)等樹種的生物量累積與分配、含碳率及含碳量表明,固碳能力在不同樹種、同一樹種不同品種、樹齡或器官間、不同立地間均存在顯著差異(Eliaset al., 2003;Zhanget al., 2009;賈慶彬等,2014;Wanget al., 2015;Donget al., 2015)。同一樹種含碳量差異主要是因種源、家系、無性系和個體間生長速度、生物量與含碳率的差異引起,選育林木良種造林可以顯著提高碳匯價值和經濟效益。目前,較為精確的含碳量測量方法是在測定各器官生物量和含碳率的基礎上推算出來的,由于該方法耗時耗力,且對林地具破壞性(林瑋等,2020),難以應用到大群體樣本,而利用異速生長方程估算含碳量是常用的替代方法(Widagdoet al., 2020;張悅等,2022)。
家系和無性系是當前主要造林樹種的良種推廣方式。無性系綜合利用加性與非加性遺傳效應,具有較高的遺傳增益,而全同胞和半同胞家系僅分別包含1/2和1/4的加性方差(Wenget al.,2008;康向陽,2019);同時,通過無性系育種可獲得目標性狀表現突出的林木新品種,是利用群體中遺傳增益最為顯著的個體而不是群體平均值,具有家系林業無法比擬的優勢(Wenget al., 2008);并且,無性系林木的生長性狀表現穩定、一致,便于集約化經營,可大幅提高林業管理效率(Wu, 2019)。但從育種研究的角度,同一樹種無性系與家系的固碳能力是否存在差異還有待揭示。
柏木(Cupressus funebris)是栽培歷史悠久的生態和用材樹種,主要分布在我國長江流域及以南地區,其材質優良,耐瘠薄,喜鈣質土,適生性和涵養水源能力強,是石漠化地區和石質山地生態修復的先鋒樹種,近年來在我國南方廣泛推廣栽植,為速生優質高固碳品種選育提供了巨大空間。本研究項目組自20世紀90年代開始收集全國主產區的柏木優樹資源,系統評價育種群體的遺傳多樣性,選育出一批適生性強的優良種質,從種子園豐產、育苗技術以及“采穗圃營建-截頂促萌-幼化-扦插”等方面實現了柏木苗木的規模化繁育(樓君等,2014;Yanget al., 2016;鄭一等,2020)。本研究以營建在浙江省開化縣林場9年生柏木無性系和家系試驗測定林為對象,探究其生長、生物量、含碳量及其分配情況,篩選并驗證通用異速生長方程的準確性,嘗試評價不同基因型的固碳潛力,以期為我國南方高固碳柏木優良品系選擇及其推廣應用奠定理論基礎。
1 材料與方法
1.1 試驗材料與栽植
柏木無性系和家系試驗測定林來自浙江省開化縣林場,地理坐標為118°24′24″E,29°8′17″N。屬亞熱帶季風性氣候,年均氣溫16.4 ℃,晝夜溫差平均10.5 ℃,年均降雨量1 814 mm,年均日照時數1 679.8 h,無霜期252天。試驗點為柏木在該地區造林的代表性立地,其中,家系測定林材料共34個系號,包括33個家系(1代無性系種子園中分單系采集的種子)和1個對照(CK1,1代無性系種子園中的混合種子),于2012年3月在苗圃內播種育苗;無性系測定林材料共36個系號,包括35個無性系(1代無性系種子園中采集種子育苗后選擇的優良單株)和1個對照(CK2,1代無性系種子園中的混合種子),于2012年5月選取10 cm長半木質化插條在苗床上扦插育苗。2013年營建試驗測定林,采用隨機完全區組設計,設置5個試驗小區,10株小區雙行排列,株行距2 m×2 m。
1.2 生長調查與生物量取樣
1.2.1 生長調查與材積計算 9年生時,全林調查樹高(tree height,H, m)、胸徑(diameter at breast height,DBH, cm)、枝下高( height under branches, UBH, cm)和冠幅(crown width CW, m),單株材積(volume,V, m3)利用公式V=0.000 057 173 591×DBH1.31.8813305×H0.9956884計算(楊育林等,2014)。
1.2.2 取樣與生物量測定 選取樹高、胸徑、材積、枝下高和冠幅作為評價指標,設置各性狀權重依次為1.5、1.5、1.5、1.0和1.0,采用TOPSIS綜合評價法(Meshramet al., 2020)分別對無性系和家系試驗林各系號的生長性狀優劣進行排序,綜合篩選出優、中、差3個等級,在3個等級類型中各選擇3個無性系(家系),每個無性系(家系)有3個區組,在每個區組中選擇3個單株,無性系和家系分別伐取樣木81株。
生物量測定分為地上和地下2部分。樹木伐倒后,樹干部分每1 m切斷并稱重記錄,之后每段裁取5 cm圓盤,分別測量每個圓盤(帶皮)的鮮質量,標記裝袋;樹冠分上、中、下3部分,分別測量每部分枝和葉的總質量,每部分選取3條有代表性的枝條,分離枝葉并稱重、裝袋、標記;地下生物量測定中,按大小分為粗根(直徑>2 cm,包含根樁)和細根(≤2 cm),稱重并取樣(Zhuet al.,2013)。將所有樣品帶回實驗室,于80 ℃烘箱烘至恒重,測量各部位樣品干質量,計算含水率(%),然后根據各器官鮮質量乘以含水率分別得出樹干干生物量(trunk biomass, TB, kg)、樹枝干生物量(branch biomass, BB, kg)、樹葉干生物量(leaf biomass, LB, kg)和根干生物量(root biomass, RB, kg),最后得出單株總干生物量(total biomass of a single tree,TBS, kg):TBS=TB+BB+LB+TRB。
1.2.3 含碳率與含碳量測定 利用Elementar Vario EL(Germany)元素分析儀測定各器官含碳率(C, %),采用木材總碳(total carbon, TC)含量測定方法,溫度設置為1 100 ℃,進樣量為20 mg(納曉瑩,2016)。根據各器官含碳率和干生物量分別得出樹干含碳量(trunk carbon stock, TCS, kg)、樹枝含碳量(branch carbon stock, BCS, kg)、樹葉含碳量(leaf carbon stock, LCS, kg)以及樹根含碳量(root carbon stock, RCS, kg),則單株總含碳量(total carbon stock of a single tree, TCSS, kg)計算公式為:TCSS=TCS+BCS+LCS+RCS。
1.3 數據處理與分析
1.3.1 方差分析和遺傳參數 采用以下線性混合模型對各性狀進行生長差異分析:
式中:μ為總體平均值;Fi為無性系或家系間效應;Bj為區組間效應;eijk為隨機誤差。
式中:δp為標準差;為性狀平均值。
1.3.2 異速生長方程構建 異速生長方程采用冪函數(Y=aXb)的對數變換形式(lnY=a+blnX),由于模型存在異方差,使用對數形式模型是消除異方差現象的方法之一(張悅等,2022)。在多元回歸模型中,自變量間可能高度相關,導致回歸系數不穩定,使用對數形式模型進行線性回歸,可通過方差膨脹系數(variance inflation factor, VIF)衡量多重共線性嚴重程度,從而對自變量做出取舍(Linet al., 2017; Moussaet al., 2018)。本研究選用以下5個方程:
式中:TCSS為各器官的含碳量;k0、k1、k2、k3為擬合系數;D為胸徑;H為樹高;UBH為枝下高。式(2)、(3)和(5)通過VIF衡量多重共線性程度(Dutcet al.,2018);采用決定系數(R2)和均方根誤差(root mean square error, RMSE)進行模型效果評價和選優。
統計分析采用SPSS 26.0軟件進行,TOPSIS綜合評價應用DPS 14.0軟件。
2 結果與分析
2.1 生長性狀差異分析和遺傳參數估算
無性系的平均樹高、胸徑和單株材積分別為6.09 m、6.15 cm和0.010 6 m3,分別高出家系水平47.81%、28.39%和120.83%,家系水平的表型變異系數高于無性系,說明家系具有較高的遺傳選擇響應。樹高、胸徑、單株材積、枝下高和冠幅的無性系重復力幅度為0.651~0.756,家系遺傳力為0.132~0.322(表1)。

表1 柏木無性系和家系生長性狀的差異分析和遺傳參數估算①Tab. 1 Difference analysis of growth traits and estimation of genetic parameters of cypress clones and families
方差分析結果表明,柏木無性系間與家系間各生長性狀均存在極顯著差異(表1)。根據TOPSIS綜合評價結果(表2),無性系中排名1~12為優等級、排名13~24為中等級、排名25~36為差等級,其中優等級的Ci均值是中等級的159.49%、是差等級的298.15%;家系中排名1~11為優等級、排名12~22為中等級、排名23~34為差等級,其中優等級的Ci均值是中等級的153.46%、是差等級的317.89%。結合保存狀況,進一步選擇出優等無性系為C6、C9、C14,中等無性系為C1、C5、C11,差等無性系為C10、C20、C34,優等無性系的胸徑均值和樹高均值分別是中等無性系的136.90%和113.84%、是差等無性系的182.89%和131.43%;選擇優等家系為F8、F18和F24,中等家系為F5、F10和F22,差等家系為F11、F26和F32,其中,優等家系的胸徑均值和樹高均值分別是中等家系的123.73%和116.32%、是差等家系的143.14%和126.99%。
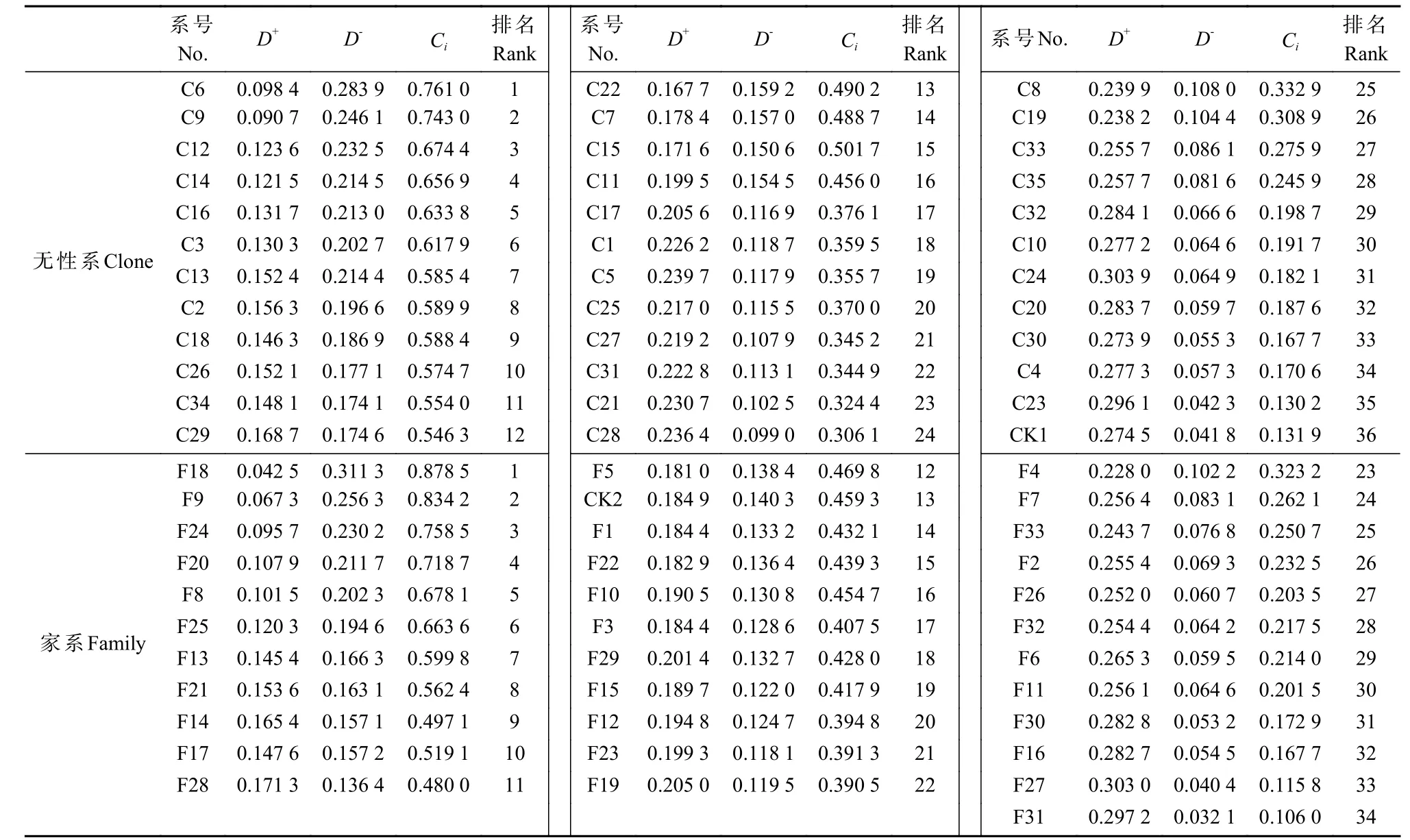
表2 TOPSIS法無性系和家系各系號優劣順序評價結果①Tab. 2 Obtaining the superior and inferior sequence of clone and family numbers by TOPSIS
2.2 生長性狀與生物量、含碳量、含碳率的關系
由圖1、圖2可知,無論家系還是無性系水平,柏木胸徑和樹高均與單株總生物量存在顯著線性關系(P<0.01),與單株總含碳量也存在顯著線性關系(P<0.01),可見,生長較好的基因型其生物量和含碳量也較高。胸徑和樹高與含碳率均無顯著線性關系,說明含碳率受生長狀況影響小。
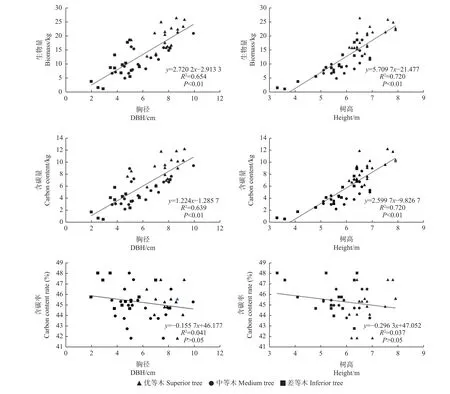
圖1 柏木無性系樹高和胸徑與生物量、含碳量、含碳率的線性關系Fig. 1 Linear relationship between height and DBH of cypress clones and biomass, carbon storage and carbon content
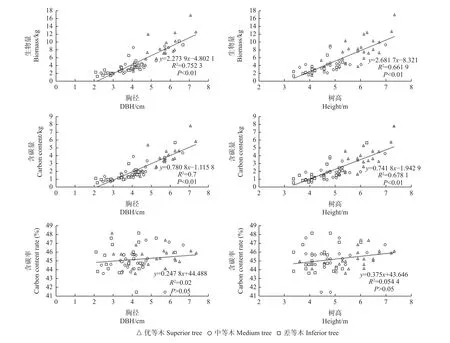
圖2 柏木家系樹高和胸徑與生物量、含碳量、含碳率的線性關系Fig. 2 Linear relationship between height and DBH of cypress families and biomass, carbon storage and carbon content
2.3 生物量和含碳量的分配模式
由表3可知,柏木無性系地上部分生物量分配相比家系高2.12%,其中,地上部分樹干生物量分配比例最高(44.36%),但較家系低0.60%,而無性系枝生物量分配比例為18.45%,較家系高5.55%,葉生物量分配比例為23.22%,較家系低2.84%;無性系地下部分生物量分配比例相比家系低2.12%,其中,粗根生物量分配比例低出家系3.45%,而細根生物量分配比例高出家系1.34%,說明無性系主要是以犧牲地下粗根的生物量分配、增加地上部分樹枝生物量和細根生物量來維持生長優勢。
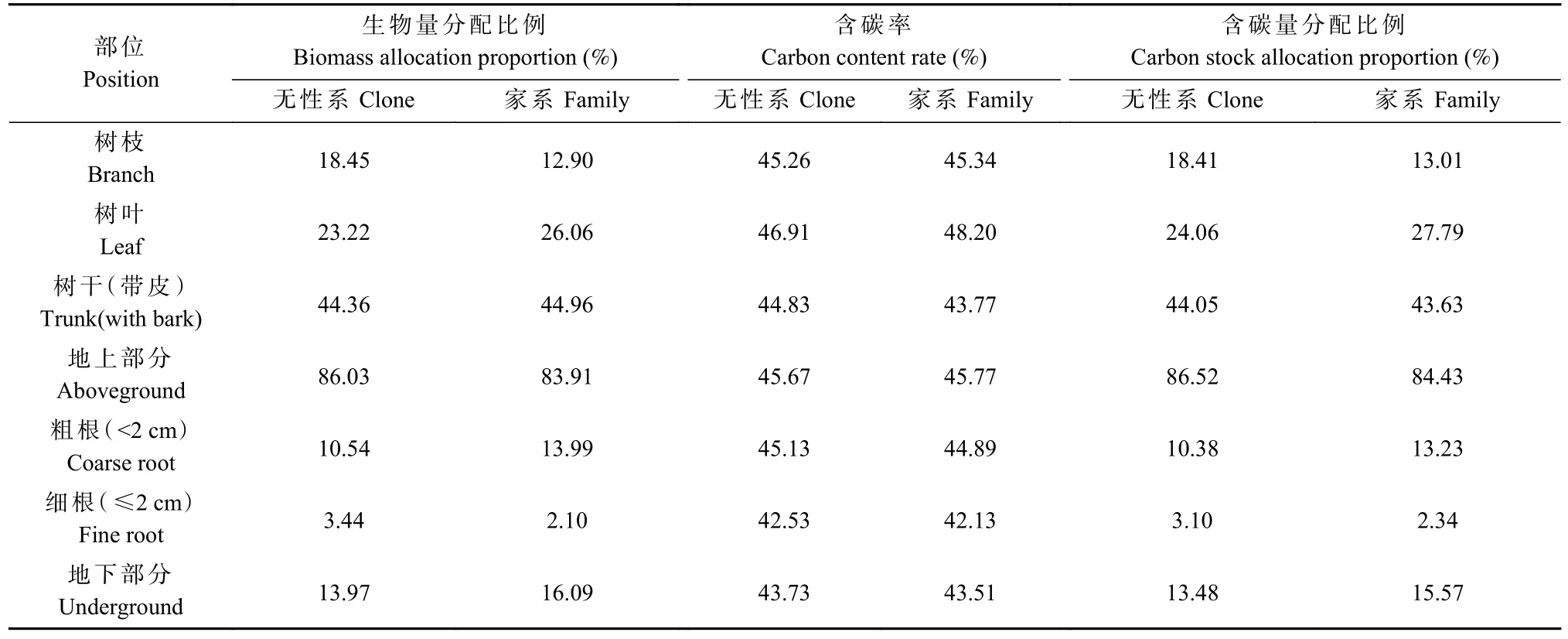
表3 柏木各器官生物量、含碳率、含碳量分配特征Tab. 3 Biomass, carbon content rate and carbon storage distribution of cypress organs
無論無性系還是家系,柏木各器官含碳率排序均為葉>枝>粗根>樹干>細根。無性系各器官含碳量占比排序為樹干>樹葉>樹枝>粗根>細根,家系各器官碳含量占比排序均為樹干>樹葉>粗根>樹枝>細根,說明樹干和樹葉是柏木含碳量的主要部分。整體上,無性系樹干、樹枝和細根的含碳量分配比例高于家系,而樹葉和粗根的含碳量分配比例低于家系。
2.4 含碳量異速生長方程的建立
構建單株總含碳量及其各器官含碳量與生長性狀之間的異速生長方程發現,以胸徑為單一變量擬合效果最差(式1),當以樹高為第二自變量加入模型(式2),模型精度有所提高,但幅度有限(表4)。當以樹高、胸徑、枝下高為獨立變量時,估算的含碳量(除無性系樹根含碳量外)精度較高,即模型(3)的R2達到最大,RMSE達到最小,且方差膨脹系數(VIF)較小,在2.977~3.982之間,不存在共線性問題。而當樹高與胸徑以D2H形式整合成一個參數作為自變量時(式4),與模型(3)相比,無性系和家系的單株總生物量、地上和地下部分生物量擬合度均有所下降,將枝下高作為獨立變量加入模型(4)后,R2均有提高,說明枝下高作為獨立變量加入模型對含碳量模型的擬合度影響較大。此外,利用模型(2)和模型(3)估算的無性系樹根含碳量精度相近,R2均為0.880,模型(2)的RMSE和VIF較模型(3)分別下降3.06%和33.31%。綜合R2、RMSE以及VIF表現,篩選模型(3)為柏木生物量的最優擬合模型。
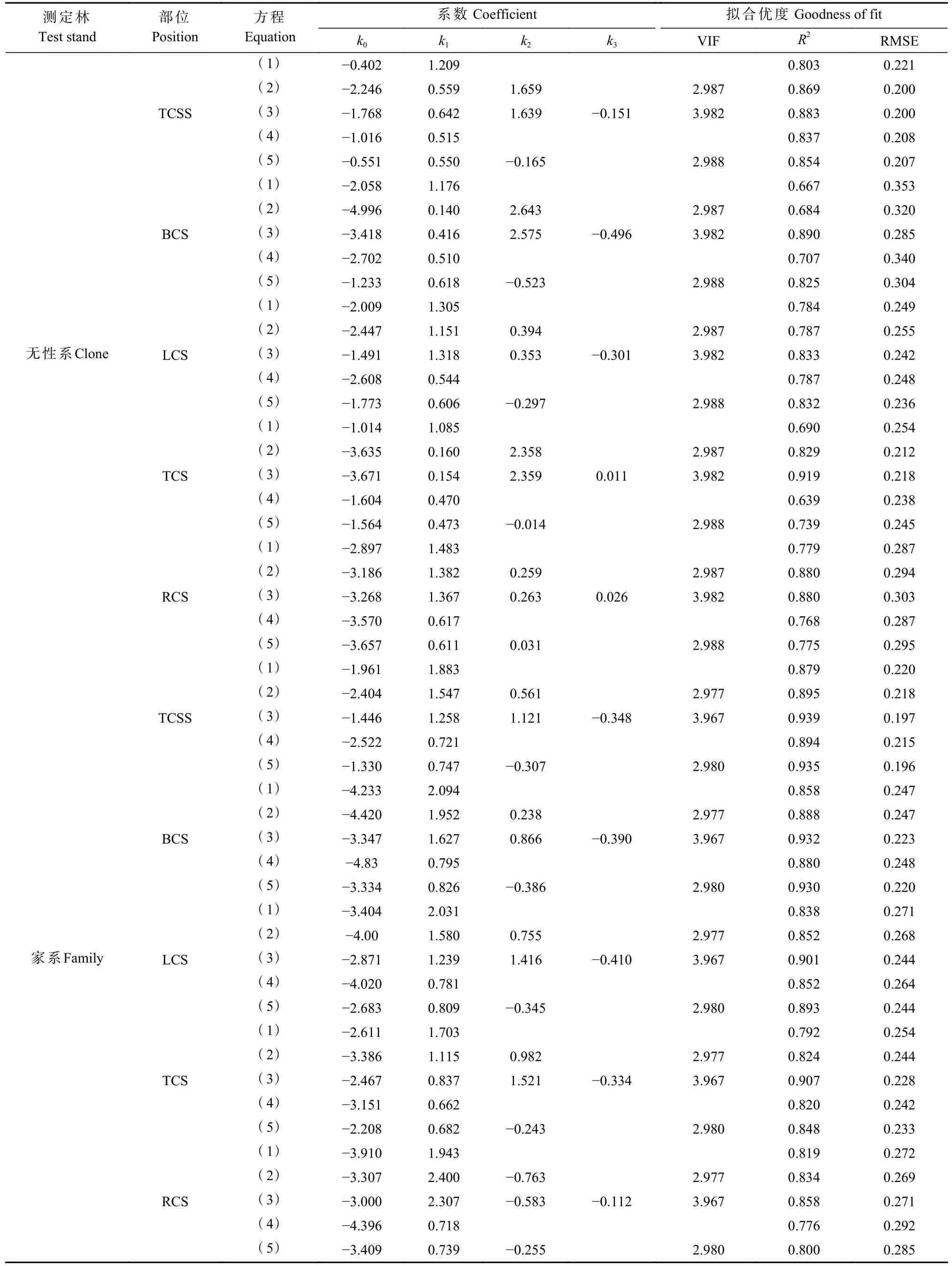
表4 柏木各器官含碳量回歸模型①Tab. 4 Regression model of carbon storage of cypress organs
2.5 優良品系選擇
利用篩選出的含碳量最佳擬合模型,計算無性系和家系試驗林的單株總含碳量。由表5可知,含碳量的無性系重復力為0.647,家系遺傳力為0.161,按照20%入選率,無性系試驗林材積和含碳量的遺傳增益分別為39.74%和51.48%,家系試驗林分別為10.79%和16.14%。以單株材積和含碳量為指標,采用獨立淘汰法篩選出生長與固碳兼優無性系6個,分別為C6、C9、C12、C16、C13和C2,其單株材積平均值高出CK1的191.49%,單株含碳量平均值高出CK1的107.11%。篩選出生長與固碳兼優家系6個,分別為F18、F9、F24、F8、F25和F13, 其單株材積平均值高出CK2的32.45%,單株含碳量平均值高出CK2的33.39%(圖3)。
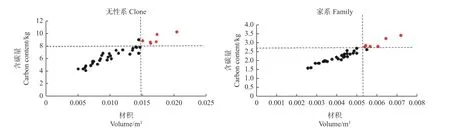
圖3 獨立淘汰法篩選生長和高固碳兼優的無性系與家系Fig. 3 Screening clones and families with high carbon fixation and growth by independent elimination method
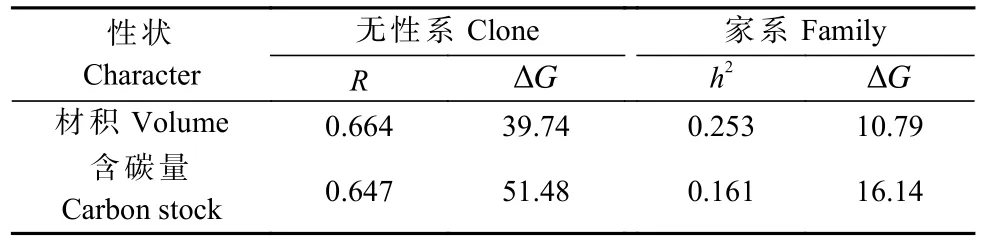
表5 20%入選率估算無性系與家系試驗林的單株材積和含碳量的遺傳增益①Tab. 5 Estimation of genetic gain of individual volume and carbon storage in clonal and family experimental forests with 20% selection rate
3 討論
已有研究表明,通過測定后選擇優良無性系造林,針葉樹可獲得額外5%~25%的遺傳增益,達到同世代家系試驗林所獲遺傳增益的2倍(Wu, 2019)。柏木無性系生長優勢明顯,樹高、胸徑和材積分別高出家系水平47.81%、28.39%和120.83%,按照20%入選率,估算無性系試驗林材積和含碳量的遺傳增益分別為39.74%和51.48%,家系試驗林分別為10.79%和16.14%,無性系試驗林相比于家系試驗林的遺傳增益高。Mullin 等(1994)對10年生黑云杉(Picea mariana)無性系測定表明,通過顯性效應和上位效應獲得的額外遺傳增益幾乎與全同胞和半同胞選擇獲得的遺傳增益相等。Wu 等(2008)對新西蘭輻射松的研究同樣表明,選擇前5%無性系,材積遺傳增益可達24%,在相同選擇強度下,比家系測定林提高1倍以上。在火炬松無性系試驗中,從每個家系選擇最佳單株繁育的無性系,前10名的遺傳增益幾乎是家系水平的2倍(Baltuniset al., 2007)。上述報道與本研究得出的結果類似,說明柏木無性系不僅更好地利用了加性遺傳方差,而且獲得了更多非加性遺傳方差,保持了優良性狀的遺傳值(Wenget al., 2008);但是,額外遺傳增益差異主要由群體大小、家系內測定的無性系數量、子代測定的準確性等因素造成,因此,為了更精準評價無性系和家系的增益水平,應在多個氣候區評估家系/無性系基因型與環境的互作效應,估計其加性和非加性遺傳方差,以預測無性系林業的穩定性和遺傳增益(Wu, 2019)。
本研究表明,生物量、含碳量與樹高和胸徑均存在極顯著線性關系,說明柏木生長快慢對生物量和含碳量具有顯著影響(郭金堂等,2016),而含碳率與樹高和胸徑不存在顯著線性關系,與張悅等(2022)對長白落葉松(Larix olgensis)各器官含碳量的測定結果類似,說明柏木生長快慢對含碳率影響較小,因此選擇生長性狀優良、生物量較大的優良品系,其固碳能力也相應較強。與家系不同,無性系的樹枝和樹葉分配比例均高于樹根,枝和葉的生物量占總生物量的41.67%,而家系的樹根分配比例高于樹枝,枝和葉的生物量占總生物量的38.96%,說明無性系具有較大的樹冠,無性系的冠幅比家系長33.33%(表1)。進一步分析發現,無性系和家系的地上與地下生物量之比分別為6.16∶1和5.22∶1,樹干與枝的生物量之比分別為2.40∶1和3.49∶1,樹干與葉的生物量之比分別為1.91∶1和1.73∶1,生物量分配策略表明,無性系是以犧牲地下粗根生物量為代價增強細根和枝葉的生長,無性系的枝具有更強的競爭能力,充足的枝和葉是無性系表現出快速生長的重要因素,生長快速的無性系優先分配更多的有機物給樹枝。Zhu等(2013)研究表明,在生理學上,生長快速的樹木吸收更多的陽光進行光合作用,并在地上積累更多有機物質。柏木喜鈣質土,生長在土壤瘠薄處,平衡生長假說認為,為滿足快速生長需求,需要更有效獲取環境中的資源,植株會將資源分配到最需要的器官中,這可能使柏木無性系的枝和細根器官分配更多的有機物(Stovallet al.,2013)。可以預期,與家系相比,無性系可以實現更高的生產力和碳儲存能力(Wu, 2019)。
本研究對無性系和家系分別擬合含碳量異速生長方程。胸徑作為最常用和最容易被精確測量的指標,在生物量和含碳量模型中應用最為廣泛,可獲得較高的擬合精度,但是本研究以胸徑為單一自變量時,模型R2均較低,這可能與樹齡有關,柏木試驗林僅9年生,胸徑較小,表型變異系數高;當以樹高和胸徑作為變量時,估算精度得到提高,有研究表明,生物量模型中加入樹高可抵消區域效應(Feldpauschet al.,2012);當將枝下高作為自變量包含在模型中則能顯著提高估算精度,同時不存在多重共線性問題,說明柏木樹高、胸徑和枝下高與含碳量存在極強的內在關聯,是含碳量模型的關鍵參數。柏木早期以高生長為主,分枝多,枝下高決定樹冠高度,直接影響植株的光合作用以及生長過程(李想等,2018;廖國莉等,2020),而樹冠特征也是影響枝葉生物量預測精度的重要變量(汪金松等,2011)。當前,優良林木基因型選擇是未來人工林生產和碳儲存能力關注的方向,從育種角度看,應用遺傳選擇可提高柏木生長潛力,表明栽植無性系可提高生產力。本研究按照20%入選率,評估無性系試驗林材積和含碳量的遺傳增益分別為39.74%和51.48%,家系試驗林分別為10.79%和16.14%,篩選出生長與固碳兼優的無性系和家系6個,為生產提供了重要材料。因此,在柏木早期生長階段,考慮胸徑、樹高、枝下高以及分枝等性狀來選擇優良品系,評估其固碳潛力更具有現實意義。
4 結論
柏木無性系生長優勢明顯,其樹高、胸徑和材積分別高出家系47.81%、28.39%和120.83%。柏木樹干生物量的分配比例最高,生物量分配模式差異是影響無性系和家系間生長差異的重要因素,無性系主要是以犧牲粗根的生物量分配(較家系低3.45%)、增加樹枝生物量(較家系高5.55%)和細根生物量(較家系高1.34%)來維持生長優勢。生物量、含碳量均與生長性狀呈極顯著線性關系,說明選擇速生、生物量較大的優良品系,可提高其固碳潛力,且無性系具有比家系更高的固碳能力。選擇將樹高、胸徑和枝下高作為特定參數,構建生長早期的含碳量模型精度高,預測效果好;進一步結合材積性狀篩選出速生與固碳兼優的無性系和家系各6個,將有助于提高柏木人工林的生產力和含碳量。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
小讀者(2021年2期)2021-03-29 05:03:48
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
光學精密工程(2016年6期)2016-11-07 09:07:19
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
