全基因組關聯分析篩選鵝蛋品質相關分子標記
2023-10-25 10:24:10高廣亮張克山趙獻芝許國洋謝友慧周莉張昌蓮王啟貴
中國農業科學 2023年19期
高廣亮,張克山,趙獻芝,許國洋,謝友慧,周莉,張昌蓮,王啟貴
全基因組關聯分析篩選鵝蛋品質相關分子標記
高廣亮1, 2,張克山1, 2,趙獻芝1, 2,許國洋1,謝友慧1, 2,周莉3,張昌蓮1, 2,王啟貴
1重慶市畜牧科學院,重慶 402460;2重慶市肉鵝遺傳改良工程技術研究中心,重慶 402460;3重慶綦江區動物疫病預防控制中心,重慶 401420
【目的】通過篩選與鵝蛋品質性狀相關的分子標記和候選基因,為解析蛋品質性狀的遺傳機制及分子標記輔助選擇提供理論支撐。【方法】采用同批次健康四川白鵝群體(209只)作為研究對象。收集了每只鵝在產蛋高峰期連續生產的5枚蛋,并測定了蛋重、蛋形指數、蛋殼強度、蛋殼厚度、蛋殼重和蛋黃重量等6個蛋品質性狀。基于前期209只四川白鵝(母鵝)2.896 Tb全基因組重測序數據(12.44×/個體),采用全基因組關聯分析的方法,篩選與蛋品質性狀相關的SNP位點和重要候選基因,并通過核酸飛行時間質譜方法檢測了這些SNP位點的基因型頻率。 【結果】經過篩選過濾,共有9 279 339個SNPs和209個個體用于后續研究。GWAS研究發現,48個SNP位點與6個鵝蛋品質性狀顯著或建議性顯著相關(閾值分別為5.43×10-9和1.09×10-7),并注釋出27個蛋品質性狀相關的候選基因,包括妊娠相關血漿蛋白A(pappalysin1, PAPPA)、蛋白絲氨酸/蘇氨酸磷酸酶調節亞基2基因(serine/threonine-protein phosphatase 4 regulatory subunit 2,PP4R2)、乙醇胺磷酸轉移酶1(ethanolamine phosphotransferase 1, EPT1)和離子型谷氨酸受體K2(glutamate receptor ionotropic, kainate 2,GRIK2)等,其中候選基因參與蛋白質代謝,促進生長因子IGF生成,在的11bp范圍內存在5個SNPs與蛋殼厚度顯著相關,另外在上有6個SNPs與蛋黃重量顯著相關,和分別與機體血鈣維持功能以及膽固醇代謝相關。功能富集研究發現,候選基因主要參與了response to growth factor(GO:0070848)、intracellular chemical homeostasis(GO:0055082)、response to hormone(GO:0009725)和regulation of the monoatomic ion transport(GO:43269)等代謝通路。【結論】經GWAS方法篩選出和分別作為蛋重、蛋黃重、蛋殼強度等蛋品質性狀潛在功能基因,為鵝蛋品質性狀的改良提供了分子遺傳標記的理論參考。
全基因組關聯分析;分子標記輔助選擇;鵝;蛋品質性狀
0 引言
【研究意義】我國是世界上養鵝量最大的國家,國家水禽產業體系2022年報告顯示我國鵝年出欄量已達4.68億只,產值526.73億元,因此針對直接影響鵝蛋經濟效益的蛋品質性狀研究具有重要的現實意義[1]。【前人研究進展】目前研究表明:品種、年齡、營養、環境、疾病等都可影響蛋品質性狀[2-7]。全基因組關聯分析(genome-wide association study, GWAS)方法是在全基因組水平上關聯定位分析重要經濟性狀的分子遺傳標記,篩選有生物學意義的候選基因。目前GWAS已廣泛運用到家禽的分子遺傳標記選擇中,如:ZHU等[8-9]對北京鴨體尺和屠體性狀等進行GWAS分析,發現高皮脂北京鴨抗脂肪生成基因存在特定的突變位點,并發現屠體重、全凈膛重等5個體組成性狀都有1個相同的全基因組顯著位點,候選基因為;Zhang等[10]在兩個高低腹脂專門化肉雞品系中用60K SNP芯片作單倍型GWAS研究,定位了7個可能控制腹脂含量的候選基因,包括、1、和等;LIU等采用600 K高密度SNP陣列,研究了產蛋后期72和80周齡的母雞蛋品質的全基因關聯分析,結果發現上的8.95—9.31 Mb(約0.36 Mb)的基因組區域與白蛋白高度和白蛋白單位密切相關,兩個最重要的SNPs占了3.12%—5.75%的表型變異,并篩選了3個與蛋殼顏色的相關的候選基因、和[11];前期本課題組通過GWAS分析篩選了與48周齡鵝蛋數、60周齡鵝蛋數候選基因以及與鵝蛋黃顏色顯著相關的、等4個基因[12]。【本研究切入點】由于鵝蛋品質性狀是一個復雜的多因素組成數量性狀,其蛋重、蛋形指數、蛋比重、蛋殼質量等性狀的遺傳力都較高,可通過遺傳分析來定位主效基因區間。【擬解決的關鍵問題】在本研究筆者通過GWAS挖掘鵝蛋品質性狀的相關分子標記,定位功能基因區間,為解析蛋品質性狀的遺傳機制及分子標記輔助選擇提供理論支撐。
1 材料與方法
1.1 實驗動物和表型測定
以重慶市家禽科研基地同批次健康四川白鵝(母鵝)群體(209只)為實驗動物。為準確記錄鵝蛋品質性狀,每只鵝從出雛后進行系統的腳號和翅號標記,鵝群28周齡時轉移到個體籠(600 mm×800 mm×900 mm)中飼養至65周(休產期),飼養過程自由飲水和采食全價飼料,收集產蛋高峰期每只鵝連續生產的5個鵝蛋,測定其蛋品質性狀(蛋重、蛋形指數、相對密度、蛋殼強度、蛋殼重、蛋殼厚度和蛋黃重)并統計其平均值作為后續GWAS研究的表型性狀。利用電子天平稱量蛋殼重、蛋黃重;利用游標卡尺測量蛋殼的鈍部、尖部和中間部位,并取上述三者的平均值計算蛋殼厚度;利用游標卡尺檢測蛋長直徑和短徑,計算蛋形指數;利用羅氏比色卡進行蛋黃顏色的測定;蛋殼強度利用蛋殼測力計(EFG-0502,Robotmation 公司)進行檢測。
1.2 DNA抽提及GWAS測序
抗凝真空管采集上述209只鵝翅靜脈血液2 mL,利用血液基因組提取試劑盒(北京天根公司,DP332)提取血液基因組DNA,NanoDrop 2000分光光度計質檢通過后,委托天津諾禾生物有限公司利用Illumina HiSeq X Ten平臺進行全基因重測序。數據過濾后,BWA軟件比對鵝基因組(ASM1303099v1)[13],GATK軟件進行SNP 基因型數據檢出[14];Plink軟件對獲得的SNP數據進行質量控制(參數設置:geno 0.1 mind 0.1 MAF 0.05 hwe 0.0000001)和主成分分析方法檢測群體結構分層情況[15]。
1.3 表型數據關聯與單倍型分析
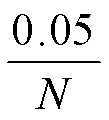

表1 驗證篩選的SNPs用引物序列表
1.4 SNPs注釋
BEDTools軟件[18]在鵝染色體基因組上提取鑒定的顯著SNPs位點上下游500 kb區域序列,Annovar軟件注釋SNP所在基因及鄰近基因[19],Metascape(http://metascape.org/)在線對候選基因進行功能分析[20]。
2 結果
2.1 蛋品質表型數據統計分析
209只四川白鵝個體高峰期平均蛋重為132.20 g;蛋形指數在1.24—1.60之間,平均蛋形指數為1.45;蛋殼強度平均值為67.20 kg·cm?2;蛋殼重平均為22.87 g;蛋殼厚度平均在0.42 cm;蛋黃重量平均值為41.20 g(表2)。

表2 統計分析鵝蛋品質性狀
2.2 測序數據分析
209只四川白鵝母鵝全基因組重測序共獲得2.896 Tb數據,測序平均覆蓋深度為12.44×。經數據過濾,2.891 Tb高質量測序數據比對到鵝參考基因組序列,比對率為96.58%—98.38%,共檢出了16 687 310個初始SNP位點;經過質量控制,最終檢出9 279 339個SNPs[12]。GWAS結果顯示,48個SNPs與上述6個蛋品質性狀顯著或建議性顯著關聯,并定位到27個基因(表3,圖1)。
以各性狀顯著關聯SNPs上下游500 k范圍內注釋基因,功能富集分析發現:候選基因主要聚類到response to growth factor(GO:0070848, Log(P)=-8.49)、intracellular chemical homeostasis(GO:0055082, Log(P)= -7.77)、response to hormone(GO:0009725, Log(P)=-7.44)和regulation of the monoatomic ion transport(GO: 43269, Log(P)= -6.08)。富集差異基因P值前20的GO分類條目如圖2所示。

表3 篩選與蛋品質性狀顯著相關的SNPs表

續表3 Continued table 3
: Pregnancy-associated plasma protein A;: Alpha-2C adrenergic receptor;: Epsilon-sarcoglycan;: Thioredoxin-like protein 1;: BCL-6 corepressor;: Mid1-interacting protein 1;: Glutaredoxin-3;: Pre-mRNA-splicing factor ATP-dependent RNA helicase DHX15;: TBC1 domain family member 14;: Dystrobrevin beta;: Ethanolamine phosphotransferase 1;: Nuclear factor of activated T-cells, cytoplasmic 1;: Cytoplasmic dynein 1 intermediate chain 1;: Protein bassoon;: NADPH oxidase 5;: KAT8 regulatory NSL complex subunit 1;: Uncharacterized protein KIAA1257;: Cytochrome P450 4V2;: Kinesin-like protein KIF16B;: Tribbles homolog 2;: Gamma-aminobutyric acid receptor subunit beta-3;: Uncharacterized protein KIAA1522;: Serine/threonine-protein phosphatase 4 regulatory subunit 2;: Activating signal cointegrator 1 complex subunit 3;: Glutamate receptor ionotropic, kainate 2;: Transcription elongation factor B polypeptide 1;:Transcription factor SOX-13
顯著關聯(閾值5.43×10-9),*潛在顯著關聯(閾值1.09×10-7)
Significant correlation (p-value 5.43×10-9), *potential significant correlation (p-value 1.09×10-7)

圖1 鵝蛋品質性狀全基因組關聯分析的曼哈頓圖和Q-Q plot圖
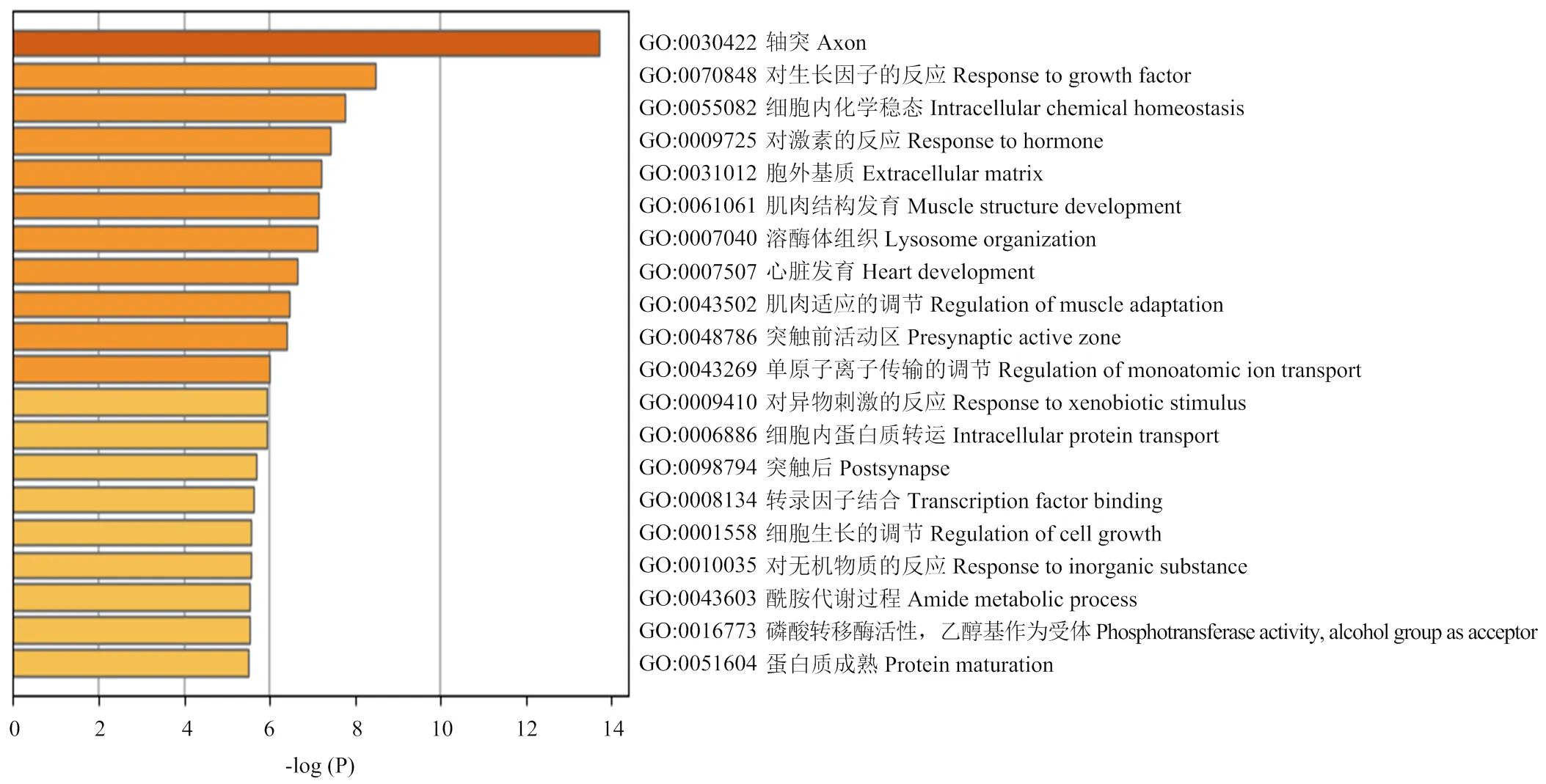
圖2 差異顯著SNPs上下游500k注釋基因聚類分析
2.3 顯著相關SNPs的MALDI-TOF MS驗證結果
針對重要候選SNPs合成合適的引物作MALDI- TOF MS驗證,結果如表4所示:對于蛋重性狀,chr29: 3653854 AA基因型個體極顯著(<0.01)高于AG和GG基因型個體;對于蛋形指數性狀,chr15: 20262147 GG基因型極顯著(<0.01)高于GT和TT基因型個體;對于蛋殼厚度性狀,chr18:18353514 CC基因型個體極顯著(<0.01)高于其他基因型個體;對于蛋黃重量性狀,chr34:3206857 GG基因型個體極顯著(<0.01)低于其他基因型個體;chr1:36829464 AA基因型個體極顯著(<0.01)高于其他基因型個體。
3 討論
3.1 鵝蛋品質性狀表型與其他家禽蛋品質的比較分析
蛋品質性狀是鵝蛋經濟效益的直接影響因素,蛋重決定了產蛋總重的大小,蛋形指數、蛋黃比例影響消費者對商品蛋的選擇,蛋重、蛋形指數、蛋殼質量等均會影響種蛋的合格率及孵化率[21-23]。本研究測定了四川白鵝產蛋高峰期蛋品質性狀,發現檢測群體的平均蛋重(132.20±18.90)g,顯著高于雞蛋和鴨蛋,但蛋形指數平均值為1.45,與雞、鴨蛋相當,正常雞蛋的蛋形指數一般為1.32—1.39,鴨蛋為1.20—1.58[24]。鵝蛋的平均蛋殼強度為(67.20±10.19)kg·cm?2,平均蛋殼厚度為(0.42±0.06)cm,明顯高于雞蛋,這可能解釋了生產中相同數量受精蛋雞蛋的孵化率可達90%以上,而鵝蛋孵化率一般在80%左右,更厚和強度更高的蛋殼增加了新生雛鵝的破殼難度而導致破殼失敗,降低了孵化率。鵝蛋的蛋黃平均重為(41.20±6.73)g,蛋黃比例達到30%以上,最大的蛋黃重量達到62.75 g。早成鳥的蛋黃比例一般比鴿子、企鵝等后代需要照顧的晚成鳥蛋黃比例高,鵝是早成鳥的一種,鵝蛋的高蛋黃比例可為幼雛供應更多的營養物質,使其出殼后能夠獨立生存[25-26]。
3.2 影響鵝蛋品質性狀表型的GWAS分析
蛋品質性狀是一個復雜的多因素控制數量性狀,遺傳因素是主要因素之一,已有的研究顯示:雞蛋的蛋重遺傳力約為0.5,殼顏色遺傳力為 0.45—0.75,蛋殼強度遺傳力約為0.3[21]。全基因組關聯技術能對數量性狀遺傳控制區間作有效定位。本研究通過GWAS在全基因組水平分析了鵝蛋品質遺傳控制因素,筆者對209只四川白鵝血樣DNA進行全基因組重測序,基于前期本課題組組裝了一個染色體水平的鵝基因組[24],有利于將高質量的全基因組重測序數據比對到鵝基因組序列并進行了GWAS分析。最終篩選并使用MALDI-TOF MS驗證了48個與蛋重、蛋殼厚度、蛋殼強度等蛋品質性狀顯著相關的SNPs。

表4 部分SNPs的基因型對鵝6個蛋品質性狀的影響
不同字母代表差異顯著(<0.05);相同字母代表差異不顯著(>0.05)
Different letters in the same row indicate a significant difference between the genotypes (<0.05); the same letter indicates no significant difference between genotypes (>0.05)
通過GWAS分析在12、15以及29號染色體上共鑒定4個SNPs與蛋重顯著相關,分別位于、、和。妊娠相關血漿蛋白A(pregnancy-associated plasma protein A,)基因編碼一種分泌型金屬蛋白酶,可裂解胰島素樣生長因子結合蛋白(),而激活IGF信號通路,參與蛋白質代謝以及對胰島素樣生長因子(IGF)運輸和攝取的調節過程[27]。在卵泡發育過程中卵丘顆粒細胞中mRNA表達水平上調是卵母細胞成熟的標志之一[28]。MAROULI等[29]通過全基因組關聯研究發現了83個與人身高相關的編碼變體,其中增高等位基因促進了的蛋白酶活性,增加了對的裂解,從而導致的生物利用度提高。在家禽中,體型和蛋重一般有著正向相關的關系,能夠促進身高,這提示可能作為蛋重正向選擇的候選基因。值得注意的是,在篩選的48個候選SNPs中有5個與蛋殼厚度相關的SNPs都定位在chr18的蛋白絲氨酸/蘇氨酸磷酸酶調節亞基2基因(serine/ threonine-protein phosphatase 4 regulatory subunit 2,),是PP4復合體5個調節亞基中的一個,參與許多關鍵的細胞途徑,包括DNA損傷反應(DNA修復、細胞周期調節和細胞凋亡)、葡萄糖代謝、細胞遷移、腫瘤發生和免疫反應等[30-31]。家禽無論在排卵還是在蛋殼形成過程中都存在著明確的時序性,這種時序性受周期調節基因的控制,而調控核心節律基因/活性[32],另外,高濃度的葡萄糖影響Ca2+吸收[33],而參與葡萄糖代謝,由此推測可能參與了蛋殼形成。通過GWAS分析發現chr1:1466542和chr1: 1467878與蛋殼強度顯著相關的SNPs都定位乙醇胺磷酸轉移酶1(ethanolamine phosphotransferase 1, EPT1)基因上,該基因編碼蛋白將CDP-乙醇胺催化形成磷脂酰乙醇胺,是磷脂代謝的重要組成部分[34]。如表3所示,6個SNPs定位在chr1上的(glutamate receptor ionotropic, kainate 2)與蛋黃重量顯著相關(P=3.46*10-8—2.87* 10-9),ZEMUNIK等[35]通過全基因組關聯分析方法在一群孤立的人群中鑒別了與生化性狀(總膽固醇、低密度脂蛋白膽固醇、高密度脂蛋白膽固醇和甘油三酯等)相關的遺傳變異基礎,發現、和等是重要的候選基因。另外,兩個定位到(activating signal cointegrator 1 complex subunit 3, ASSC3)上的SNPs chr1 rs36803702C/T和rs36829464 A/G 同樣與蛋黃重量顯著相關,前期研究發現蛋白酶前體轉化酶亞硫酸酯酶/切割酶9型(Proprotein convertase subtilisin/kexin type 9,PCSK9)是最主要的低密度脂蛋白膽固醇調節子,可減少低密度脂蛋白膽固醇的含量,而和能作用核糖體,保護細胞免受因PCSK9抑制劑(PF8503)引起的細胞毒性[36]。蛋黃主要由膽固醇和磷脂組成,本研究發現的候選基因、等均參與膽固醇代謝,由此推測、可作為蛋黃重量相關的候選基因。
3.3 候選基因的功能聚類分析
家禽的產蛋行為受激素調控,FSH促進卵泡生長成熟,LH促進排卵, PRL與抱窩性相關,性激素維持鵝的生殖特征,甲狀旁腺激素和降鈣素調控蛋殼中鈣的沉積[37-39]。在本研究中筆者通過對注釋的功能基因進行聚類分析也發現response to hormone(GO: 0009725)是P值最高的GO分類條目之一。每個雞蛋中的鈣量通常占雞體內鈣儲存總量的10%左右,身體中絕大多數鈣存在于細胞內,僅有不足0.1%位于細胞外,這部分包括電離鈣、與蛋白質結合的鈣以及與陰離子結合的鈣三種形式,其中電離鈣是生理活性形式,家禽能在24 h內將身體10%的鈣形成蛋殼[40- 41],而不引起機體出現低血鈣癥,這與家禽獨特的血清鈣平衡維持密切相關,在進行GWAS分析的時候筆者也發現與蛋品質顯著相關的候選基因明顯聚到intracellular chemical homeostasis (GO:0055082)、regulation of the monoatomic ion transport(GO:0043269)等通路。
4 結論
本研究通過GWAS方法在全基因組水平上篩選與蛋品質性狀顯著相關的分子遺傳標記,注釋篩選了重要候選基因、、和分別作為蛋重、蛋黃重、蛋殼強度等蛋品質性狀潛在功能基因。研究結果促進了鵝蛋品質性狀分子標記選擇研究與運用,也將為四川白鵝蛋品質性能的選育提供理論支撐。
[1] 侯水生, 劉靈芝. 2022年水禽產業現狀、未來發展趨勢與建議. 中國畜牧雜志, 2023, 59(3): 274-280.
HOU S S, LIU L Z. Present situation, future development trend and suggestions of waterfowl industry in 2022. Chinese Journal of Animal Science, 2023, 59(3): 274-280. (in Chinese)
[2] 王一冰, 陳芳, 茍鐘勇, 李龍, 林廈菁, 張盛, 蔣守群. 快大型黃羽肉種雞VD3需要量研究. 中國農業科學, 2021, 54(16): 3549-3560.
WANG Y B, CHEN F, GOU Z Y, LI L, LIN X J, ZHANG S, JIANG S Q. Requirement of vitamin D3on fast-growing yellow-feathered breeder hens. Scientia Agricultura Sinica, 2021, 54(16): 3549-3560. (in Chinese)
[3] KOWALSKA E, KUCHARSKA-GACA J, KU?NIACKA J, LEWKO L, GORNOWICZ E, BIESEK J, ADAMSKI M. Egg quality depending on the diet with different sources of protein and age of the hens. Scientific Reports, 2021, 11: 2638.
[4] NASRI H, VAN DEN BRAND H, NAJJAR T, BOUZOUAIA M. Egg storage and breeder age impact on egg quality and embryo development. Journal of Animal Physiology and Animal Nutrition, 2020, 104(1): 257-268.
[5] SUN C J, LIU J N, YANG N, XU G Y. Egg quality and egg albumen property of domestic chicken, duck, goose, Turkey, quail, and pigeon. Poultry Science, 2019, 98(10): 4516-4521.
[6] DA SILVA TEIXEIRA M, TRIGINELLI M V, DE ATAíDE COSTA T, LARA L J C, SOTO-BLANCO B. Effects of caffeine on egg quality and performance of laying hens. Frontiers in Veterinary Science, 2020, 7: 545359.
[7] WANG J, YUE H Y, WU S G, ZHANG H J, QI G H. Nutritional modulation of health, egg quality and environmental pollution of the layers. Animal Nutrition, 2017, 3(2): 91-96.
[8] ZHU F, CUI Q Q, HOU Z C. SNP discovery and genotyping using Genotyping-by-Sequencing in Pekin ducks. Scientific Reports, 2016, 6: 36223.
[9] ZHU F, YIN Z T, WANG Z, SMITH J, ZHANG F, MARTIN F, OGEH D, HINCKE M, LIN F B, BURT D W, ZHOU Z K, HOU S S, ZHAO Q S, LI X Q, DING S R, LI G S, YANG F X, HAO J P, ZHANG Z D, LU L Z, YANG N, HOU Z C. Three chromosome-level duck genome assemblies provide insights into genomic variation during domestication. Nature Communications, 2021, 12: 5932.
[10] ZHANG H, SHEN L Y, XU Z C, KRAMER L M, YU J Q, ZHANG X Y, NA W, YANG L L, CAO Z P, LUAN P, REECY J M, LI H. Haplotype-based genome-wide association studies for carcass and growth traits in chicken. Poultry Science, 2020, 99(5): 2349-2361.
[11] LIU Z, SUN C J, YAN Y Y, LI G Q, SHI F Y, WU G Q, LIU A Q, YANG N. Genetic variations for egg quality of chickens at late laying period revealed by genome-wide association study. Scientific Reports, 2018, 8: 10832.
[12] GAO G L, GAO D F, ZHAO X Z, XU S S, ZHANG K S, WU R, YIN C H, LI J, XIE Y H, HU S L, WANG Q G. Genome-wide association study-based identification of SNPs and haplotypes associated with goose reproductive performance and egg quality. Frontiers in Genetics, 2021, 12: 602583.
[13] LI H, DURBIN R. Fast and accurate long-read alignment with Burrows-Wheeler transform. Bioinformatics, 2010, 26(5): 589-595.
[14] MCKENNA A, HANNA M, BANKS E, SIVACHENKO A, CIBULSKIS K, KERNYTSKY A, GARIMELLA K, ALTSHULER D, GABRIEL S, DALY M, DEPRISTO M A. The Genome Analysis Toolkit: a MapReduce framework for analyzing next-generation DNA sequencing data. Genome Research, 2010, 20(9): 1297-1303.
[15] PURCELL S, NEALE B, TODD-BROWN K, THOMAS L, FERREIRA M A R, BENDER D, MALLER J, SKLAR P, DE BAKKER P I W, DALY M J, SHAM P C. PLINK: a tool set for whole-genome association and population-based linkage analyses. The American Journal of Human Genetics, 2007, 81(3): 559-575.
[16] ZHOU X, STEPHENS M. Genome-wide efficient mixed-model analysis for association studies. Nature Genetics, 2012, 44(7): 821-824.
[17] KARSSEN L C, VAN DUIJN C M, AULCHENKO Y S. The GenABEL project for statistical genomics. F1000Research, 2016, 5: 914.
[18] QUINLAN A R. BEDTools: the swiss-army tool for genome feature analysis. Current Protocols in Bioinformatics, 2014, 47: 11.12.1- 11.1234.
[19] WANG K, LI M Y, HAKONARSON H. ANNOVAR: functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Research, 2010, 38(16): e164.
[20] ZHOU Y Y, ZHOU B, PACHE L, CHANG M, KHODABAKHSHI A H, TANASEICHUK O, BENNER C, CHANDA S K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nature Communications, 2019, 10: 1523.
[21] KETTA M, T?MOVá E. Eggshell structure, measurements, and quality-affecting factors in laying hens: a review. Czech Journal of Animal Science, 2016, 61(7): 299-309.
[22] HISASAGA C, GRIFFIN S E, TARRANT K J. Survey of egg quality in commercially available table eggs. Poultry Science, 2020, 99(12): 7202-7206.
[23] PICARD DRUET D, VARENNE A, HERRY F, HéRAULT F, ALLAIS S, BURLOT T, LE ROY P. Reliability of genomic evaluation for egg quality traits in layers. BMC Genetics, 2020, 21(1): 17.
[24] LI Y, GAO G L, LIN Y, HU S L, LUO Y, WANG G S, JIN L, WANG Q G, WANG J W, TANG Q Z, LI M Z. Pacific Biosciences assembly with Hi-C mapping generates an improved, chromosome- level goose genome. GigaScience, 2020, 9(10): 114.
[25] LIKER A, SZéKELY T. Mortality costs of sexual selection and parental care in natural populations of birds. International Journal of Organic Evolution, 2005, 59(4): 890-897.
[26] WILLIAMS T D. Physiology, activity and costs of parental care in birds. The Journal of Experimental Biology, 2018, 221(Pt 17): jeb169433.
[27] GYRUP C, OXVIG C. Quantitative analysis of insulin-like growth factor-modulated proteolysis of insulin-like growth factor binding protein-4 and -5 by pregnancy-associated plasma protein-A. Biochemistry, 2007, 46(7): 1972-1980.
[28] KORDUS R J, HOSSAIN A, CORSO M C, CHAKRABORTY H, WHITMAN-ELIA G F, LAVOIE H A. Cumulus cell pappalysin-1, luteinizing hormone/choriogonadotropin receptor, amphiregulin and hydroxy-delta-5-steroid dehydrogenase, 3 beta- and steroid delta- isomerase 1 mRNA levels associate with oocyte developmental competence and embryo outcomes. Journal of Assisted Reproduction and Genetics, 2019, 36(7): 1457-1469.
[29] MAROULI E, GRAFF M, MEDINA-GOMEZ C, LO K S, WOOD A R, KJAER T R, FINE R S, LU Y C, SCHURMANN C, HIGHLAND H M, RüEGER S, et al. Rare and low-frequency coding variants alter human adult height. Nature, 2017, 542(7640): 186-190.
[30] LEE J, ADELMANT G, MARTO J A, LEE D H. Dephosphorylation of DBC1 by protein phosphatase 4 is important for p53-mediated cellular functions. Molecules and Cells, 2015, 38(8): 697-704.
[31] PARK J, LEE D H. Functional roles of protein phosphatase 4 in multiple aspects of cellular physiology: a friend and a foe. BMB Reports, 2020, 53(4): 181-190.
[32] KLEMZ S, WALLACH T, KORGE S, ROSING M, KLEMZ R, MAIER B, FIORENZA N C, KAYMAK I, FRITZSCHE A K, HERZOG E D, STANEWSKY R, KRAMER A. Protein phosphatase 4 controls circadian clock dynamics by modulating CLOCK/BMAL1 activity. Genes & Development, 2021, 35(15/16): 1161-1174.
[33] POHOREC V, KRI?AN?I? BOMBEK L, SKELIN KLEMEN M, DOLEN?EK J, STO?ER A. Glucose-stimulated calcium dynamics in beta cells from male C57BL/6J, C57BL/6N, and NMRI mice: a comparison of activation, activity, and deactivation properties in tissue slices. Frontiers in Endocrinology, 2022, 13: 867663.
[34] AHMED M Y, AL-KHAYAT A, AL-MURSHEDI F, AL-FUTAISI A, CHIOZA B A, PEDRO FERNANDEZ-MURRAY J, SELF J E, SALTER C G, HARLALKA G V, RAWLINS L E, et al. A mutation ofunderlies a new disorder of Kennedy pathway phospholipid biosynthesis. Brain, 2017: aww318.
[35] ZEMUNIK T, BOBAN M, LAUC G, JANKOVI? S, ROTIM K, VATAVUK Z, BENCI? G, DOGAS Z, BORASKA V, TORLAK V, SUSAC J, ZOBI? I, RUDAN D A, PULANI? D, MODUN D, MUDNI? I, GUNJACA G, BUDIMIR D, HAYWARD C, VITART V, WRIGHT A F, CAMPBELL H, RUDAN I. Genome-wide association study of biochemical traits in Korcula Island, Croatia. Croatian Medical Journal, 2009, 50(1): 23-33.
[36] LIAUD N, HORLBECK M A, GILBERT L A, GJONI K, WEISSMAN J S, CATE J H D. Cellular response to small molecules that selectively stall protein synthesis by the ribosome. PLoS Genetics, 2019, 15(3): e1008057.
[37] CHEN Q Y, DUAN J D, WU H Z, LI J B, JIANG Y L, TANG H, LI X Y, KANG L. Expression dynamics of gonadotropin-releasing hormone-I and its mutual regulation with luteinizing hormone in chicken ovary and follicles. General and Comparative Endocrinology, 2019, 270: 96-102.
[38] GHANEM K, JOHNSON A L. Follicle dynamics and granulosa cell differentiation in the Turkey hen ovary. Poultry Science, 2018, 97(10): 3755-3761.
[39] TANIMOTO R, SEKII K, MOROHAKU K, LI J Z, PéPIN D, OBATA Y. Blocking estrogen-induced AMH expression is crucial for normal follicle formation. Development, 2021, 148(6): dev197459.
[40] DE MATOS R. Calcium metabolism in birds. Veterinary Clinics of North America: Exotic Animal Practice, 2008, 11(1): 59-82.
[41] DESCALZO E, CAMARERO P R, SáNCHEZ-BARBUDO I S, MARTINEZ-HARO M, ORTIZ-SANTALIESTRA M E, MORENO- OPO R, MATEO R. Integrating active and passive monitoring to assess sublethal effects and mortality from lead poisoning in birds of prey. The Science of the Total Environment, 2021, 750: 142260.
Identification of Molecular Markers Associated with Goose Egg Quality Through Genome-Wide Association Analysis

1Chongqing Academy of Animal Science, Chongqing 402460;2Chongqing Engineering Research Center of Goose Genetic Improvement, Chongqing 402460;3Chongqing Qijiang Animal Disease Control Center, Chongqing 401420
【Objective】 The objective of this study was to screen molecular markers and candidate genes related to goose egg quality traits, to provide a theoretical support for the analysis of the genetic mechanism of egg quality traits and marker-assisted selection. 【Method】 In this study, a batch of healthy Sichuan White Geese (209 individuals) was selected as the research subjects. Five eggs from each goose during the peak egg production period were collected, and then six egg quality traits were measured, including egg weight, egg shape index, eggshell strength, eggshell thickness, eggshell weight, and egg yolk weight. Based on the whole-genome resequencing data (2.896 Tb, 12.44×/individual) of 209 Sichuan White Geese (female geese), a genome-wide association analysis was conducted to identify SNP loci and important candidate genes associated with egg quality traits. The genotype frequencies of the SNP loci were determined using the nucleic acid flight time mass spectrometry method. 【Result】After filtering, a total of 9 279 339 SNPs and 209 individuals were included for further analysis. The GWAS analysis identified 48 SNP loci significantly or suggestively associated with six egg quality traits (thresholds: 5.43×10-9and 1.09×10-7). These loci were annotated to 27 candidate genes related to egg quality traits, including Pregnancy-associated plasma protein A (), Serine/threonine-protein phosphatase 4 regulatory subunit 2 (), Ethanolamine phosphotransferase 1 (), and Glutamate receptor ionotropic, kainate 2 (). Among them, the candidate genewas involved in protein metabolism and promotes the generation of insulin-like growth factor. Five SNPs within the 11 bp range ofwere significantly associated with eggshell thickness. Additionally, six SNPs on thegene were significantly associated with yolk weight.andwere respectively associated with blood calcium homeostasis and cholesterol metabolism in organisms. Functional enrichment analysis revealed that the candidate genes were mainly annotated to “response to growth factor” (GO:0070848), “intracellular chemical homeostasis” (GO:0055082), “response to hormone” (GO:0009725), and “regulation of monoatomic ion transport” (GO:43269). 【Conclusion】 The GWAS analysis showed that theandare potential functional genes associated with various egg quality traits, such as egg weight, egg yolk weight, and shell strength, providing theoretical references for molecular genetic marker-assisted selection of goose egg quality traits.
genome-wide association analysis; molecular marker-assisted selection; goose; egg quality traits
10.3864/j.issn.0578-1752.2023.19.015
2022-05-10;
2023-08-08
重慶市科研機構績效激勵引導專項項目(cstc2022jxjl80007)、重慶市科研院所績效激勵引導專項項目(22527J)、財政部和農業部:國家現代農業產業技術體系(CARS-42-51)、重慶市技術創新與應用發展專項重點項目(cstc2019jscx-gksbX0097)
高廣亮,E-mail:guanglianggaocq@hotmail.com。張克山,E-mail:zhangkshLK1988@163.com。高廣亮和張克山為同等貢獻作者。通信00作者王啟貴,E-mail:wangqigui@hotmail.com
(責任編輯 林鑒非)
