不同水位條件下鄱陽湖越冬白鶴的移動模式與棲息地利用*
2023-10-19 07:06:14江紅星陳麗霞王藝璇
林業科學 2023年8期
王 瑩 江紅星 陳麗霞 王藝璇 高 彤
(中國林業科學研究院森林生態環境與自然保護研究所 生物多樣性保護國家林業和草原局重點實驗室 北京 100091)
白鶴(Leucogeranus leucogeranus)是世界上15 種鶴類中特定依賴濕地生存的3 種鶴類之一(Meineet al.,1996),是我國I 級重點保護野生動物,世界自然保護聯盟紅皮書將其列為“極危”等級。白鶴全球種群分為西部、中部和東部3 個種群,它們均在俄羅斯北部繁殖,遷徙數千公里,分別在伊朗、印度和中國越冬。目前,白鶴全球種群數量估計為3 500~4 000 只,幾乎全部為東部種群(http://www.birdlife.org)。白鶴東部種群繁殖于俄羅斯西伯利亞東北部的苔原地帶,遷徙途徑我國松嫩平原、遼河平原和華北平原,98%的種群在江西鄱陽湖越冬(Kanaiet al.,2002;馬克·巴特等,2005;Houet al.,2020)。因此,鄱陽湖濕地生態環境的穩定性對于全球白鶴種群的穩定與發展至關重要。
近些年來,鄱陽湖水位的年內和年際間變化,導致白鶴在鄱陽湖的空間分布格局和棲息地利用模式發生了顯著變化(Houet al.,2020)。歷史上,鄱陽湖越冬白鶴主要以湖區淺水沼澤和泥灘中的沉水植物苦草(Vallisneriaspp.)塊莖為主要食物(曾南京等,2002;孫志勇等,2010;吳建東等,2013;Burnhamdet al.,2017)。鄱陽湖4—10 月為沉水植物苦草的生長期,其中7—10 月為冬芽形成期(萬文豪等,2002),水位過深或過淺都會抑制苦草光合作用,影響苦草生長和冬芽形成(Dobberfuhl,2007;Wanget al.,2008),進而影響越冬期白鶴覓食棲息地的選擇(Wuet al.,2009;黎磊等,2015)。
2016 年越冬期以來,在鄱陽湖水位變化加劇,越冬白鶴對周邊農田棲息地的利用率顯著增加,特別是藕塘和稻田(王文娟等,2019;Houet al.,2020),但有關越冬白鶴的移動模式和棲息地利用尚缺乏定量對比研究。本文基于2016—2022 年13 只衛星跟蹤白鶴在鄱陽湖越冬期的定位數據,利用 ArcGIS 10.7 空間分析功能和鄱陽湖區土地利用類型數據,對比分析豐水年和平水年越冬白鶴的移動距離、活動區面積大小、棲息地利用情況,以便為鄱陽湖越冬白鶴種群保護和棲息地適應性管理提供科學依據。
1 研究區概況
鄱陽湖(115°49′—116°46′E,28°11′—29°51′N)位于江西省北部和長江以南,是中國最大的淡水湖。鄱陽湖是與長江連通的典型季節性吞吐湖泊,受亞熱帶季風性氣候、長江和鄱陽湖五河注入的影響,水位年變化幅度大。每年4—9 月為豐水期,10 月—翌年3 月為枯水期,豐水和枯水季節湖面水位變化最大可達9~14 m,有“高水是湖,低水似河”的獨特景觀(張本,1988)。在枯水季節隨著鄱陽湖水位下降,鄱陽湖周邊不同高程的淺水洼地相繼與主湖分開,形成子湖泊。這些子湖泊地勢平緩,多呈淺碟型,其生境呈“草灘—泥灘—淺水水域”的環帶狀分布,這種復雜多樣的濕地生境為不同生態環境需求的越冬水鳥提供了重要的食物資源和棲息場所(馬克·巴特等,2005;金斌松等,2016)。
2 研究方法
2.1 白鶴衛星跟蹤
2016—2020 年,在我國東北白鶴中途停歇地對救助恢復后的13 只白鶴佩戴了背負式衛星跟蹤器。跟蹤器由湖南環球信士科技有限公司生產,型號為HQBG3621S 和HQBG5037S,質量分別為24 g 和40 g,符合跟蹤器質量不超過研究對象體質量3%~5%的國際標準(Higuchiet al.,1996;1998)。本研究僅對定位精度包括A、B、C 3 個等級的數據進行分析,定位誤差分別為5、10、20 m。
2.2 數據分割
以鄱陽湖的北端29.51°N 為標志,確定13 只白鶴跟蹤個體每年遷到與遷離鄱陽湖的日期,并統計不同年份的越冬天數與有效位點數(表1)。
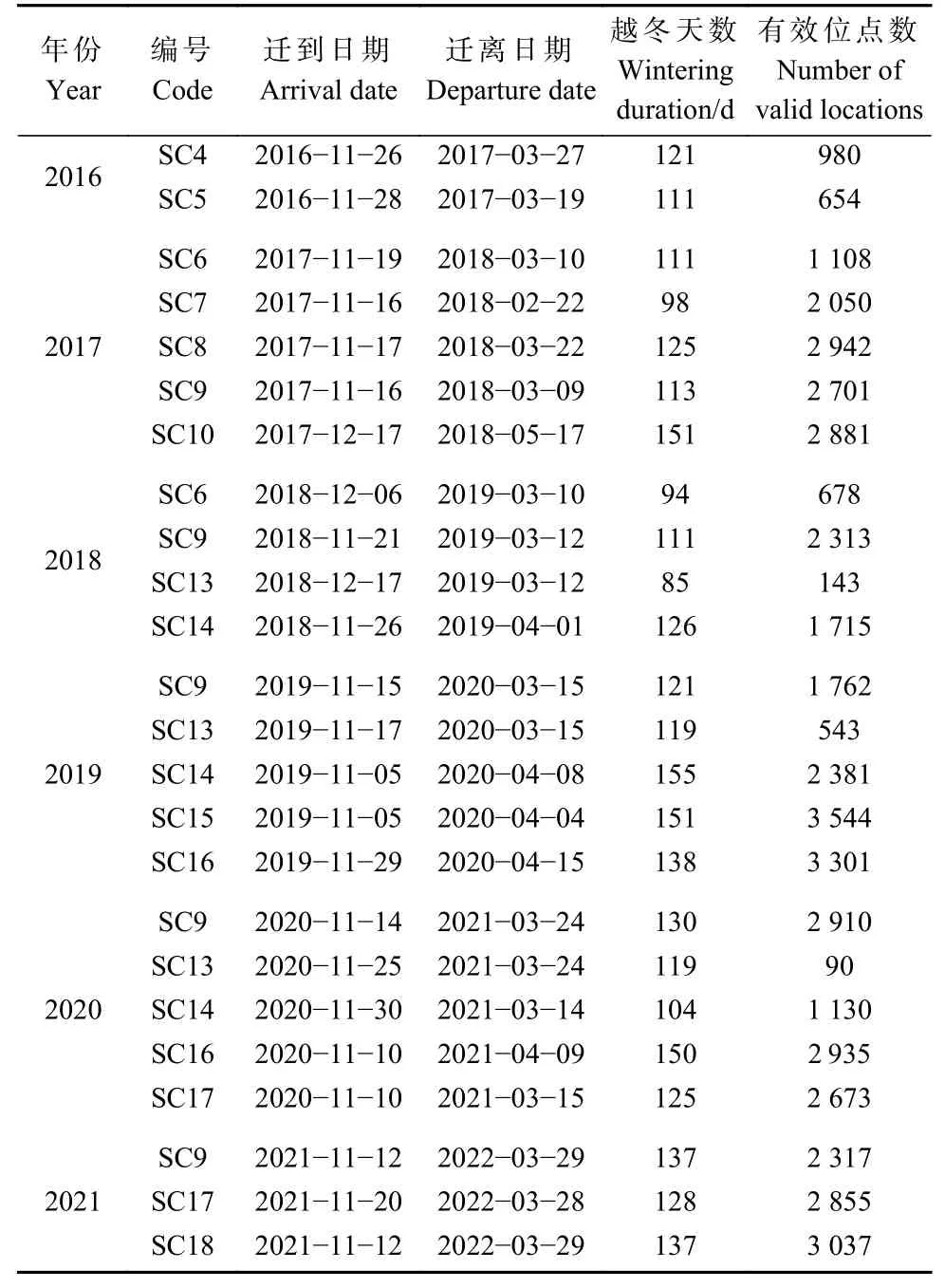
表1 2016—2021 年越冬期鄱陽湖白鶴衛星跟蹤信息Tab. 1 Satellite tracking information of Siberian Cranes in winters from 2016 to 2021 in Poyang Lake
不同水位年組界定,主要依據鄱陽湖水文站水位數據以及湖區白鶴自然食物苦草塊莖生物量。自2003 年以來,三峽大壩的運行對鄱陽湖水位變化產生了重要影響(圖1a)。對比2003—2022 年間鄱陽湖星子站4—10 月平均水位與2016—2021 年度4—10 月平均水位,2016、2017、2020 和2021 年度4—10 月平均水位均高于2003—2022 年間平均水位,而2018 和2019 年度4—10 月平均水位均低于2003—2022 年間平均水位(圖1b)。結合郝能祖等(2019)和余定坤等(2020)監測結果,2018 年和2019 年湖區的苦草密度均高于其他4 個年份。鑒于此,本研究將6 個研究年份劃分為3 個水位年組:2016 年和2017 年越冬期為W1 豐水年組、2018 年和2019 年越冬期為W2 平水年組、2020 年和2021 年越冬期為W3 豐水年組。豐水年組內和組間的4—10 月水位差異不顯著,平水年組內的4—10 月水位差異不顯著。
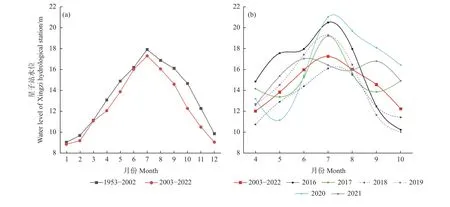
圖1 星子水文站月平均水位Fig. 1 The monthly average water levels of Xingzi Hydrological Station
根據歷年白鶴的越冬習性,將白鶴遷到鄱陽湖至12 月中旬定義為越冬前期;12 月下旬至翌年2 月中旬定義為越冬中期,白鶴種群數量與分布相對穩定;翌年2 月下旬至遷離鄱陽湖定義為越冬后期(夏少霞等,2010;胡振鵬等,2012)。根據楊秀林(2019)通過計算白鶴每天每小時移動距離,將6:00—17:59 期間的定位數據界定為白鶴的日間活動位點,將18:00—翌日5:59 期間的定位數據界定為白鶴的夜間活動位點。
2.3 移動距離與活動區大小
不同越冬階段的白鶴移動距離為相應越冬階段連續2 個位點間歐式距離之和。越冬期白鶴移動總距離為整個越冬期連續2 個位點間的歐式距離之和。
活動區(home range)面積在一定程度能反映棲息地質量,目前多使用核密度估計法(Kernel Density Estimator,KDE)(Worton,1995;Powell,2000)。為剔除白鶴處于飛行狀態的位點,本研究篩選出速度小于3 km·h-1的位點用于活動區面積計算。使用R 4.2.0“adehabitatHR”包生成每只白鶴個體不同越冬階段的90%KDE 和50%KDE 范圍,其中90%KDE 代表活動區大小,50%KDE 代表核域大小,將生成的shp 文件導入ArcGIS 10.7 分別計算90%KDE 和50%KDE 面積。
2.4 土地利用類型分類
從中國科學院資源環境科學數據中心(http://www.resdc.cn/)下載研究區2015、2018 和2020 年的土地利用類型數據。該數據包括6 個一級地類和25 個二級地類。考慮到白鶴主要利用稻田、藕塘、自然濕地和草地生境(Jiaet al.,2013;Burnhamdet al.,2017;王文娟等,2019;Houet al.,2020),使用ArcGIS 10.7 的重分類功能,提取研究區的農田(水田、旱地)、濕地(湖泊、沼澤地、灘地)、草地和其他類型(林地、城鄉、工礦、居民用地和未利用土地)等4 種土地利用類型。利用Spatial Analyst 工具中的多值提取至點功能,提取跟蹤白鶴個體日間和夜間衛星跟蹤位點所對應的土地類型。不同越冬階段各土地類型的利用率為對應階段該土地類型位點數除以總位點數(Mintonet al.,2003;王昱熙等,2020)。
2.5 統計分析
所獲得的數據從兩方面進行統計分析:1)3 個水位年組(W1、W2、W3)之間的對比分析;2)豐水年組(W1 和W3 合并)與平水年組(W2)的對比分析。
首先進行獨立樣本Kolmogorov-Smirnov Test 檢驗數據是否符合正態性。若符合正態分布,采用單因素方差分析(One-way ANOVA)分析和2 個獨立樣本t檢驗。若不符合正態分布,采用非參數檢驗方法中的多個獨立樣本Kruskal-Wallis H 檢驗和2 個獨立樣本Mann-Whitney U 檢驗。統計數據均表示為算術平均值±標準差(Mean ± SD),統計分析在SPSS 22.0 中進行。
3 結果與分析
3.1 越冬期不同階段的移動距離
越冬期白鶴移動總距離在豐水年W1 組、W3 組以及合并組(W1+W3)分別為[(1 368 ± 446) km,n=7]、[(1 544 ± 240) km,n=6]和[(1 398 ± 398) km,n=13],平水年W2 組為[(760 ± 420) km,n=9]。其中,豐水年3 組的白鶴移動總距離顯著大于平水年W2 組(W1/W2 組:P= 0.006;W3/W2 組:P= 0.001;合并組/W2 組:P=0.001);豐水年組之間的白鶴移動總距離無顯著差異(P> 0.05)。
從越冬期不同階段的白鶴移動距離來看,越冬前期豐水年合并組的白鶴移動距離顯著大于平水年W2組(合并組/W2 組:P= 0.034),其他組間無顯著差異;越冬中期,豐水年3 組的白鶴移動距離顯著大于平水年W2 組(W1/W2 組:P= 0.000;W3/W2 組:P= 0.001;合并組/W2 組:P= 0.000);越冬后期,各組間無顯著差異(P> 0.05)(圖2)。
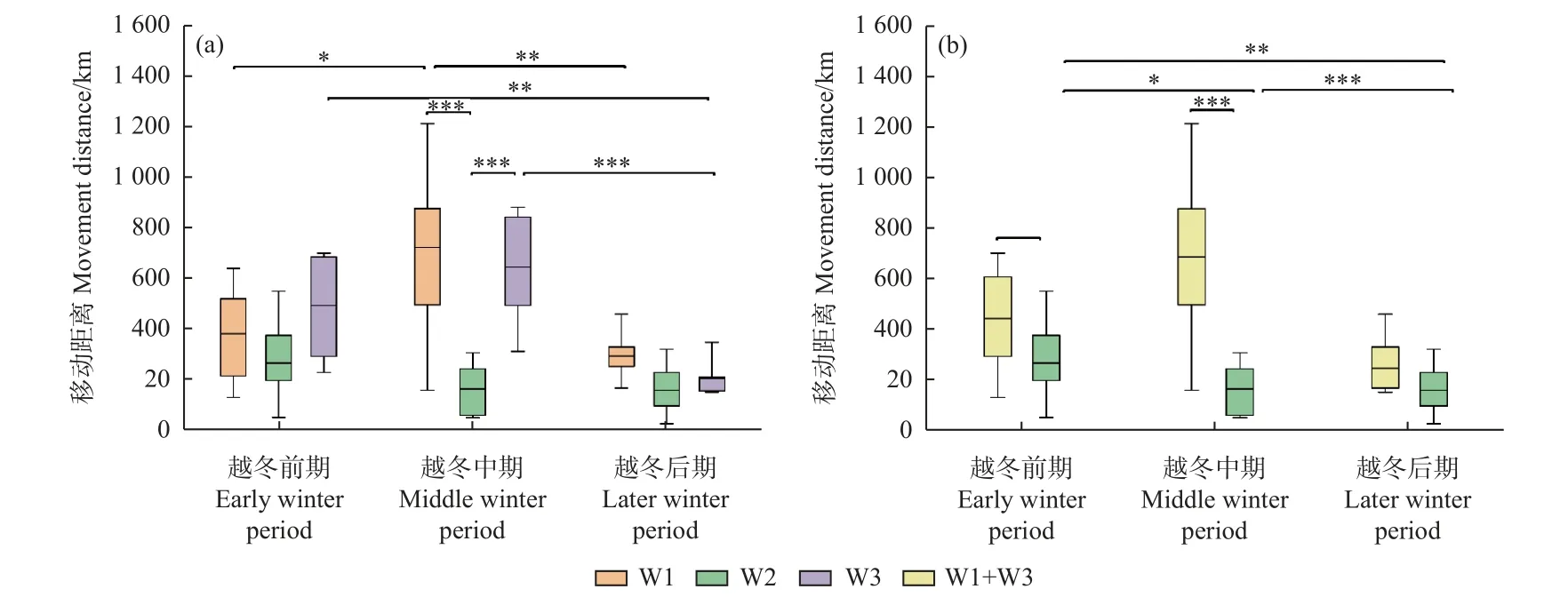
圖2 越冬期不同階段的白鶴移動距離Fig. 2 Movement distances of Siberian Cranes in different winter stages
從豐水年和平水年越冬期不同階段的移動距離可知,豐水年3 組白鶴移動距離為越冬中期 > 前期 >后期,除W1 組越冬前期和后期間的移動距離及W3組越冬前期和中期間的移動距離差異不顯著外,其余均差異顯著(P< 0.05)。平水年W2 組不同越冬階段的白鶴移動距離均差異不顯著(P> 0.05)(圖2)。
3.2 越冬期不同階段的活動區面積
從越冬期白鶴活動區大小來看,豐水年3 組白鶴90%KDE 活動區面積分別為[(3 771 ± 3 043) hm2,n=7]、[(2 192 ± 352) hm2,n=6]和[(2 935 ± 2 052) hm2,n=13],50%KDE 活動區面積分別為(467 ± 377) hm2、(307 ± 117) hm2和(402 ± 298) hm2。平水年W2 組白鶴的90%KDE 和50%KDE 活動區面積分別為[(1 898 ±1 056) hm2,n=9] 和[(328 ± 220) hm2,n=9],各水位年組間均無顯著差異(P> 0.05)。
從越冬期不同階段的白鶴活動區大小來看,越冬前期,豐水年合并組90%KDE 活動區面積顯著大于平水年W2 組(P= 0.027),其余各組間均無顯著差異;越冬中期,豐水年W1 組的90%和50%KDE 活動面積均顯著大于平水年W2 組和豐水年W3 組(W1/W2 組:P=0.000;P=0.002;W1 組/W3 組:P=0.003;P=0.007),豐水年合并組90%和50%KDE 活動區面積均顯著大于平水年W2 組(P=0.036;P=0.007);越冬后期,豐水年合并組90%KDE 活動區面積顯著大于平水年W2 組(P=0.025)(圖3)。
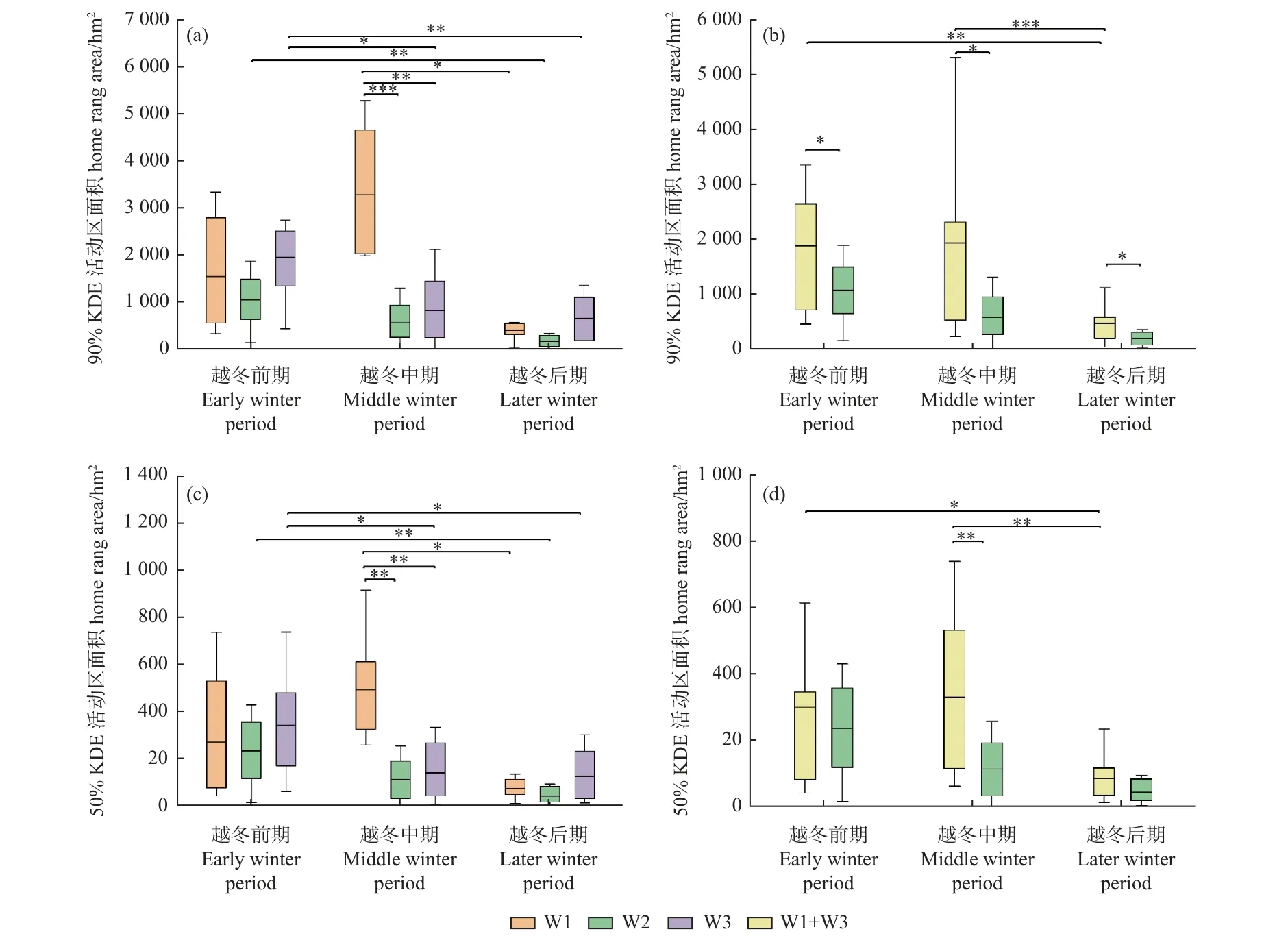
圖3 越冬期不同階段的白鶴活動區面積Fig. 3 Home ranges of Siberian Cranes in different winter stages
從豐水年和平水年越冬期不同階段的活動區面積可知,豐水年W3 組和合并組白鶴的90%和50%KDE活動區面積為越冬前期 > 中期 > 后期,除W3 組越冬中期和后期,以及合并組越冬前期與中期活動區面積差異不顯著,其他階段均差異顯著;W1 組為越冬中期 >前期 > 后期,僅越冬中期和后期差異顯著(P= 0.014;P= 0.025)。平水年W2 組90%KDE 和50%KDE 為越冬前期 > 中期 > 后期,其中僅越冬前期和后期差異顯著(P= 0.004;P= 0.009)(圖3)。
3.3 越冬期不同階段的棲息地利用率
3.3.1 越冬期不同階段的日間棲息地利用率 從白鶴越冬期日間棲息地利用率來看,豐水年3 組白鶴對農田利用率分別為52.2% ±22.7%(n=7)、44.7% ± 20.1%(n=6)和48.2% ± 20.9%(n=13),對濕地利用率分別為44.9% ± 21.1%、55.2% ± 20.0%和50.4% ± 20.5%。平水年組W2 白鶴對農田利用率為11.0% ± 12.3%(n=9),對濕地利用率為(89.0% ± 12.3%)。豐水年3 組白鶴對農田利用率顯著高于平水年組(W1/W2 組:P=0.006;W3/W2 組:P= 0.016;合并組/W2 組:P= 0.000),對濕地利用率顯著低于平水年組(W1/W2 組:P=0.001;W3/W2 組:P= 0.019;合并組/W2 組:P= 0.000)。豐水年組間白鶴對農田和濕地利用率無顯著差異(P> 0.05)。
從越冬期不同階段的白鶴日間棲息地利用率來看,越冬前期和中期,豐水年3 組白鶴對農田利用率均顯著高于平水年組(P< 0.05),對濕地利用率均顯著低于平水年組(P< 0.05)。越冬后期,各組間農田利用和濕地利用率差異不顯著(P> 0.05)(圖4)。
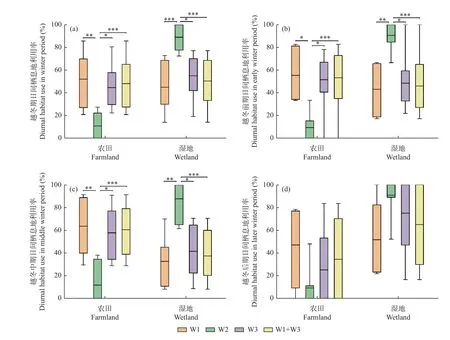
圖4 不同水位年組白鶴的日間棲息地利用比例Fig. 4 Diurnal habitat use ratio of Siberian Cranes in different water year groups
從豐水年和平水年越冬期不同階段的日間棲息地利用率來看,豐水年3 組和平水年組白鶴3 個越冬階段的農田和濕地利用率均無顯著差異(P> 0.05)(圖4)。
3.3.2 越冬期不同階段的夜間棲息地利用率 從白鶴越冬期夜間棲息地利用率來看,豐水年3 組白鶴農田利用率分別為15.7 ± 22.2%(n=7)、15.1% ± 16.1%(n=6)和15.4% ± 18.4%(n=13),濕地利用率分別為84.1% ± 22.1%、84.9% ± 16.0%和84.5% ± 18.4%,平水年組W2 的白鶴農田利用率為5.1% ± 8.7%(n=9),濕地利用率為94.9% ± 8.7%。豐水年組與平水年組的白鶴夜間農田和濕地利用率均無顯著差異(P> 0.05)。
從越冬期不同階段的白鶴夜間棲息地利用率來看,越冬中期,豐水年W1 組和合并組的白鶴農田利用率均顯著高于平水年組(P< 0.05),濕地利用率均顯著低于平水年組(P< 0.05)(圖5)。
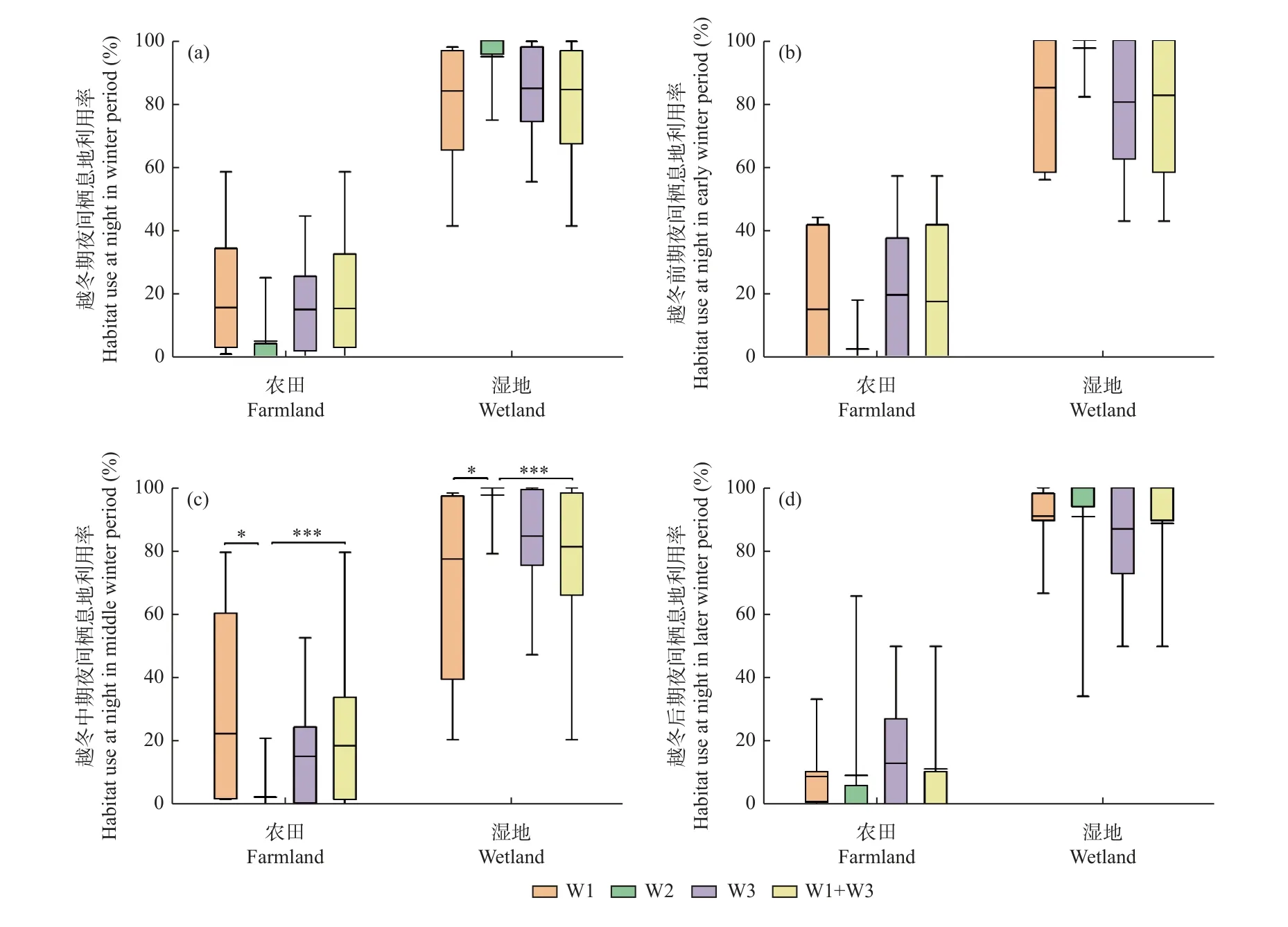
圖5 白鶴在不同水位年組的夜間棲息地利用比例Fig. 5 Habitat use ratio of Siberian Cranes at night in different water year groups
從豐水年和平水年越冬期不同階段的夜間棲息地利用率來看,豐水年3 組和平水年組的白鶴3 個越冬階段的農田和濕地利用率均無顯著差異(P> 0.05)(圖5)。
4 討論
4.1 不同水位年白鶴越冬期的移動模式
避難所理論(refuging theory)認為鳥類每天的移動距離是對棲息地周邊食物資源可獲得性的一種適應性行為,也是增加自身適合度的一種覓食策略(Hamiltonet al.,1970)。隨著時間推移,由于棲息地附近的食物資源枯竭,聚集的鳥類個體將增加棲息地和覓食地之間的移動距離,來滿足能量補給需求;相應地,個體付出代價和風險也隨之增加。每天的移動距離會影響個體的能量消耗和能量平衡,水鳥更多傾向于選擇較近區域覓食(Gauthieret al.,1984;MacArthuret al.,1996;Van der Graafet al.,2005)。當棲息地食物資源缺乏時,鳥類會增加移動距離即去較遠區域覓食,其每日能量攝入與消耗之間的平衡可能會受到移動模式的影響(Pearseet al.,2010;Bavecoet al.,2011)。
研究發現平水年白鶴越冬期移動距離顯著小于豐水年,平水年越冬3 個階段的90%KDE 活動區面積也均顯著小于豐水年合并組的相應越冬階段,這表明平水年白鶴喜食的苦草等自然食物資源充足,白鶴很容易從棲息地及其周圍得到能量補給,無需增加移動成本去較遠區域覓食。這與吳海峰等(2018)對鄱陽湖越冬白枕鶴(Grus vipio)活動區面積隨著水位升高而增大的研究結果一致。平水年白鶴移動距離隨著越冬時間推移而逐漸減小,與越冬中期和后期鄱陽湖水位下降,洲灘裸露,自然食物苦草冬芽的可獲得性增加有關(孫志勇等,2010)。
相反,豐水年苦草等沉水植物的生長受高水位影響,越冬期自然食物資源短缺,白鶴被迫增加移動距離去較遠區域覓食,以滿足自身能量攝入需求。豐水年移動距離隨著越冬時間推移出現先增后減現象,與自然食物短缺和種間競爭有關,導致白鶴越冬前期和中期大量進入藕塘和稻田等農田棲息地覓食,越冬后期白鶴在自然濕地集群以待北遷(王文娟等,2019;Houet al.,2020)。劉成林等(2011)研究表明鄱陽湖水位越高,草灘和淺水區越冬候鳥棲息地越小,鳥類生存的自然棲息空間進一步縮小。枯水期鄱陽湖水位14.18 m(星子站,黃海高程)以上時, 水陸過渡帶將縮小乃至消失,鄱陽湖濕地作為候鳥棲息地的功能將喪失。
4.2 不同水位年白鶴越冬期的覓食策略
食物資源的豐富度與可獲得性是影響越冬水鳥覓食棲息地利用的重要因素(Luotoet al.,2007;Saygiliet al.,2011)。食物資源對動物活動區大小產生重要影響,豐富的食物資源使得個體活動區較小,貧乏的食物資源則使得個體活動區較大(B?rgeret al.,2008;Corrialeet al.,2013)。水位通過影響食物資源分布和可獲得性,間接影響越冬候鳥活動區大小(劉成林等,2011;Pickens et al.,2017;吳海峰等,2018)。近些年來,由于鄱陽湖水文情勢發生變化,白鶴利用農田棲息地的比例呈現不同程度增加,這得到鳥類學家的廣泛關注(鐘毅峰,2020;Hou et al.,2021;底明曉等,2022),但這些調查研究主要基于某個越冬階段或某個越冬區域的調查,可能會因時間或空間局限導致對白鶴整個越冬期棲息地利用模式產生偏差,這也是與本研究結果有所不同的主要原因。
平水年,白鶴自然食物資源的可獲得性是其覓食策略的主要影響因素,與平水年白鶴越冬期日間利用濕地生境比例達到89.0% ± 12.3%(n = 9)直接相關。研究發現平水年白鶴活動區面積在越冬前期顯著大于中期和后期,與不同越冬階段各水位年組移動距離變化趨勢基本保持一致。Ivey 等(2015)對越冬期沙丘鶴(Grus canadensis)的移動距離和活動區大小研究發現,飛行移動距離越小,活動區面積也越小。平水年苦草等沉水植物正常生長,張笑辰等(2014)對鄱陽湖越冬食性相似的白鶴和小天鵝(Cygnus columbianus)的研究發現,越冬前期苦草冬芽分布的水深大于中期,由于白鶴形態學上的限制,導致其無法在水位較深的區域取食,而小天鵝可以在更深區域選擇食物資源更豐富的覓食地。豐水年,在自然食物資源匱乏條件下,白鶴會從自然濕地轉移到周邊藕塘和稻田(王文娟等,2019;底明曉等,2022)。豐水年越冬期日間白鶴利用農田棲息地的比例達到50%左右,其中越冬前期和中期對農田棲息地的利用比例高于自然濕地,尋找適宜覓食地和種間競爭是影響其覓食策略的主要因素。豐水年日間白鶴提高對農田的利用率,與薛琳(2019)對不同越冬期鄱陽湖越冬白鶴日間生境選擇研究結果一致。自然濕地中的苦草冬芽易消化(Foxet al.,2011),白鶴容易獲取;而在藕塘中蓮藕埋藏深且體積大,不易獲取;稻田中稻谷易獲取,但纖維素含量高,不易消化(植毅進等,2019)。盡管 如此,自然濕地中白鶴覓食時間顯著高于農田生境,警戒時間也顯著要低(袁芳凱等,2014;邵明勤等,2018)。因此,豐水年白鶴轉移到農田生境覓食,是增加自身適合度的一種適應性行為,但仍不會放棄自然濕地覓食棲息地。
不論豐水年還是平水年,白鶴夜間主要利用自然濕地,這與胡振鵬等(2012)的觀察結果一致。在豐水年的越冬中期,由于自然食物資源短缺與種間競爭,白鶴可能會增加農田覓食時間,較晚返回自然濕地,導致其夜間農田利用率顯著高于平水年組。有關白鶴的食性改變是否影響其生理健康狀況等還需開展相關研究,以期為白鶴越冬棲息地管理提供科學依據和支撐。
5 結論
豐水年白鶴通過增加移動距離、活動區面積以及農田棲息地利用比例來應對水位變化造成的食物資源短缺,滿足能量需求來提高自身適合度。豐水年白鶴日間對農田利用率僅50%左右,夜間仍主要利用自然濕地;平水年白鶴的主要覓食地和夜宿地仍是自然濕地。因此,應加強鄱陽湖區自然濕地適應性管理,以提高不同水文條件下的濕地功能和恢復力,確保越冬白鶴的健康穩定與越冬安全。
