不同類型鹽生植物適應鹽脅迫的生理生長機制及Na+逆向轉運研究進展
2023-02-02 06:46:34劉佳欣張會龍鄒榮松楊秀艷朱建峰張華新
生物技術通報 2023年1期
關鍵詞:植物
劉佳欣 張會龍 鄒榮松 楊秀艷 朱建峰 張華新
(1.中國林業科學研究院生態保護與修復研究所 國家林業和草原局鹽堿地研究中心,北京 100091;2.中國林業科學研究院天津林業科學研究所,天津 300457;3.中國林業科學研究院黃河三角洲綜合試驗中心,東營 257500)
鹽漬土是一系列受土體中鹽堿成分作用的土壤,是各種鹽土和堿土以及其他不同程度鹽化和堿化的各種類型土壤的統稱,也稱鹽堿土[1]。據統計,世界鹽漬土面積約為9.543 8×109hm2,廣泛分布于100 多個國家和地區。我國是世界鹽漬土面積第三大國,鹽漬土面積達9.913 3×108hm2,約占全國土地面積的10%,分布在23 個省、市、自治區,其中,西北、華北、東北及沿海地區是我國鹽漬土的主要集中分布地區[2]。在世界范圍內,鹽漬土帶來了人口、資源、環境等諸多問題。鹽漬土環境不利于植物的生長,鹽漬土中過量礦質元素如Na+或Cl-對植物產生的不利影響,稱為鹽脅迫[3]。鹽脅迫通過滲透脅迫和離子脅迫在整個植物和細胞水平上影響植物生理[4-5],如過量Na+會干擾K+的吸收和代謝,K+參與酶的活化、蛋白質的合成、滲透調節、光合作用等生理過程[6],K+缺乏會引起呼吸速率和離子毒性增加,植物生長變化,礦物分布改變,以及光合作用速率降低[7],還會引起枯萎病,繼而導致植物葉片壞死;此外Na+取代Ca2+造成膜不穩定[8],膜通透性改變[9],導致植物減產甚至死亡[10]。鹽漬土中存在一種特有的鹽生植物生態系統,系統中的鹽生植物不僅能夠在鹽漬土中生長,還能夠改善鹽漬土環境[11]。
大多數植物不能適應鹽漬土環境,但鹽生植物(halophytes)能在離子濃度至少200 mmol/L 以上的生境中生長和完成生活史[12]。例如唐古特白刺(Nitraria tangutorum)、中國檉柳(Tamarix chinensis)、芨芨草(Achnatherum splendens)和黃花補血草(Limonium aureum)[13]等。Brekle 根據鹽生植物體內離子運輸和積累的特點,將其分成稀鹽鹽生植物(euhalophytes)、泌鹽鹽生植物(recretohalophytes)和拒鹽鹽生植物(pseudohalophytes)3 種。
離子區隔化(ion compartmentalization)利用特定轉運蛋白對有毒離子進行細胞分隔,是鹽生植物維持細胞內適度的Na+/K+比的一種關鍵模式[5]。鹽生植物通過調節離子的吸收和將離子區隔化來抵抗或減輕鹽脅迫造成的傷害,主要是通過將過量Na+區隔在植物組織器官的局部或細胞的局部,甚至排出體外,從而避免或降低Na+對植物正常生長和生理代謝的影響。本文重點討論了3 類鹽生植物的生長形態特征,并按植物體內對Na+運輸和積累特點對3 種鹽生植物的典型耐鹽機制研究進展進行了闡述,旨在為今后鹽生植物及其耐鹽機制研究提供相關依據,為植物耐鹽分子育種奠定基礎。
1 稀鹽鹽生植物耐鹽機制
稀鹽鹽生植物,也叫真鹽鹽生植物,是將鹽離子區隔在肉質化組織和液泡中的一類鹽生植物[14]。稀鹽鹽生植物是鹽生植物中耐鹽性最強的一類植物,多能在200-400 mmol/L 的鹽漬土環境中生長[1]。稀鹽鹽生植物絕大部分是藜科植物,例如鹽地堿蓬(Suaeda salsa)、鹽爪爪(Kalidium foliatum)、鹽穗木(Halostachys caspica)等。
1.1 稀鹽鹽生植物耐鹽形態特征
稀鹽鹽生植物的內在結構和外觀形態在一定程度上適應鹽脅迫,其形態上最突出的特征是莖葉肉質化。肉質化組織含水量高,可以將體內鹽分稀釋,使植物體不受高鹽環境的傷害[15]。莖肉質化稀鹽鹽生植物,皮層分化為內部皮層和外部皮層兩部分,內部皮層發育成貯水組織,細胞呈球形或橢圓形,外部皮層發育成柵欄組織,細胞內含大量葉綠體。鹽節木(Halocnemum strobilaceum)、鹽穗木[16]等植物都是莖肉質化的稀鹽鹽生植物,其形態存在以上特點。葉肉質化的稀鹽鹽生植物,葉肉質且無柄,呈圓柱狀或半圓柱狀,氣孔少,葉表皮細胞排列緊密,幾乎全部由貯水細胞組成,表皮外被角質層。Gao等[17]研究表明假木賊(Halocnemum strobilaceum)在鹽溶液濃度為0.9%-5.4%時,葉肉質化程度增加。劉彧等[18]研究表明鹽地堿蓬葉片對Na+和Cl-的積累是葉片肉質化的主要原因,在鹽脅迫下,葉片肉質化,細胞數目增多。另外,堿蓬屬(Suaeda)、鹽爪爪屬(Kalidium)、假木賊屬(Anabasis)的植物,也是葉肉質化的稀鹽鹽生植物。
1.2 稀鹽鹽生植物耐鹽生理機制
稀鹽鹽生植物除憑莖葉肉質化稀釋植物體內鹽分外,主要是通過離子區隔化作用,將離子運輸到細胞的特定部位,從而降低鹽離子對胞質中的細胞器、酶等的毒害,且伴隨著細胞濃度的增大,水勢降低,細胞吸水及抗滲透脅迫能力增強[19]。劉彧等[18]發現鹽地堿蓬葉片細胞將Na+區隔到液泡中,以減少Na+對植物造成的滲透脅迫及離子脅迫,有利于鹽地堿蓬在高鹽條件下生存。西伯利亞白刺(Nitraria sibirica)通過Na+液泡區隔化增強葉片組織的耐鹽性,保持胞質溶液中較低Na+含量,提高滲透調節能力,從而在鹽脅迫下維持正常生理活動[20]。1.3 稀鹽鹽生植物耐鹽分子機制
細胞內Na+/K+濃度的平衡是保證植物在鹽漬環境下進行正常生理代謝的關鍵。稀鹽鹽生植物在鹽脅迫條件下,通過鹽誘導Na+/H+逆向轉運體將Na+從胞質中排出或在液泡中分隔[21],其中質膜Na+/H+逆向轉運蛋白SOS1(plasma membrane Na+/H+exchanger 1)能夠將胞質中過量的Na+排出到胞外;液泡膜Na+/H+逆向轉運蛋白NHX(vacuolar Na+/H+antiporter)如NHX1 和NHX2 能夠將胞質中過量的Na+逆濃度梯度轉運至液泡中,SOS1 和NHX 均可減少胞質內Na+的累積,維持細胞的離子穩態,抵御鹽脅迫[22]。植物分泌途徑的細胞膜上同時存在兩類致電H+泵,液泡膜H+-腺苷三磷酸酶(vacuolar H+-ATPase,簡稱液泡膜H+-ATPase)、液泡膜焦磷酸酶(vacuolar H+-pyrophosphatase,簡稱液泡膜H+-PPase)[23]和質膜H+- 腺苷三磷酸酶(plasma membrane H+-ATPase,簡稱質膜H+-ATPase),它們為離子區隔化作用提供能量,并催化H+由胞質向液泡和胞質向胞外的運輸[24]。鹽脅迫下,鹽地堿蓬質膜H+-ATP 酶和液泡膜H+-ATP 酶的表達和活性顯著升高[25]。Liu 等[26]用不同濃度的NaCl(100、200、300 和400 mmol/L) 和Ca2+(0、5、10、15、20 mmol/L)處理唐古特白刺幼苗時發現,鹽脅迫(NaCl濃度為300 mmol/L)顯著促進液泡膜H+-ATP 酶和H+-PPase 酶活性。一定濃度的外源Ca2+有效增強了唐古特白刺液泡膜和質膜的離子泵活性,促進細胞內Na+區隔化,從而提高唐古特白刺的耐鹽性。
Qiu 等[27]從擬南芥(Arabidopsis thaliana)中分離鑒定了位于質膜的Na+/H+逆向轉運蛋白SOS1和位于液泡膜的Na+/H+逆向轉運蛋白NHX1,認為這2 個轉運蛋白共同作用使胞質中Na+維持在較低濃度[28]。Na+/H+逆向轉運蛋白依靠跨液泡膜的H+濃度梯度對Na+進行轉運,因此可以推測通過超表達液泡膜H+-ATPase,使質子梯度增加,能夠提高NHXs 對 Na+的區隔能力。在鹽脅迫條件下,西伯利亞白刺Na+/H+逆向轉運蛋白NsNHX1 和NsSOS1 的表達水平顯著上調,過表達NsSOS1 減少了轉基因植株Na+的含量,過表達NsNHX1 增加了轉基因植株Na+的含量,表明NsSOS1 介導過量Na+的外排,NsNHX1 介導過量Na+的區隔化,它們對提高轉基因植物耐鹽性有重要作用[22]。另外,有研究表明,NHX 類蛋白在K+區隔化進入液泡和維持細胞內的pH 平衡中也起著重要作用[29](表1)。

表1 稀鹽鹽生植物關鍵耐鹽基因Table 1 Key salt-tolerant genes in euhalophytes
2 泌鹽鹽生植物耐鹽機制
泌鹽鹽生植物是一類通過鹽腺或鹽囊泡將吸收到體內的鹽分分泌到體外的鹽生植物。大部分泌鹽鹽生植物的耐鹽性低于稀鹽鹽生植物[1]。常見的泌鹽鹽生植物有中華檉柳(Tamarix chinensis)、二色補血草(Limonium bicolor)等。
2.1 泌鹽鹽生植物耐鹽形態特征
泌鹽鹽生植物的主要形態特征是其葉片或莖上形成了專門分泌鹽分的鹽腺或鹽囊泡,鹽腺或鹽囊泡可以分泌植物體內過量的鹽離子,因此鹽腺和鹽囊泡對泌鹽鹽生植物適應鹽脅迫起重要作用。根據鹽離子運輸方式的不同,又可將泌鹽鹽生植物分為向內和向外泌鹽植物。向內泌鹽植物(endoreoretohalophytes)將體內的鹽分分泌到葉表面的囊泡中暫時貯存,而后鹽囊泡破裂將體內過量鹽離子排出。鹽囊泡有1 個柄和1 個膨大的泡狀細胞,柄細胞通過大量胞間連絲與泡狀細胞、葉肉細胞相連。藜(Chenopodium album)的鹽囊泡是從莖尖和腋芽處的莖葉表皮細胞分化而來,部分表皮細胞生長變大,凸起于莖葉表面,形成鹽囊泡的原始細胞,原始細胞進一步分裂分化為1 個柄細胞和1 個泡狀細胞,成熟后的鹽囊泡外被角質層[44]。利用掃描電鏡發現四翅濱藜(Atriplex canescens)鹽囊泡與其它藜科植物鹽囊泡結構相似,是由體積龐大的腎形囊泡細胞、細長的柄細胞以及基部的表皮細胞共同構成鹽囊泡復合體,且隨著葉齡增加,鹽囊泡逐漸破裂退化[45]。
向外泌鹽植物(exo-reoretohalophytes)利用鹽腺和分泌細胞將鹽離子分泌到體外。鹽腺分為單細胞鹽腺、雙細胞鹽腺和多細胞鹽腺[46]。單細胞鹽腺有大液泡和很少的細胞器,具分泌細胞鹽腺。目前對單細胞鹽腺鹽生植物研究較少。雙細胞鹽腺由1個基細胞和1 個帽細胞組成,兩個細胞通過胞間連絲相連[47],細胞質稠密,有一個明顯細胞核,沒有中央液泡,且腺體細胞壁角質化或栓質化。Kuster等[48]通過對報春花(Jacquinia armillaris)葉片腺狀毛的解剖和組織化學研究發現,其葉片鹽腺是由一個收集細胞和一個單柄細胞組成,其頂端有多個分泌細胞。非洲虎尾草(Chloris gayana)是具有雙細胞鹽腺的泌鹽鹽生植物[49]。另外,禾本科的獐毛屬(Aeluropus)、鼠尾粟屬(Sporobolus)、大米草屬(Spartina)、結縷草屬(Zoysia)等9 個屬也存在雙細胞鹽腺[50]。雙子葉泌鹽鹽生植物的鹽腺均為多細胞鹽腺,多細胞鹽腺呈球形或盤狀結構,由基底收集細胞和遠端分泌細胞構成,整個結構常陷入表皮中[46]。馮中濤[51]研究發現二色補血草鹽腺由16個細胞組成,中央為4 個長的分泌細胞,每個分泌細胞外側均有1 個毗鄰細胞,再向外依次包圍著4個內杯狀細胞和4 個外杯狀細胞,鄰近的細胞稱為收集細胞;細胞核較大,富含線粒體、高爾基體和小囊泡,但缺少中央大液泡,也沒有葉綠體;細胞之間通過胞間連絲連接,鹽腺外被角質層。不同的鹽生植物其鹽腺呈現不同的形狀。
2.2 泌鹽鹽生植物耐鹽生理機制
泌鹽鹽生植物通過鹽腺或鹽囊泡向體外分泌植物體內過多的鹽分,避免鹽分在體內過量積累,從而應對鹽脅迫對植物的傷害[52]。關于泌鹽鹽生植物分泌鹽的機制,前人經過大量研究,從組織化學和超微結構的角度提出了3 個假說:(1)1955年Arisz等[53]提出滲透機制假說,認為鹽腺的泌鹽是一個物理過程,鹽腺中離子的主動積累使分泌細胞滲透壓降低,從而導致靜水壓力的顯著增加。當鹽腺內壓力達到最高時,分泌細胞通過分泌孔周期性地形成液滴,釋放靜水壓力,將離子排出鹽腺外[54]。鹽草(Distichlis spicata)的超微結構證實了這一假說,其鹽腺只有一層薄膜,類似于一個瓣膜,將細胞質壁層隔開[55]。滲透機制假說還受到了一些研究結果的挑戰,鹽腺泌鹽存在離子選擇性和主動運輸的特征,即鹽腺泌鹽不是簡單的物理過程[56]。(2)1967年Ziegler 和Luttge[57]提出胞飲反向活動假說,認為鹽腺的分泌作用是通過貯存在小細胞質囊泡中的鹽液向細胞外表面排出的過程。通過使用電子顯微鏡對二色補血草[58]和檉柳[59]鹽腺進行掃描,觀察到電子密度物質主要聚集在囊泡中,特別是在鹽處理的植物中,大量的囊泡與質膜融合,從而證明了這一假說。(3)1971年Levering 和Thomson[60]提出鹽腺的分泌作用類似于動物液流運輸系統假說,研究者假設大米草屬基底細胞形成了廣泛的膜系統,鹽腺通過膜上的離子通道分泌離子,類似于動物組織上皮細胞的吸收和分泌[52]。
鹽囊泡的泌鹽方式不同于鹽腺,鹽腺直接將鹽分分泌至體外,而鹽囊泡則是通過積累大量鹽分并膨脹,最終破裂,將鹽分釋放到植物體外[61],具體過程主要包括:Na+從葉肉細胞排出,經柄細胞轉運至鹽囊泡,最后被區隔在鹽囊泡液泡[62]。研究表明,四翅濱藜鹽囊泡Na+積累過程是Na+沿表皮細胞-柄細胞-鹽囊泡復合體(EC-SC-EBC complex)逆著其濃度梯度的單向極性跨膜轉運[63]。Barkla 等[64]通過對冰葉日中花(Mesembryanthemum crystallinum)鹽囊泡中的離子組成分析發現,其中含有大量Na+和Cl-,并進一步證明離子區隔化提高了該植物的耐鹽性。
2.3 泌鹽鹽生植物耐鹽分子機制
泌鹽鹽生植物鹽的分泌是植物活躍的生理過程。在鹽脅迫條件下,H+-ATPase 的表達隨著鹽分泌的增加而增加[65]。張春蕊等[66]在剛毛檉柳(Tamarix hispida)中鑒定獲得液泡膜H+-PPase 基因ThVP1,發現在NaCl 脅迫下,剛毛檉柳根和葉中的ThVP1基因表達明顯上調。在對二色補血草的鹽腺結構觀察時發現其鹽腺中不存在葉綠體,且鹽腺中的線粒體比葉肉細胞的線粒體大,證明線粒體能夠為鹽腺分泌鹽分提供能量[56]。Panda 等[67]指出通過三羧酸循環的上調來持續提供能量,同時不間斷地合成蛋白質和維持光合機制的結構完整性,是柳枝梭梭(Haloxylon Sal Icornicum)耐鹽的主要機制。
Hamada 等[68]從鹽生植物北濱藜(Atriplex gmelini)中分離出一個Na+/H+逆向轉運蛋白基因,并將其命名為AgNHX1,發現AgNHX1的遷移模式與液泡膜H+-ATPase 相關,而與液泡膜H+-PPase 無關,AgNHX1定位于液泡膜,鹽脅迫下,在mRNA和蛋白水平上均上調表達。通過AlNHX轉化的鹽生植物小獐毛(Aeluropus littoralis)表現出較高的耐鹽性(400 mmol/L NaCl)[69],由此可以看出AgNHX1產物在耐鹽性中具有重要作用[70]。Dang[71]和Yuan等[56,72]陸續在紅砂(Reaumuria trigyna)和補血草屬(Limonium)植物中進行了轉錄組測序,獲得了參與鹽腺分泌的一系列候選基因,為提高植物耐鹽性提供可能(表2)。
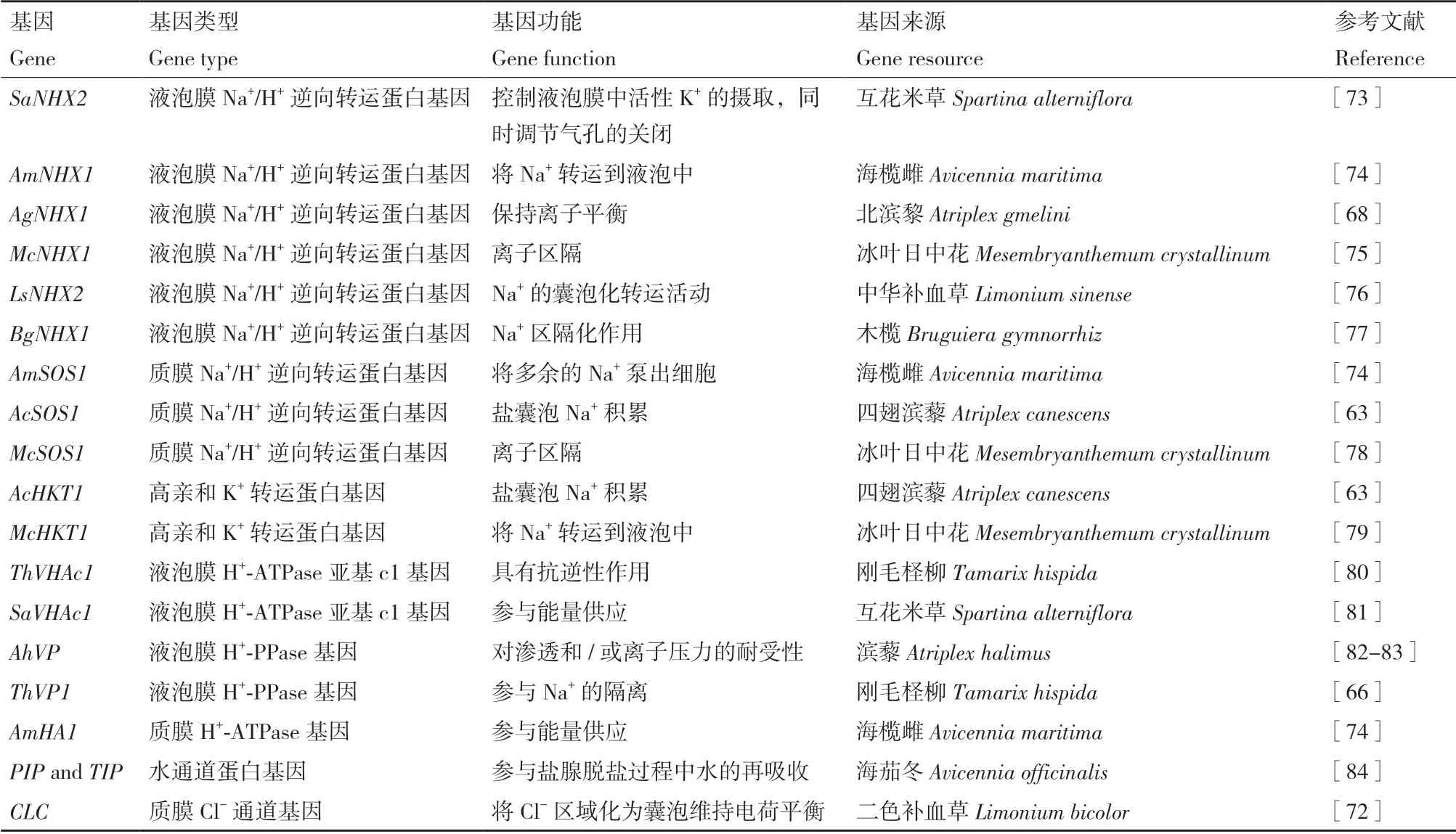
表2 泌鹽鹽生植物關鍵耐鹽基因Table 2 Key salt-tolerant genes in recretohalophytes
3 拒鹽鹽生植物耐鹽機制
拒鹽鹽生植物,又稱假鹽鹽生植物,拒絕或少吸收外界鹽分,能夠將鹽離子積累在皮層細胞液泡和根部木質部的薄壁細胞中,減少向上部莖葉運輸鹽離子[14],降低鹽分對植物的脅迫作用。沙棗(Elaeagnus angustifolia)[85]、蘆葦(Phragmites australis)[86]、莎草(Cyperus rotundus)等是較為常見的拒鹽鹽生植物。拒鹽鹽生植物種類較稀鹽鹽生植物和泌鹽鹽生植物少[1]。
3.1 拒鹽鹽生植物耐鹽形態特征
拒鹽鹽生植物根部皮層栓質化,栓質化的主要成分是難溶于水的脂肪類物質,具有不透水、不透氣性,導致溶解于土壤水分中的鹽分難以進入根中;莖的表皮切向壁具有發達的角質層,維管束近于星散分布;葉的上表皮向外延伸的表皮毛較多,氣孔下陷;葉肉組織排列疏松,胞間隙較大,葉脈維管束鞘由兩層細胞構成[87]。在高濃度鹽脅迫下,拒鹽植物小花堿茅(Puccinellia tenuiflora)的內皮層顯著加厚,根內皮層細胞表面的木栓質沉積受鹽脅迫誘導而增強[88]。經過鹽處理的燈芯草屬(Juncus)植物,根和莖中具有發育良好的通氣組織,且根皮脫落,皮質很厚,凱氏帶(Casparian strips)木質化[89],對植物抵抗鹽脅迫發揮重要作用。
3.2 拒鹽鹽生植物耐鹽生理和分子機制
拒鹽鹽生植物的拒鹽機理,不同植物不盡相同。有些拒鹽鹽生植物拒Na+或者拒Cl-,有些既拒Na+又拒Cl-[1]。蘆葦在鹽脅迫下,Na+轉運速率從根至芽依次降低,從而避免在芽中積累過量的Na+[90]。陳琳等[91]對潮灘蘆葦和河灘蘆葦的耐鹽性進行比較發現,因潮灘蘆葦的根系有更高的排Na+能力而耐鹽性更強。彭彥輝[92]通過觀察小花堿茅根系電鏡掃描結果發現,控制Na+/K+選擇性吸收是發生在根內皮層上的。小花堿茅通過較高的Na+/K+選擇性吸收能力,能減少Na+在體內的凈積累[93]。Abideen等[94]和王樹鳳等[95]觀察到,根系生長可以補償其吸收功能的損失,由于鹽脅迫引起的干重積累和表面積的增加,這些補償效應有利于根系對K+的吸收和Na+的貯存,由此推測根系補償性生長可能是沙棗增強耐鹽性的一種機制[96]。Liu 等[96]研究進一步證實了沙棗根Na+和Cl-貯存的可擴展能力,有效地限制Na+從莖到葉的運輸,以及根補償生長是沙棗對鹽脅迫的生理適應機制。
Perez-Jimenez 等[97]通過γ 輻射獲得具有抗鹽性的5 個柑橘(Citrus×paradisi)砧木突變體,并對突變體抵抗鹽度水平的程度進行評估,數據表明柑橘是通過積累較少Na+或 Cl-來提高耐鹽性。Zhang等[98]發現PtNHX1、PtSOS1 和PtHKT1;5 等轉運蛋白在小花堿茅的不同組織協同調控,使植物維持Na+穩態,增加小花堿茅的耐鹽性。李靜[99]采用非損傷微測技術分析小花堿茅響應鹽脅迫的各種離子流動態變化機制,推測小花堿茅在高鹽脅迫初期,Ca2+的大量內流可能在SOS 系統信號轉導過程中發揮了關鍵作用,從而激活SOS1 以增強Na+外流。楊升等[100]研究指出,在鹽脅迫下,沙棗幼苗根系進行Na+積累和K+外流,為減少Na+積累,通過根系Na+/H+逆向轉運蛋白將Na+從體內排出(表3)。

表3 拒鹽鹽生植物關鍵耐鹽基因Table 3 Key salt-tolerant genes in pseudohalophytes
4 總結與展望
通過對3 類鹽生植物耐鹽形態、生理及分子機制的總結發現,大部分鹽生植物主要是通過兩類基因應對胞質中過高的Na+,其中一類是離子轉運蛋白編碼基因,如液泡膜Na+/H+逆向轉運蛋白編碼基因NHX1和質膜Na+/H+逆向轉運蛋白編碼基因SOS1;另一類是提供能量的基因,如液泡膜 H+-ATPase 和H+-PPase 基因以及質膜H+-ATPase 基因。液泡膜H+-ATPase 和H+-PPase 及質膜H+-ATPase 產生H+梯度并提供能量,液泡膜Na+/H+逆向轉運蛋白NHX1 將胞質中過量的Na+逆濃度梯度轉運至液泡中,質膜Na+/H+逆向轉運蛋白SOS1 將胞質中過量的Na+排出到胞外,這樣就可以使植物胞質中的Na+濃度降低到植物耐受的范圍內。
在生理和形態上,3 類鹽生植物應對鹽脅迫的策略各有不同。稀鹽鹽生植物通過離子區隔化應對鹽脅迫,將過量Na+區隔在液泡中,并通過莖葉肉質化將體內鹽分進行稀釋,使植物體不受高鹽環境的傷害(圖1-a)[102-103];泌鹽鹽生植物擁有其特有的生理結構——鹽腺或鹽囊泡,能夠利用鹽腺和分泌細胞將鹽離子分泌到體外,或是把體內鹽分分泌到葉表面的鹽囊泡中暫時貯存,而后鹽囊泡破裂將體內多余鹽分釋放,從而避免鹽分在體內過量積累以維持植物體正常生理活動(圖1-b)[104-105];拒鹽鹽生植物通常是將鹽離子積累在皮層細胞液泡和根部木質部薄壁細胞中,減少向上運輸鹽離子,且根外皮層栓質化不透水透氣性,進一步實現不吸收或很少吸收鹽離子以適應鹽脅迫(圖1-c)[29]。經前人大量研究,大部分的稀鹽鹽生植物耐鹽性高于泌鹽鹽生植物,高于拒鹽鹽生植物,這3 種鹽生植物耐鹽性的不同,可能與其生理生長機制的差異有關,但仍需要進行大量的實驗進行驗證。

圖1 三類鹽生植物Na+逆向轉運耐鹽機制模式圖Fig.1 Model diagram of Na+ antiport mechanisms for salt tolerance in three halophytes
除了以上研究,還有許多問題亟待進一步探究:一是離子轉運蛋白和提供能量的兩大類基因在各類鹽生及非鹽生植物中均存在,但不同植物的耐鹽性差異巨大,因此,應進一步挖掘鹽生和非鹽生植物耐鹽基因,比較不同種植物間相同基因如NHX1、SOS1的功能差異,研究相同基因在不同植物體內的調控機制,并進行基因功能的驗證分析;二是決定及調控鹽生植物在鹽脅迫下特化出組織結構的基因需要進一步挖掘,如稀鹽鹽生植物的肉質化莖和葉、泌鹽鹽生植物的鹽腺和鹽囊泡、拒鹽鹽生植物栓質化根;三是植物耐鹽是一個由多基因協調控制的復雜生物學過程,以往的研究大多是將單一目的基因轉入到植物體內以提高植物的耐鹽性,今后可以將離子轉運蛋白和提供能量的兩類基因同時轉化到植物中,與單一基因的轉化進行比較,探索更有效的多基因轉化組合,為耐鹽植物的育種提供依據。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
