歐文氏菌DE2對三種豆科牧草幼苗生長的影響
2023-10-08 08:57:06何林鑫田望軍靳振海陳海雁張振粉
草地學報 2023年9期
關鍵詞:生長
何林鑫, 黃 榮, 田望軍, 靳振海, 趙 亮, 陳海雁, 張振粉
(甘肅農業大學草業學院, 草業生態系統教育部重點實驗室, 中-美草地畜牧業可持續發展研究中心, 甘肅 蘭州 730070)
梨火疫病(Fire blight)是薔薇科植物的毀滅性病害之一,該病在北美一直是梨(Pyrusspp.)、蘋果(MaluspumilaMill.)、櫻桃(Prunusspp.)等果樹的毀滅性細菌病害,也是最古老、研究最深入的植物病害之一[1]。由于受侵染的花、枝條和病葉會變成黑褐色并呈現出枯萎的癥狀,但是仍掛在樹上不脫落形似火燒一樣,故稱此病害為“火疫病”[2-3]。2016年5月梨火疫病在新疆伊犁州首次報道發生[4]。目前,梨火疫病已在我國新疆、甘肅2個省(自治區)65個縣、市(區)[5]發生并對果農造成嚴重損失,其病菌幾乎能夠侵染寄主植物的所有組織[6];寄主植物的花被病菌感染后呈現整個花序呈黑色,干枯萎蔫下垂;葉片被感染后會出現葉脈褪綠黃化,最終失水變為褐色[7];果實被感染后果面開始變黑、腐爛并出現乳白色的菌膿[8];枝干感病后呈現皮層收縮、下陷,形成較大的梭形紅褐色或黑褐色干腐狀病斑[9]。除此之外,還有許多研究證實,梨火疫病菌會引起植物自身體內存在的防御機制酶的變化,王宏等(2019)在對4種早熟梨葉片進行抗病性鑒定時,發現病原菌會誘導了過氧化物酶(Peroxidase,POD)、超氧化物歧化酶(Superoxide dismutase,SOD)活性的上升[10]。何臨梓(2022)在對4種梨砧木對梨火疫病進行抗病相關酶活性變化分析時,發現梨火疫病病原菌誘導了4種梨砧木過氧化氫酶(Catalase,CAT)、超氧化物歧化酶(Superoxide dismutase,SOD)、多酚氧化酶(Polyphenol oxidase,PPO)等五種酶活性的升高和丙二醛(Malondialdehyde,MDA)含量的增加[11]。因此,梨火疫病病菌對植物生長的影響需要被高度關注。
甘肅是草地資源大省,同時也是我國草地畜牧業占重要地位的省份之一,全省草地總面積1 790萬hm2,位居全國第6位[12],其中,紫花苜蓿(Medicagosativa)作為優質牧草是甘肅省的主要種植牧草,其在甘肅省的種植面積占全國種植面積的1/3,位居全國之首[13];草木犀(Melilotusofficinalis)對環境具有很強的適應能力,作為重要的鹽堿地改良作物在甘肅大面積種植[14];沙打旺(Astragalusadsurgens)是我國重要的栽培豆科牧草,具有出色的抗風沙能力,也是甘肅主要的種植牧草之一[15]。不可否認,病害也是牧草生產的主要限制因素之一,可直接影響牧草的生長,降低牧草產量、影響牧草品質,嚴重時導致植物死亡,從而引致草地早衰和退化,甚至導致家畜中毒,降低家畜生產性能[16]。更為重要的是,目前梨火疫病已經傳入甘肅省武威市(涼州區,民勤縣),張掖市(甘州區,肅南縣,民樂縣,臨澤縣,高臺縣,山丹縣)[17]。由于受到地理、環境和氣候等因素影響,甘肅部分地區不適宜種植糧食作物,農民為了提高收益選擇種植果樹并在林間種植牧草作為飼料來以飼養家畜[18],已經形成了林草交錯的生態群。然而,果樹感染梨火疫病,其感病組織脫落后掉入林間草地中,梨火疫病病菌直接與牧草接觸,是否會感染林下常見豆科牧草,進而限制草地生產率,還缺少相關科學研究。一旦新發病原微生物進入當地生態系統,將嚴重威脅生態健康和限制農牧業的可持續發展。
綜上所示,本試驗以紫花苜蓿、草木犀、沙打旺為供試豆科牧草,以本實驗室已分離鑒定的梨火疫病優勢致病菌株歐文氏菌DE2(Erwiniasp.DE2)為供試病原細菌菌株,通過生長瓶[19]萌發試驗測定DE2接種后對3種牧草種子的苗期生長及體內相關酶活性的影響,分析接種DE2后的3種牧草苗期生長和生理指標,來確定梨火疫病的優勢致病菌對我國主要栽培草地的豆科牧草的潛在危害,以期為評估和綜合防控林草系統中新發病原微細菌提供科學基礎。
1 材料與方法
1.1 試驗材料
供試種子:草木犀、紫花苜蓿、沙打旺,由甘肅省農科院提供,并保存在4℃冰箱。
供試菌株:梨火疫病的優勢致病菌——歐文氏菌DE2,由本實驗室分離鑒定并保存,該菌GenBank注冊號為:OQ608623。
供試培養基:大豆酪蛋白瓊脂培養基(Tryptose soya agar,TSA/TSB)[20],主要用于菌株純化。
1.2 菌株培養
參照劉華威[21]的細菌培養方法稍作改動。首先將供試菌株接種于TSA培養皿上,放入28℃培養箱中培養24 h后進行活化,再挑取單菌落利用平板劃線法接種于TSA培養皿上放入28℃培養箱中培養24 h,然后挑選單菌落接種于TSB培養液中,放入恒溫培養箱中,設置溫度為28.5℃,轉速為160 r·min-1培養24 h,將菌液離心后棄置上清液,加入無菌水震蕩搖勻,將其懸浮為濃度108cfu·mL-1的菌懸液[22]。
1.3 確定菌懸液濃度建立DE2的標準曲線方法
參照朱艷靜[23]的方法,有改動。取6支無菌試管,取培養好的菌懸液,按照不同的設定稀釋倍數(5,10,20,50,100,200倍),加入無菌水依次進行稀釋,用分光光度計在600 nm標準波長下測得一組不同濃度菌懸液的吸光度,測試前用無菌水進行調零,在無菌工作臺上吸取上述稀釋得到的不同濃度梯度的菌懸液,分別用無菌水按10倍比例稀釋成不同濃度的菌懸液。取稀釋倍數在10-4~10-8每個濃度的菌懸液吸取1 mL注入TSA培養皿上進行平板涂布,放置28℃的培養箱中培養24 h后用肉眼觀察,點出菌落數后記錄,挑選菌落數在30~300個的平板作為菌落總數測定的標準,并乘以其對應的稀釋倍數,最終計算出不同濃度菌懸液的菌液濃度。以吸光度ABS值為橫坐標,菌懸液濃度為縱坐標繪制標準曲線。
1.4 發芽試驗和接種方法
采用生長瓶(高度12.0 cm,直徑8.0 cm)用高壓滅菌鍋進行滅菌(121℃,26 min)后進行接種。參照《牧草種子檢驗規程GB/T2930.4-2001》[24]挑選干凈的種子,在無菌工作臺中用75%酒精振蕩消毒2 min,無菌水沖洗2次;1% 次氯酸鈉(NaClO)浸泡5 min,無菌水沖洗3~4次,用無菌濾紙瀝干種子表面水分。在無菌工作臺中以200 mL的OD600為1.0左右的DE2菌液為培養基質加入生長瓶中,每種牧草取25粒種子,保持一定距離、整齊的擺放于生長瓶發芽床,設置4個重復,對照用無菌水代替菌液。在溫度23℃、濕度45%、光照+黑暗(18 h+6 h)的組培室進行培養,觀察14 d后取樣。
1.5取樣方法
接種后的14 d,每個生長瓶取10株幼苗測定其生長指標;采集每個處理組和對照組的幼苗整株各1.0 g左右,測定其生理指標,設置3次重復。
1.6 指標的測定及方法
1.6.1測定發芽指標 根據《草種子檢驗規程發芽試驗GB/T2930.4-2017》[25],草木犀種子在種植后第4 d進行初次計數(發芽勢),在第7 d進行末次計數(發芽率);紫花苜蓿種子在種植后第4 d天進行初次計數,在第10 d進行末次計數;沙打旺種子在種植后第4 d進行初次計數,在第14 d進行末次計數。
1.6.2測定生長指標 根長(Root length,RL)和苗長(Seedling length,SL):隨機從各處理取出10株幼苗,測量幼苗由植株最高部到根基部為苗長,根基部到根尖為根長。
鮮重(Fresh weight,FW)和干重(Dry weight,DW):隨機從各處理取出10株幼苗,用吸水紙吸干幼苗表面水分測定其鮮重,用錫紙包裹幼苗放入烘箱,75℃烘48 h至恒重后稱重即為干重。
1.7 數據分析
運用Excel與Origin pro 2021進行數據整理并繪圖,利用統計軟件SPSS 13.0對試驗結果進行分析。
2 結果與分析
2.1 建立DE2標準曲線及確定菌懸液濃度
如圖1所示,以吸光度ABS值為橫坐標,菌懸液濃度為縱坐標繪制的標準曲線,其建立的回歸方程為y=1E+08x-6E+06(R2=0.999 7),標準曲線的回歸系數可達0.99以上,準確度很高。根據曲線可確定后續接種試驗中所采用的DE2菌懸液濃度為108cfu·mL-1時其OD600為1.0。
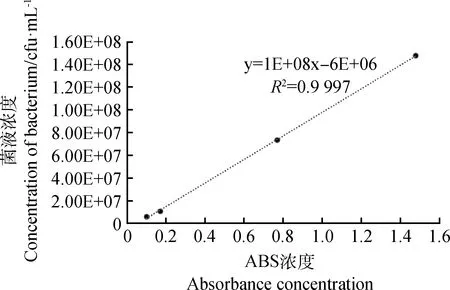
圖1 歐文氏菌DE2菌株的標準曲線
2.2 DE2對3種牧草種子的發芽的影響
由表1所示,沙打旺、草木犀與紫花苜蓿在接種DE2后發芽勢、發芽率和發芽指數相比對照都有降低。其中草木犀分別降低了28.48%,18.99%,28.57%,差異顯著(P<0.05);紫花苜蓿分別降低了22.73%,20.01%,22.51%,差異顯著(P<0.05);沙打旺的發芽勢和發芽率下降不明顯,但發芽指數相比對照顯著降低(P<0.05)。表明DE2抑制了3種牧草種子的發芽。
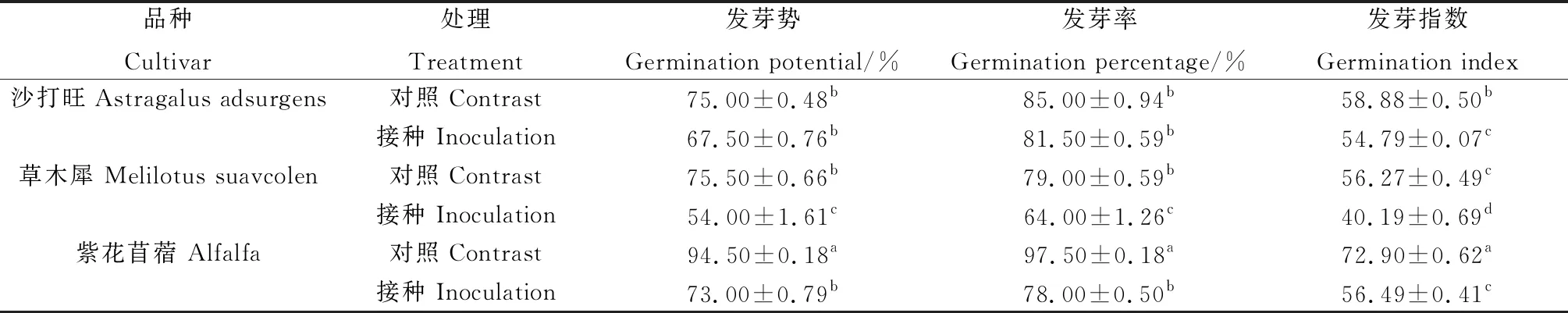
表1 DE2對3種牧草種子萌發的影響
2.3 DE2對3種牧草幼苗生長指標的影響
2.3.1DE2對3種牧草幼苗鮮重、干重、根長和苗長的影響 由圖2所示,接種后的3種牧草相比于對照的鮮重和苗長都無顯著變化,干重相比于對照3種牧草都有減少但是不顯著,根長都明顯降低,分別降低了89.50%,92.69%,93.46%,差異顯著(P<0.05)。表明DE2對3種牧草幼苗的根系生長有明顯抑制作用。
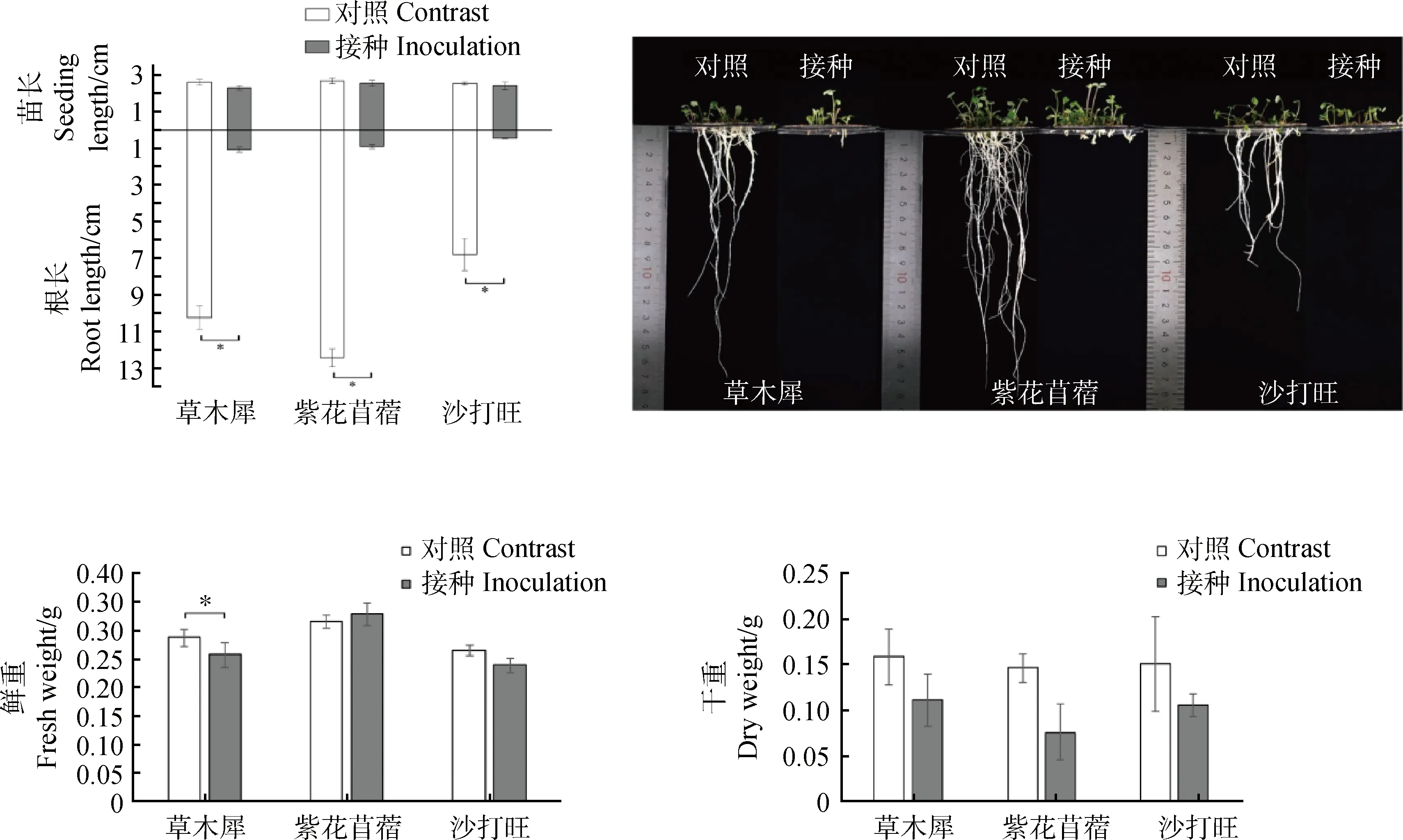
圖2 接種DE2后3種牧草的鮮重、干重、根長和苗長
2.4 DE2對3種牧草幼苗生理指標的影響
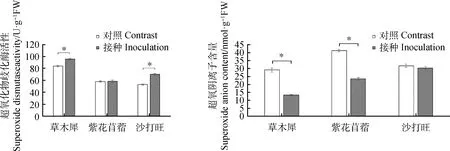
圖3 3種牧草接種DE2后SOD活性與含量的變化
2.4.2DE2對3種牧草CAT與GSH-Px活性與H2O2含量的影響 由圖4所示,草木犀、紫花苜蓿、沙打旺的CAT活性與對照相比明顯增加,分別增長了138.14%,32.36%,73.67%,差異顯著(P<0.05);草木犀、紫花苜蓿的GPX活性相較于對照降低,分別降低了12.55%,26.33%(P<0.05),而沙打旺的GPX活性增加了38.11%(P<0.05);草木犀與沙打旺的H2O2含量相較與對照有所增加,分別增加了43.29%,35.56%,差異顯著(P<0.05),紫花苜蓿的H2O2含量無明顯變化。
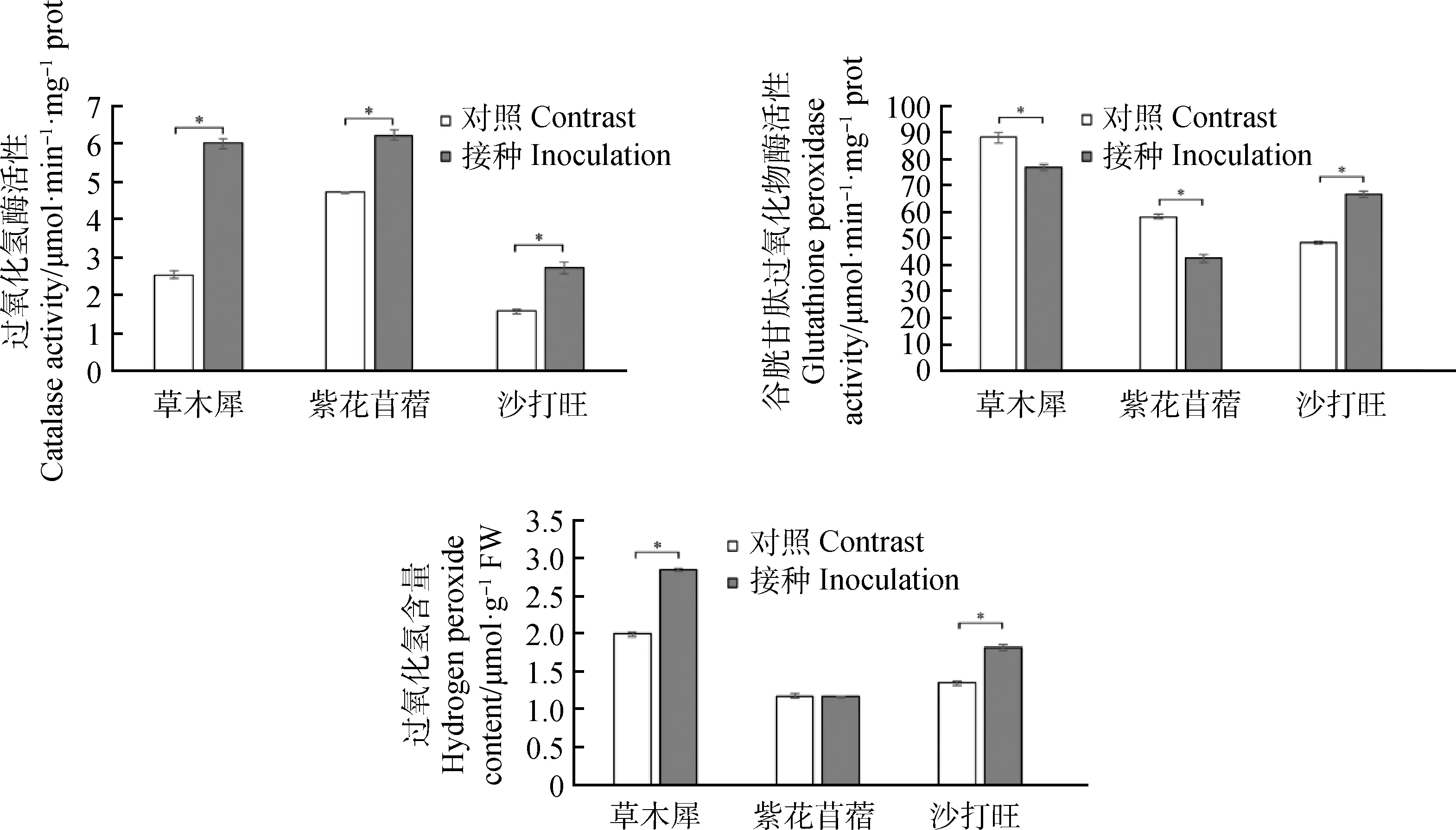
圖4 3種牧草接種DE2后CAT與GSH-Px活性與H2O2含量的變化
2.4.3DE2對3種牧草MDA與可溶性蛋白含量的變化 由圖5所示,接種后草木犀、紫花苜蓿的MDA含量相較于對照均有增加,分別增加了8.65%,6.35%,差異顯著(P<0.05),而沙打旺的MDA含量降低了37.17%(P<0.05);接種后草木犀、紫花苜蓿、沙打旺的可溶性蛋白含量相較與對照組均有所降低,分別降低了12.01%,31.90%,17.12%,差異顯著(P<0.05)。
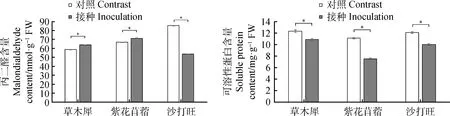
圖5 3種牧草接種DE2后MDA與可溶性蛋白含量的變化
2.5 接種DE2后3種牧草各生理指標的增長率與降低率
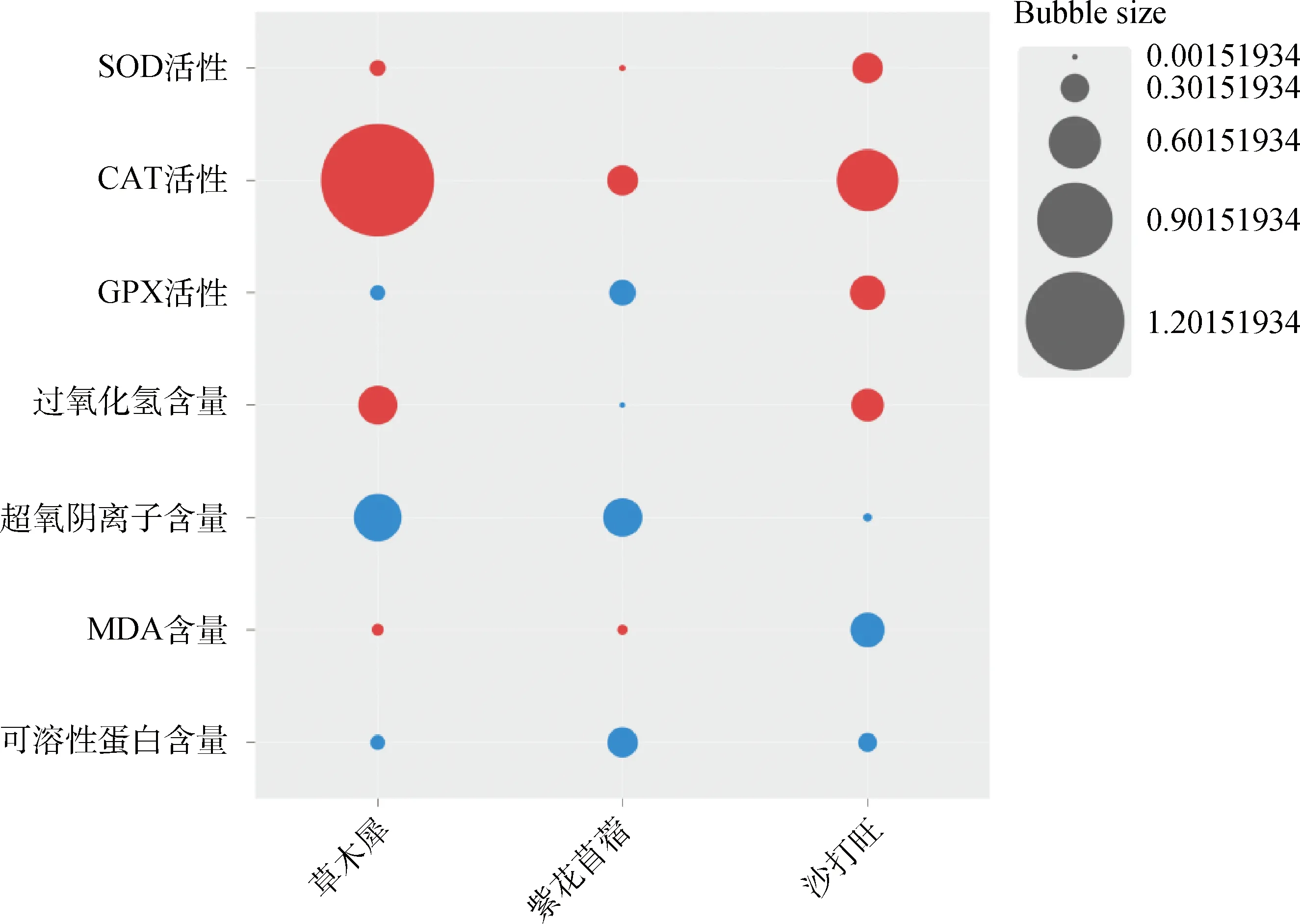
圖6 接種DE2后3種牧草各生理指標的增長率與降低率
2.6 接種DE2后3種牧草幼苗各自指標間的相關性分析
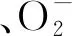
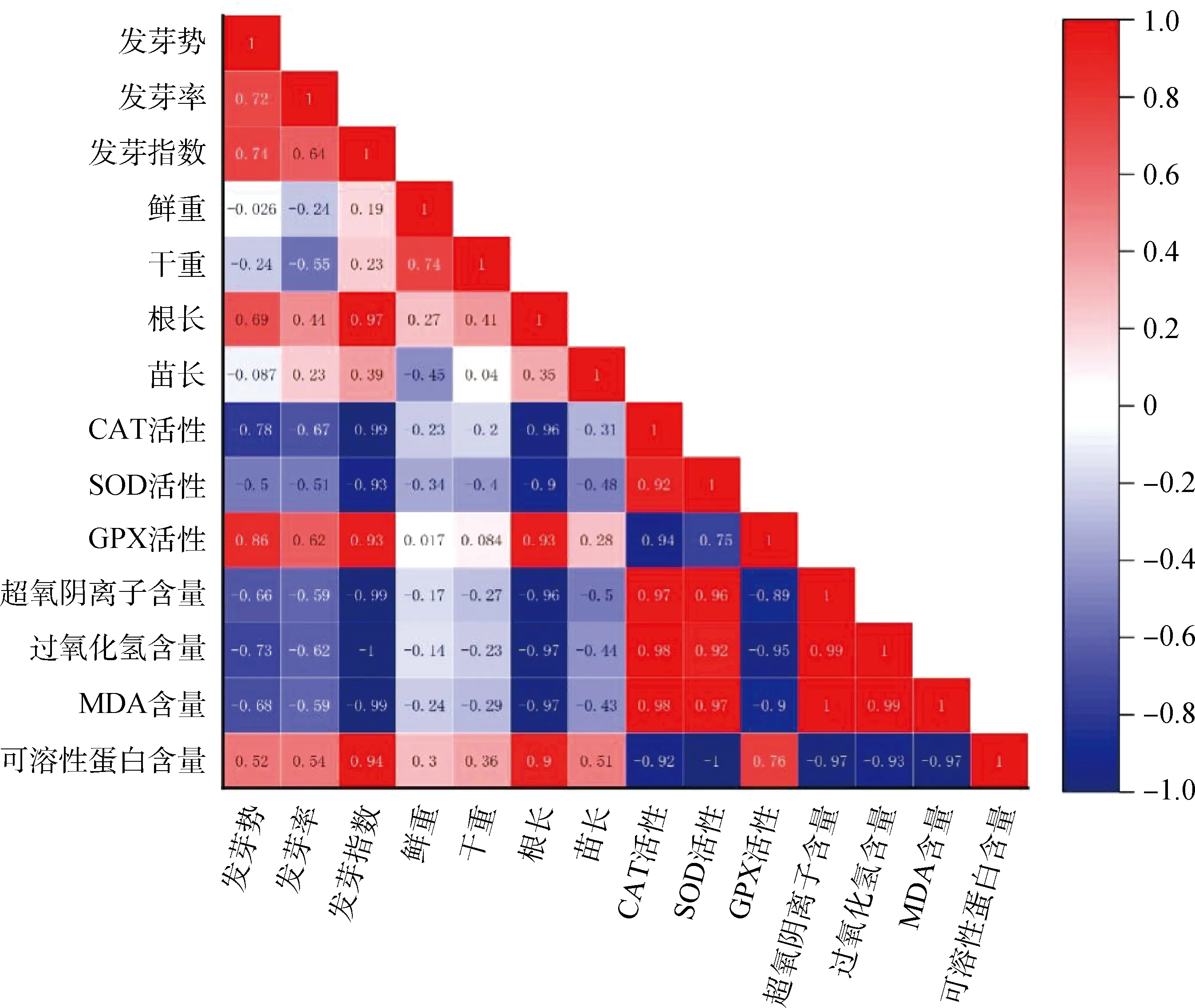
圖7 接種DE2后草木犀各指標間的相關性分析
圖8 接種DE2后紫花苜蓿各指標間的相關性分析
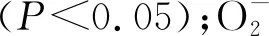
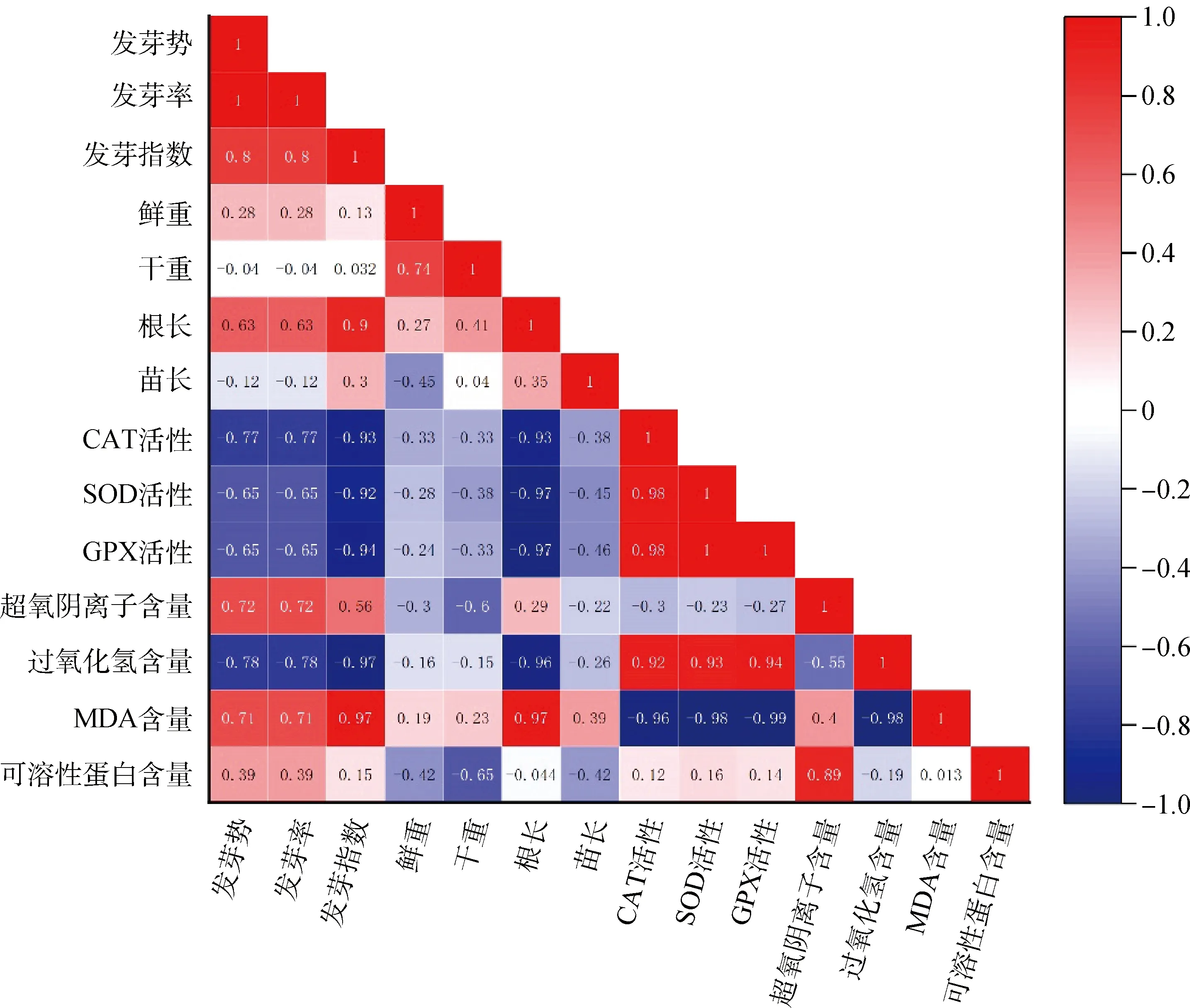
圖9 接種DE2后沙打旺各指標間的相關性分析
3 討論
3.1 DE2對3種豆科牧草幼苗生長的影響
植物病害可影響植株生長的各個階段,研究表明,植物病害可以通過侵染植株葉片影響植株光合和呼吸作用植物病害,還可以通過侵染植物根部和莖部,使植株根、莖腐爛,水分和養分運輸受阻,從而使植株產量下降,品質降低[26-27]。在種子外部的病原菌則會直接侵染種子使其壞死或者產生一些有毒的代謝物質,影響種子的萌發,進而影響植物生長[28]。本試驗表明,接種DE2后3種牧草種子的發芽勢、發芽率、發芽指數均有降低,其中紫花苜蓿與草木犀的各發芽指標顯著降低,與安調過[29]、邢小萍[30]對于黑胚病致病菌會抑制小麥(Triticumaestivum)種子的發芽結果一致。此外本研究還發現接種DE2后各牧草幼苗的鮮重、干重和苗長與對照無顯著差異,但各牧草的根長均顯著低于對照,表明該菌具有從豆科牧草根部侵入的能力,而侵入位點可能位于根毛生長的部位或者根尖區域[31]。還有研究表明,植物病原菌突破根表皮后,首先會對寄主根的皮層區域的細胞造成損傷,且病原菌感染蒺藜苜蓿的根部后造成的主要癥狀便是根生長的停滯[32]。根據汪建軍[33]的報道,接種燕麥(Avenasativa)細菌性葉枯病致病菌后,燕麥根冠比顯著低于對照,這與本試驗結果相一致。說明DE2具有很強的致病性與破壞性,明顯抑制了草木犀、紫花苜蓿與沙打旺種子的萌發和幼苗根系的生長。
3.2 DE2對3種牧草幼苗生理指標的影響
CAT也是生物防御系統的關鍵酶之一,生物學功能是催化細胞內H2O2的分解防止過氧化[40],如陶明福[41]報道檳榔受到黃化病病原菌的侵染后,染病植株體內CAT活性總體增大,增加了對H2O2的降解,從而有利于檳榔植株體內H2O2含量的降低,本研究表明3種豆科牧草接種DE2后CAT活性均顯著增加,而接種后的草木犀與沙打旺的H2O2含量也顯著增加,紫花苜蓿的H2O2含量相比于對照組無明顯變化,說明草木犀與沙打旺幼苗產生的CAT不足以分解來自DE2造成的氧化損失而產生的H2O2,草木犀與沙打旺CAT活性顯著升高可能與其限制病菌擴展能力有關[42]。GPX是一種含巰基的過氧化物酶,可以清除機體內的H2O2、有機氫過氧化物及脂質過氧化物,阻斷ROS自由基對機體的進一步損傷[43]。本試驗中接種后的草木犀與紫花苜蓿的GPX活性相比對照都降低,沙打旺的GPX活性顯著增加而接種后的沙打旺的H2O2濃度也顯著增加,說明沙打旺對DE2的抗性弱,當植物收到脅迫時,多數GPX的表達及活性會增強,但也有表達活性降低的情況[44],例如,在滲透或甲基紫精誘導脅迫條件下,大麥(Hordeumvulgare)的2個GPX同工酶活性增強,而第3個同工酶(HvGPX3)的活性下調[45],雖然在逆境脅迫下,GPX的表達增強,但也有表達下調,因此植物同一物種中的相同亞細胞器來源的不同GPX,或不同亞細胞來源的同一種GPX,其具體的作用機制都需要進一步比較研究[46]。
MDA是膜脂過氧化最重要的產物之一,它的產生還能加劇膜的損傷,因此在植物衰老生理和抗性生理研究中,MDA含量是一個常用指標,可通過MDA了解膜脂過氧化的程度,從而間接測定膜系統受損程度以及植物的抗逆性[47]。可溶性蛋白是重要的滲透調節物質和營養物質,他們的增加和積累能提高細胞的保水能力,對細胞的生命物質及生物膜起到保護作用,因此經常用作篩選抗性的指標之一[48]。本試驗中,接種DE2后草木犀與紫花苜蓿的MDA含量相比于對照顯著增加,說明草木犀與紫花苜蓿在受到DE2侵染時脂膜過氧化而丙二醛不斷地積累,使得蛋白質變性,細胞膜的通透性不斷增大,從而導致電解質大量的外滲,使植物細胞破壞。這與蔣學飛[49]的報道中植株在受到病菌侵害MDA大量增加相一致。本試驗中,接種后的草木犀、紫花苜蓿與沙打旺的可溶性蛋白含量相比對照都有所下降,可能是隨著DE2侵染脅迫的加重,植株體內積累的ROS可以攻擊某些蛋白質的氨基酸殘基,尤其是半胱氨酸(Cys)、酪氨酸(Tyr)、色氨酸(Tpr)、甲硫氨酸(Met)和苯丙氨酸(Phe),使之形成羰基衍生物;ROS還能夠引起分子內和分子間交聯,如形成二硫鍵等,導致體內可溶性蛋白含量有所下降[50]。
4 結論
本試驗確定梨火疫病優勢致病菌歐文氏菌DE2在人工接種條件下,均能危害3種供試豆科牧草的幼苗階段,具體表現為:歐文氏菌DE2抑制草木犀與紫花苜蓿的發芽勢、發芽率、發芽指數,以及沙打旺的發芽指數;接種DE2后3種牧草的根部生長受到的抑制尤其明顯,且不同程度的影響其幼苗生理抗氧化指標。本試驗確定梨火疫病的優勢致病菌對我國主要栽培草地的豆科牧草具有潛在危害,為評估和綜合防控林草系統中新發病原微細菌提供科學基礎。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
