基于渦度系統(tǒng)的干旱區(qū)棗林能量平衡分析
2023-09-07 04:46:56張百和王騰飛
灌溉排水學(xué)報(bào) 2023年8期
喬 英,張百和,邱 凱,王騰飛
(新疆理工學(xué)院,新疆 阿克蘇 843100)
0 引 言
【研究意義】生態(tài)系統(tǒng)中地表能量是陸-氣之間能量和物質(zhì)傳輸、轉(zhuǎn)化的基礎(chǔ),也是生態(tài)系統(tǒng)中一切過程和功能的動力,而地表能量研究的核心問題是以太陽能輸入為驅(qū)動的有機(jī)化學(xué)能循環(huán)及能量交換規(guī)律[1]。所以,深入探究生態(tài)系統(tǒng)的能量循環(huán)規(guī)律,不僅有助于了解區(qū)域氣候系統(tǒng)的能量和物質(zhì)循環(huán)以及氣候變化過程[2],而且還為維持生態(tài)系統(tǒng)的能量和水碳循環(huán)提供重要的科學(xué)依據(jù)[3]。【研究進(jìn)展】能量平衡閉合率(Energy balance ratio,EBR),表示為生態(tài)系統(tǒng)的地表有效能量(潛熱通量LE+顯熱通量H)與可利用能量(凈輻射Rn+土壤熱通量G)的比值[4],國內(nèi)外學(xué)者們深入分析了多種生態(tài)系統(tǒng)的能量平衡,Wilson 等[4]對FLUXNET 的22 個通量站的50 個站點(diǎn)年進(jìn)行能量平衡分析,結(jié)果顯示EBR在53%~99%之間,平均值為79%;李正泉等[5]對ChinaFLUX 的8個通量站進(jìn)行能量平衡分析,結(jié)果顯示EBR平均約為73%。國內(nèi)外學(xué)者們的研究結(jié)果均顯示地表的能量均無法閉合,能量平衡閉合率均在52%~99%之間[4-8],生態(tài)系統(tǒng)種類包括落葉闊葉林[9]、農(nóng)田生態(tài)系統(tǒng)(稻麥輪作)[10]、干旱區(qū)人工檸條林[11]、熱帶季雨林[12]等。
目前,渦度協(xié)方差技術(shù)(Eddy covariance technique,EC)作為監(jiān)測生態(tài)系統(tǒng)能量交換過程的最有效、最準(zhǔn)確的方法,已經(jīng)得到了微氣象學(xué)家和生態(tài)學(xué)家們的認(rèn)可,成為檢驗(yàn)其他測定方法、評價(jià)各類模型模擬精度的標(biāo)準(zhǔn)方法[13]。棗樹因耐旱、喜光照、結(jié)果早,兼顧經(jīng)濟(jì)和生態(tài)效益[14]被成功引進(jìn)新疆,目前種植面積已達(dá)50 萬hm2,成為新疆特色經(jīng)濟(jì)林果業(yè)的支柱[15]。阿克蘇地區(qū)棗樹的種植面積(占全疆34.34%)和產(chǎn)量(占全疆54.70%)均為新疆首位[15]。【切入點(diǎn)】當(dāng)前國內(nèi)外學(xué)者對農(nóng)田生態(tài)系統(tǒng)的研究多為一年生作物,對于人工經(jīng)濟(jì)林的研究相對較少,干旱區(qū)棗林生態(tài)系統(tǒng)的能量平衡分析及各能量分項(xiàng)的變化規(guī)律還需進(jìn)一步探索。
【擬解決的關(guān)鍵問題】綜上,本文以干旱區(qū)人工棗林生態(tài)系統(tǒng)為研究對象,利用渦度相關(guān)系統(tǒng)在田間開展2 a 的觀測試驗(yàn),采用REddyProc 包[16]插補(bǔ)通量缺失數(shù)據(jù),計(jì)算干旱區(qū)人工棗林的熱儲存項(xiàng),評價(jià)棗林能量平衡閉合率,并量化和分析棗林生態(tài)系統(tǒng)的能量變化規(guī)律和能量分配規(guī)律,為干旱區(qū)棗林的水碳循環(huán)研究和優(yōu)化水碳管理模式提供參考。
1 材料與方法
1.1 研究區(qū)概況
研究區(qū)位于新疆阿克蘇市農(nóng)業(yè)示范園(80°18'—80°22'E,41°05'—41°07'N,海拔1 198 m),面積約170 hm2。研究區(qū)氣候干旱少雨屬溫帶沙漠氣候,年均太陽總輻射量5 671.36 MJ/m2,年均日照時間2 911 h,年均降水量68.40 mm,年均氣溫11.20 ℃,年均蒸發(fā)量1 993 mm[17]。研究區(qū)土壤質(zhì)地為砂土,砂粒量87.27%,土壤密度為1.63 g/cm3,田間持水率為8.93%,地下水埋深大于10 m。試驗(yàn)作物為棗樹,品種為灰棗,株行距2 m×4 m,冠層高度為(4±0.2)m[18]。棗樹每年4 月10 日—10 月20 日為生育期,10 月21日—次年4 月9 日為休眠期。
1.2 試驗(yàn)方法
1.2.1 數(shù)據(jù)采集
實(shí)驗(yàn)儀器有渦度相關(guān)系統(tǒng)(IRGASON 型一體機(jī))、四分量輻射計(jì)(CNR4 型)、土壤溫度計(jì)(TCAV 型)、土壤熱通量板(HFP01 型)、土壤水分探頭(Hydra ProbeⅡ型)、數(shù)據(jù)采集器(CR3000 型)[14],詳見表1。

表1 阿克蘇站點(diǎn)實(shí)驗(yàn)儀器匯總Table 1 Summary of experimental instruments at the Aksu site
1.2.2 通量數(shù)據(jù)計(jì)算及數(shù)據(jù)插補(bǔ)質(zhì)量控制
使用軟件LoggerNet 4.5(Campbell Science Inc,USA)按照標(biāo)準(zhǔn)數(shù)據(jù)質(zhì)量控制程序在線計(jì)算[19],并根據(jù)穩(wěn)態(tài)測試、完整湍流特征、超聲風(fēng)速儀的水平定位等,按照微氣象學(xué)經(jīng)驗(yàn)將數(shù)據(jù)質(zhì)量劃分為0~9 級[14]。根據(jù)數(shù)據(jù)質(zhì)量分級,將質(zhì)量差的數(shù)據(jù)刪除并使用R 語言的REddyPrco 包[16]進(jìn)行數(shù)據(jù)插補(bǔ),并采用交叉驗(yàn)證法來評價(jià)插補(bǔ)數(shù)據(jù)的質(zhì)量[14];同時,使用能量閉合率來檢驗(yàn)插補(bǔ)數(shù)據(jù)質(zhì)量。
選擇2018 年和2019 年的30 min 尺度數(shù)據(jù),采用Kljun 等[20]和Kormann 等[21]的源區(qū)模型計(jì)算阿克蘇站點(diǎn)的通量貢獻(xiàn)區(qū),90%的貢獻(xiàn)區(qū)(通量源區(qū))的圓形半徑分別為356.50、350.00 m。
1.2.3 能量平衡計(jì)算
根據(jù)能量守恒定律,生態(tài)系統(tǒng)能量平衡可用式(1)表示[22],其中的熱儲存項(xiàng)S可用式(2)表示[23],熱儲存項(xiàng)的計(jì)算方法參見Dou 等[24]文獻(xiàn)計(jì)算。能量平衡閉合率(Energy balance ratio,EBR)是指地表有效能量(潛熱通量LE+顯熱通量H)與可利用能量(凈輻射Rn-土壤熱通量G)之比[25],EBR可用式(3)計(jì)算[26]。為了提高能量平衡閉合率,很多研究也將熱儲存項(xiàng)S記入可利用能量中,EBR可用式(4)計(jì)算[26]。
式中:Rn為太陽凈輻射(W/m2);G為土壤熱通量(W/m2);H為顯熱通量(W/m2);LE為潛熱通量(W/m2);S為熱儲存項(xiàng)(W/m2);Q為其他來源的能量(忽略不計(jì),W/m2);Ss為一定深度范圍內(nèi)的土壤熱儲存項(xiàng)(W/m2);Sa為大氣顯熱儲存項(xiàng)(W/m2);Se為大氣潛熱儲存項(xiàng)(W/m2);Sc為冠層熱儲存項(xiàng)(W/m2);Sp為植物光合儲存項(xiàng)(W/m2);St為樹木枝干熱儲存項(xiàng)(W/m2)[23]。
2 結(jié)果與分析
2.1 氣溫、降水量和灌溉量的變化情況
觀測期(2018、2019 年)日平均氣溫、降水量、灌溉量如圖1 所示。觀測期間,日平均氣溫在-12.72~29.16 ℃之間變化,年平均氣溫為10.99 ℃,其中月平均氣溫最低出現(xiàn)在2018 年12 月(-8.36 ℃)和2019 年1 月(-8.45 ℃),月平均氣溫最高出現(xiàn)在2018 年7 月(24.07 ℃)和2019 年7 月(24.74 ℃),6—8 月的月平均氣溫在20 ℃以上。降水集中在4—10 月(生育期)內(nèi),2018、2019 年年降水量分別為81.50、57.40 mm,單日最大降水量為13.20 mm。灌溉集中在4—10 月,2018、2019 年灌溉量分別為847.15 mm(灌溉31 次)、848.25 mm(灌溉30 次)。
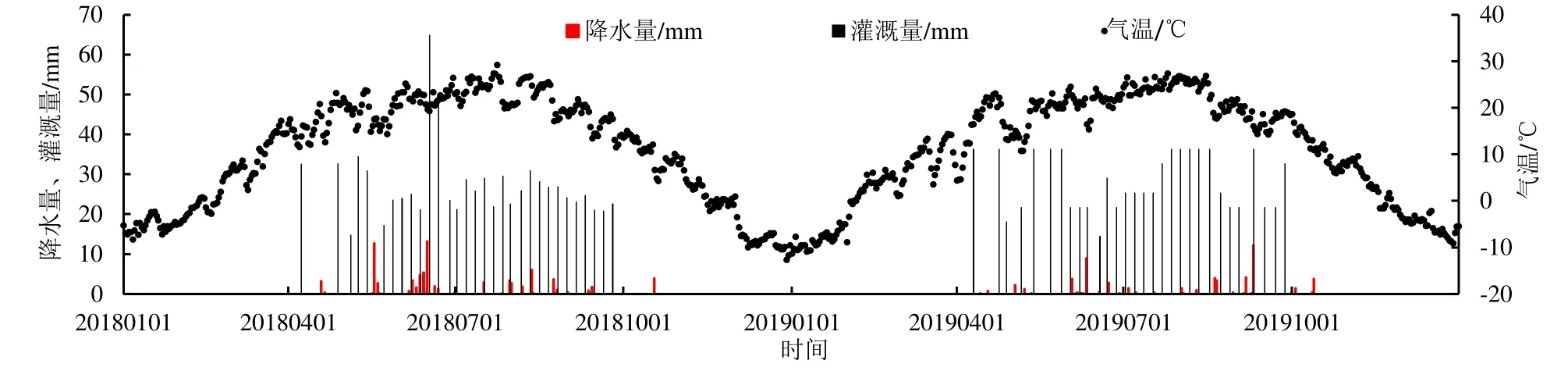
圖1 研究區(qū)2018年和2019年氣溫、降水量及灌溉量Fig.1 Distribution of temperature, precipitation and irrigation of the study area in 2018 and 2019
2.2 能量通量值
使用REddyPrco 包插補(bǔ)后的通量數(shù)據(jù)及式(2)計(jì)算了干旱區(qū)棗林的能量分項(xiàng),詳見表2。由表2 可知,2018、2019年凈輻射Rn年度合計(jì)為1 475 227.47、1 481 026.93 W/m2,在生育期明顯大于休眠期;2018、2019 年潛熱通量LE年度合計(jì)為898 349.68、922 381.08 W/m2,潛熱通量主要集中在棗樹生育期,在休眠期較小;2018、2019年顯熱通量H年度合計(jì)為205 778.37、183 786.36 W/m2,主要集中在休眠期,在生育期較小;2018、2019 年土壤熱通量G年度合計(jì)為-15 839.17、-13 877.22 W/m2,在生育期為正,休眠期為負(fù);且Rn、LE、H、G在2018 年和2019 年情況相似。由表2 可知,顯熱熱儲存項(xiàng)Sa為S中的最大項(xiàng),2018、2019年Sa年度合計(jì)分別為22 053.05、17 847.04 W/m2,占凈輻射Rn的1.49%、1.21%;其次是植物光合熱儲能Sp,2018、2019 年Sp年度合計(jì)分別為13 153.85、11 345.06 W/m2,占凈輻射Rn的0.89%、0.77%;土壤熱儲存項(xiàng)Ss、冠層熱儲存項(xiàng)Sc、潛熱熱儲存項(xiàng)Se在2018 年和2019 年的年度合計(jì)均較小。
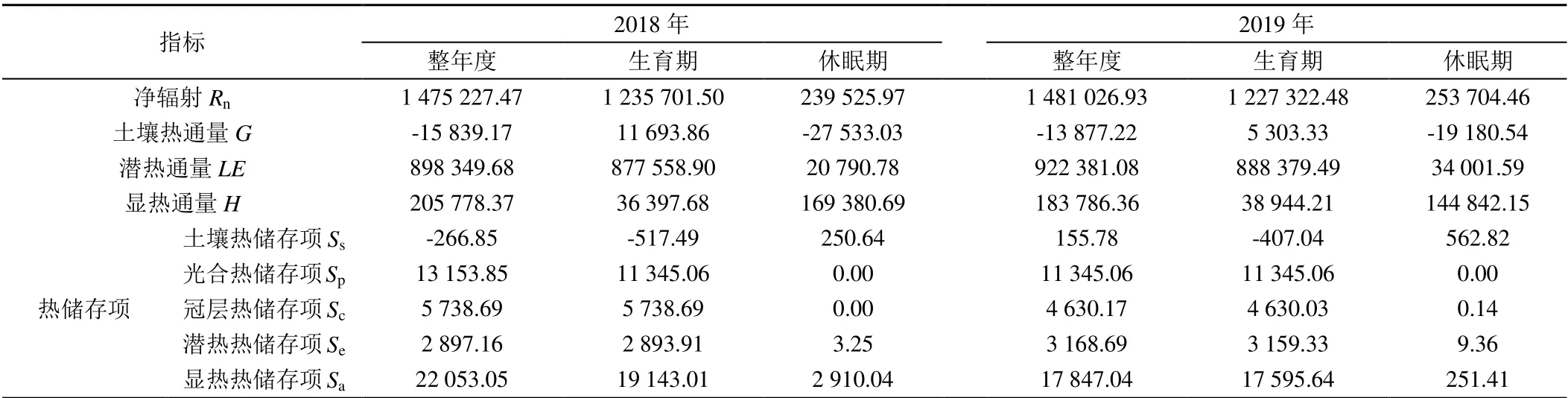
表2 干旱區(qū)棗林的能量分項(xiàng)Table 2 Energy subdivision of date jujube forests in arid areas W/m2
2.3 能量通量的月平均日變化特征
2018、2019 年REddyProc 插補(bǔ)后的能量通量(凈輻射Rn、潛熱通量LE、顯熱通量H、土壤熱通量G)的月平均日變化規(guī)律見圖2。由圖2 可知,Rn、LE、H、G的月平均日變化均呈單峰變化趨勢,2018 年和2019 年各月的能量變化趨勢相似。Rn、LE、H、G在白天為正,夜間為負(fù),正負(fù)轉(zhuǎn)換的交點(diǎn)與白晝變化的時間一致。各月Rn的最小值均為負(fù),在-74.16~-47.51 W/m2之間波動;Rn在夜間為負(fù)值,在白天為正值,說明在夜間地表損失熱量,而在白天吸收熱量。2018年Rn的最大值為653.05 W/m2,出現(xiàn)在7 月;2019年Rn的最大值為638.98 W/m2,出現(xiàn)在6 月;2 a 最大值均出現(xiàn)在15:30。2018 年和2019 年LE的最大值均出現(xiàn)在8 月,最大值分別為331.85、290.20 W/m2,且最大值出現(xiàn)時間和Rn一致,均在15:30。LE在生育期明顯大于休眠期,在5—9 月,LE為Rn的主要分配項(xiàng)。H在2018 年和2019 年的變化幅度相似,在-80.51~180.45 W/m2之間波動,2018 年H的最大值為180.45 W/m2,出現(xiàn)在4 月;2019 年H的最大值為165.06 W/m2,出現(xiàn)在5 月。H在1—4 月和10—12月為Rn的主要分配項(xiàng)。G在2018 年和2019 年的變化幅度相似,在-24.09~71.16 W/m2之間波動,2018年G的最大值為34.52 W/m2,出現(xiàn)在4 月;2019 年G的最大值為71.16 W/m2,出現(xiàn)在3 月;G占Rn的比值較小。

圖2 2018、2019年干旱區(qū)棗林1—12月能量的月平均日變化規(guī)律Fig.2 Monthly average daily variation of energy in jujube forests in arid areas from January to December 2018 and 2019
2.4 干旱區(qū)棗林生態(tài)系統(tǒng)的能量平衡分析
2018、2019 年干旱區(qū)棗林生態(tài)系統(tǒng)各月能量平衡閉合線性回歸參數(shù)和能量平衡閉合率情況見表3。表3 顯示,未計(jì)入熱儲存項(xiàng)時,各月的有效能量和可利用能量之間的統(tǒng)計(jì)回歸斜率k為0.32~0.69,2018年k值最低為0.37,出現(xiàn)在1 月,2019 年k值最低為0.32,出現(xiàn)在12 月;各月能量平衡閉合線性回歸參數(shù)決定系數(shù)R2為0.60~0.92;斜率k和R2在3—10 月之間變化較平緩,極小值出現(xiàn)在11—12 月、1—2 月。各月的能量平衡閉合率在34.05%~80.78%之間,閉合率最高為80.78%(2019 年9 月);閉合率最低為34.05%(2018 年12 月);未計(jì)入熱儲存項(xiàng)時,表明各月的能量平衡閉合程度較好。由于受太陽高度、棗林生育期中地表覆蓋度和葉面積指數(shù)、灌溉、氣象等因素變化的影響,各月的線性回歸斜率和能量平衡閉合比率存在一定的波動。計(jì)入熱儲存項(xiàng)后,各月的回歸斜率k、R2、能量平衡閉合率均有所提高,但增加幅度較小。表3 顯示,棗樹生育期的能量平衡閉合率明顯高于休眠期,同時線性回歸斜率和R2也是生育期明顯大于休眠期;計(jì)入熱存儲項(xiàng)后,k、R2、EBR也均有提高。
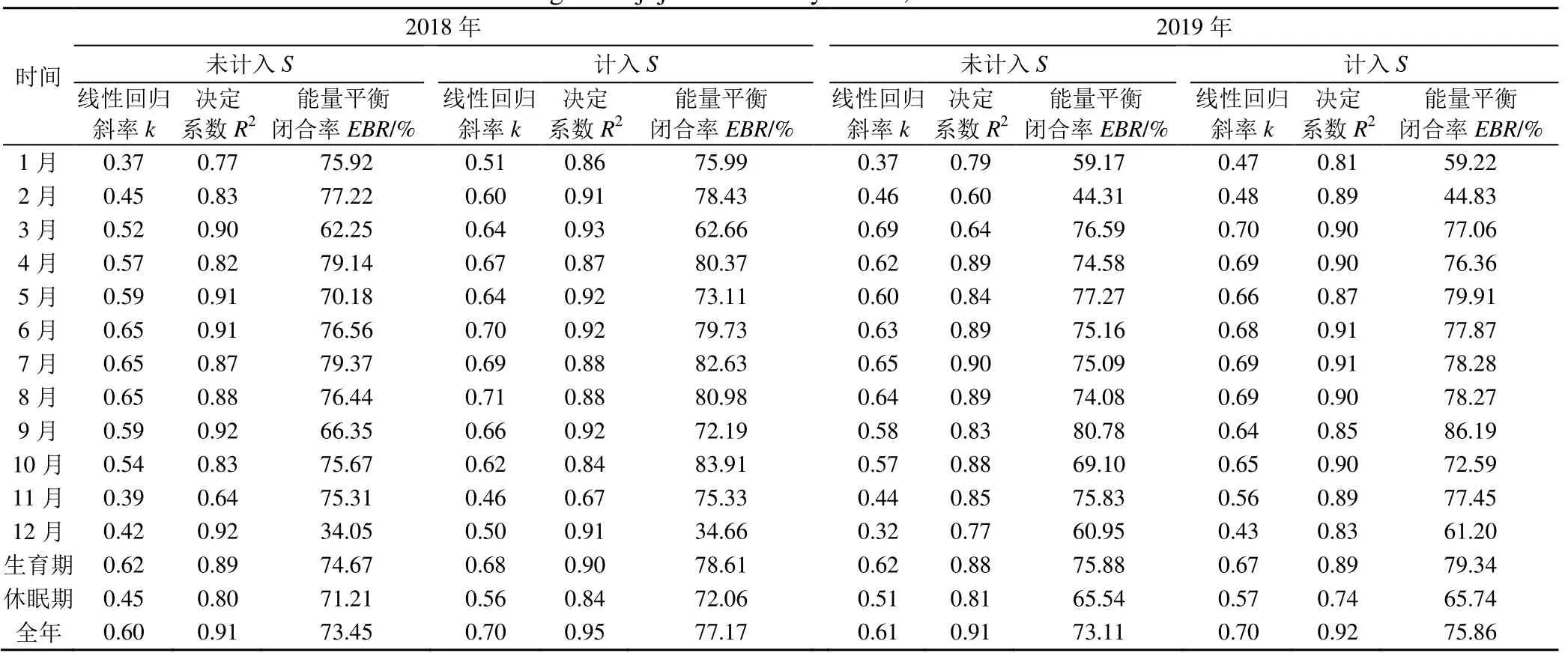
表3 2018、2019年干旱區(qū)棗林各月能量平衡閉合線性回歸參數(shù)和能量平衡閉合率Table 3 Energy balance closure linear regression parameters and energy balance closure rates in arid region of jujube forests by month, 2018 and 2019
干旱區(qū)棗林生態(tài)系統(tǒng)30 min 尺度的能量平衡如圖3 所示。由圖3 和表3 可知,未計(jì)入熱儲存項(xiàng)的干旱區(qū)棗林生態(tài)系統(tǒng)2018、2019 年30 min 尺度上的有效能量(H+LE)與可利用能量(Rn-G)的直線擬合斜率為0.60(R2=0.91)和0.61(R2=0.91),能量平衡閉合率分別為73.45%、73.11%。將熱儲存項(xiàng)計(jì)入可利用能量,2018 年和2019 年(H+LE)與(Rn-G-S)的直線擬合斜率為0.70(R2=0.95)和0.70(R2=0.92),記入熱儲存項(xiàng)后能量平衡閉合率有所提高,干旱區(qū)棗林2018 年和2019 年的能量平衡閉合率分別為77.17%、75.86%,且2 a 的閉合率相似。學(xué)者們[5-10]認(rèn)為生態(tài)系統(tǒng)能量平衡不閉合程度在10%~30%之間,本文研究結(jié)果在此范圍內(nèi),說明渦度相關(guān)法在干旱區(qū)棗林生態(tài)系統(tǒng)通量觀測中可靠性較高,下墊面符合觀測范圍;同時,也證明REddyPrco 包插補(bǔ)的通量數(shù)據(jù)質(zhì)量較好。

圖3 干旱區(qū)棗林生態(tài)系統(tǒng)30 min尺度的能量平衡Fig.3 Energy balance of date Jujube forests ecosystems in arid areas at the half-hourly scale
2.5 生育期和休眠期的熱儲存項(xiàng)分項(xiàng)分析
土壤熱儲存項(xiàng)Ss、顯熱熱儲存項(xiàng)Sa、潛熱熱儲存項(xiàng)Se、冠層熱儲存項(xiàng)Sc和光合熱儲存項(xiàng)Sp在棗樹生育期和休眠期的日平均變化見圖4。

圖4 棗林的能量分項(xiàng)在生育期和休眠期的日平均變化Fig.4 Average daily changes in the energy subdivision of date Jujube forests during the reproductive and dormant periods
圖4(a)顯示,2018 年和2019 年土壤熱儲存項(xiàng)Ss日平均變化在生育期和休眠期規(guī)律相似,差異不大,均在-51.17~69.15 W/m2之間呈單峰變化,與土壤熱通量變化規(guī)律相似。Ss在白天為正值,表示為吸收熱量;而在夜間為負(fù)值,表示為釋放熱量。圖4(b)顯示,2018 年和2019 年潛熱熱儲存項(xiàng)Se日平均變化規(guī)律在生育期和休眠期不同:在休眠期潛熱熱儲存項(xiàng)的變化范圍在0 附近,變化幅度較小,為±0.05 W/m2,呈單峰變化;在生育期潛熱熱儲存項(xiàng)在-0.23~0.58 W/m2之間呈雙峰變化規(guī)律,在09:00 和20:00 均有峰值,說明在晝夜交替的時候潛熱熱儲存項(xiàng)變化較大。光合熱儲存項(xiàng)Sp和冠層熱儲存項(xiàng)Sc在休眠期均為0,所以圖4(c)只繪制了生育期的光合熱儲存項(xiàng)和冠層熱儲存項(xiàng)的平均日變化。圖4(c)顯示,光合熱儲存項(xiàng)Sp呈單峰變化規(guī)律,在夜間為0,處于0~2.95 W/m2之間,Sp隨著凈輻射的變化而變化,當(dāng)凈輻射在15:00達(dá)到峰值時,Sp繼續(xù)升高在18:00 左右達(dá)到峰值,并隨著凈輻射的減小而迅速降低。2 a 的冠層熱存儲項(xiàng)Sc均較小,在-0.39~1.55 W/m2之間呈單峰型變化。Sc在夜間變化平緩,變化幅度小;白天隨著凈輻射的增大而增大,到達(dá)峰值時間與凈輻射一致。圖4(d)顯示,2018 年和2019 年顯熱熱儲存項(xiàng)Sa在生育期和休眠期變化規(guī)律不同。生育期的顯熱熱儲存項(xiàng)在-0.80~6.50 W/m2之間呈單峰變化;Sa在夜間變化平緩幅度小,白天隨著凈輻射的增大而迅速變大,在12:00左右達(dá)到峰值,峰值持續(xù)6 h 左右,在18:00 之后緩慢降低。休眠期的顯熱熱儲存項(xiàng)在-2.74~3.35 W/m2之間變化,其變化幅度比生育期小,在夜間為負(fù),白天為正;日出后,隨著凈輻射迅速升高,Sa也迅速增大,在12:00 左右達(dá)到峰值,然后逐漸減小。
3 討 論
3.1 棗林生態(tài)系統(tǒng)的能量分配特征
在太陽輻射能量的驅(qū)動下,農(nóng)田生態(tài)系統(tǒng)內(nèi)進(jìn)行能量流動、物質(zhì)合成轉(zhuǎn)移和水碳循環(huán)等運(yùn)動過程。因下墊面的作物類型、種植密度、生育期、季節(jié)、晝夜、氣象、人為干擾等差異,會導(dǎo)致凈輻射進(jìn)入生態(tài)系統(tǒng)后,各個能量分項(xiàng)等在系統(tǒng)內(nèi)的分配變化存在差異[27]。同時,生態(tài)系統(tǒng)的能量分配變化規(guī)律也可以反映下墊面的干濕狀況[28]。
5—10 月,干旱區(qū)棗林生態(tài)系統(tǒng)的LE/Rn的比值大于50%,LE/Rn的比值最大出現(xiàn)在2018 年8 月(92.10%)和2019 年9 月(96.43%)。在4 月和11月,LE/Rn比值在17.33%~35.18%之間,在1—3 月和12 月,LE/Rn的占比均較小,在3.35%~12.25%之間。分析其原因,本文研究區(qū)是干旱區(qū),年降水量較小,作物生長依賴于灌溉,棗林只在生育期進(jìn)行灌溉,時間為每年的4—10 月,灌溉后,土壤濕潤,土壤蒸發(fā)較大,所以5—10 月干旱區(qū)棗林LE/Rn的占比大于50%,說明凈輻射主要用于下墊面的LE,即蒸散(包括植株蒸騰和土壤蒸發(fā))。而4 月的LE/Rn為27.23%和33.41%,是因?yàn)? 月棗林雖然進(jìn)行了灌溉,土壤濕潤導(dǎo)致土壤蒸發(fā)較大,但是棗樹處于萌芽期,葉面積指數(shù)較小導(dǎo)致植株蒸騰較小,綜合起來表現(xiàn)為LE/Rn比3 月有明顯增加(增幅為21.57%和21.16%)。LE/Rn在8—9月最大,處于86.47%~96.43%之間,此時也是棗林灌溉量最大、葉面積指數(shù)最大的時期。這表明在棗林的生育期(5—10 月)能量分配由LE占據(jù)主導(dǎo)。
在1—4 月和11—12 月,棗林生態(tài)系統(tǒng)中的H為Rn的最大支出項(xiàng);在1—3 月和11、12 月,H/Rn占比均大于50%,在4 月H/Rn占比接近50%(2018 年為47.59%,2019 年為45.19%),這表明,在棗樹休眠期,能量分配由H占據(jù)主導(dǎo)。Trenberth 等[29]分析了全球能量收支,認(rèn)為地表吸收的能量有近1/2 消耗在潛熱,同時,能量分配方式與生態(tài)系統(tǒng)中的水分平衡受大氣水分供需影響。也有學(xué)者[2,6,9]進(jìn)一步研究表明,植被覆蓋較好的生態(tài)系統(tǒng)(農(nóng)田、森林和草甸等)能量分配由LE占據(jù)主導(dǎo),這也驗(yàn)證了本文的研究結(jié)果,對于干旱區(qū)的棗林生態(tài)系統(tǒng)能量分配來說,水分條件是決定能量分配的關(guān)鍵因素,若生態(tài)系統(tǒng)水分充足,能量分配則以潛熱通量為主,反之則以感熱通量為主[30-31]。干旱區(qū)棗林的能量分配方案由水分條件決定,受到水分條件、植被類型和氣候特征的影響[32]。
3.2 能量平衡閉合率
干旱區(qū)棗林生態(tài)系統(tǒng)在2018 年和2019 年的全年能量平衡閉合率為73.45%、73.11%,這與前人[4-12]在其他的生態(tài)系統(tǒng)中觀測的能量平衡閉合率相似,在合理的范圍內(nèi)(能量平衡不閉合率處于10%~30%之間)。根據(jù)Kljun 等[20]和Kormann 等[21]的源區(qū)模型計(jì)算出的下墊面90%通量貢獻(xiàn)區(qū)的圓形半徑分別為356.50、350.00 m,而實(shí)際試驗(yàn)區(qū)的面積覆蓋90%通量貢獻(xiàn)區(qū);試驗(yàn)區(qū)的下墊面范圍是合適的,渦度儀器的觀測高度也是合適的,這說明在干旱區(qū)棗林使用渦度相關(guān)觀測系統(tǒng)測量能量是可行的,得到的觀測數(shù)據(jù)可信度高。
干旱區(qū)棗林生態(tài)系統(tǒng)的全年能量平衡閉合程度不高,這可能是由于忽略了熱存儲項(xiàng),于是本文利用式(4)重新計(jì)算了能量平衡閉合率,結(jié)果顯示記入熱存儲項(xiàng)后,2018 年和2019 年的EBR提高了3.72%、2.75%,達(dá)到了77.17%、75.86%,與美國溫帶落葉林[33]、加拿大北部楊樹林[34]、西雙版納熱帶季節(jié)雨林[24]的增幅范圍相似;但是比黃土高原檸條林[23]、黑河中游生育期的春小麥農(nóng)田[22]等的增幅低。多位學(xué)者指出[13,22-23],下墊面土壤熱儲量Ss為熱儲存項(xiàng)中的最大占比,忽略Ss是造成地表能量平衡不閉合的主要原因。但是根據(jù)表2 顯示,Ss占凈輻射的比例小于1%,這與岳平等[2]、高紅貝等[22]研究結(jié)果不一致。這可能是因?yàn)楸疚难芯繀^(qū)處于干旱區(qū),作物需要灌溉,棗林使用深井水進(jìn)行滴灌,井水溫度低于灌溉時的氣溫和土壤溫度,導(dǎo)致棗林每次灌溉時降低了土壤熱儲量Ss,故本文雖然計(jì)算了Ss,但是EBR提高幅度小,Ss占凈輻射的比例小于1%。
4 結(jié) 論
1)Rn、LE、H、G的日均值均呈單峰變化趨勢,2018 年和2019 年的變化趨勢相似。
2)未計(jì)入熱儲存項(xiàng)時,各月的有效能量和可利用能量之間的統(tǒng)計(jì)回歸斜率為0.32~0.70,決定系數(shù)R2為0.64~0.94,能量平衡閉合率在34.05%~80.78%之間;計(jì)入熱儲存項(xiàng)后,各月的有效能量和可利用能量之間的統(tǒng)計(jì)回歸斜率、決定系數(shù)、能量平衡閉合率均有所提高,但增幅較小。
3)2018 年和2019 年未計(jì)入熱儲存項(xiàng)的全年能量平衡閉合率分別為73.45%、73.11%;計(jì)入熱儲存項(xiàng)后,能量平衡閉合率有所提高,但增幅較小。
4)干旱區(qū)棗林生育期的能量平衡閉合率高于休眠期;計(jì)入熱存儲項(xiàng)后,生育期能量平衡閉合率的增幅也明顯大于休眠期。
5)土壤熱儲存項(xiàng)的日平均變化在生育期和休眠期規(guī)律相似;潛熱熱儲存項(xiàng)的日平均變化在休眠期呈單峰變化,生育期呈雙峰變化;光合熱儲存項(xiàng)在生育期呈單峰變化;冠層熱存儲項(xiàng)在生育期呈單峰變化;顯熱熱儲存項(xiàng)在生育期呈單峰變化,休眠期波動幅度較小。
6)干旱區(qū)棗林的能量分配受水分條件、植被類型和氣候特征的影響,且能量分配方案由水分條件決定,若生態(tài)系統(tǒng)水分充足,能量分配則以潛熱通量為主,反之則以感熱通量為主。
(作者聲明本文無實(shí)際或潛在的利益沖突)
