牛蛙源米爾伊麗莎白菌的分離鑒定與藥敏試驗(yàn)
2023-09-06 08:48:50向圣漢趙雨賀彭治鑫李俊慧李偉
水產(chǎn)學(xué)雜志 2023年4期
關(guān)鍵詞:耐藥
向圣漢,趙雨賀,彭治鑫,李俊慧,李偉
(長(zhǎng)江大學(xué)生命科學(xué)學(xué)院,湖北 荊州 434000)
隨著蛙類(lèi)養(yǎng)殖業(yè)的興起,蛙類(lèi)病害問(wèn)題也日漸突出。蛙類(lèi)病害主要有真菌性疾病[1]、細(xì)菌性疾病[2]、寄生蟲(chóng)疾病[3],環(huán)境因素也可造成病害發(fā)生[4]。在人工養(yǎng)殖環(huán)境下,細(xì)菌引起蛙類(lèi)的多種重要疾病。目前已報(bào)道的致病菌如腦膜膿毒性黃桿菌(Flavobacterium menigosepticum)可引起牛蛙(Rana catesbeiana)歪脖子病[5],氣單胞菌(Aeromonas sp.)可引起牛蛙紅腿病[6]、敗血癥[7]、腹水病[8]、白內(nèi)障[9]等多種疾病。不同致病菌可能會(huì)導(dǎo)致類(lèi)似的病癥,如遲緩愛(ài)德華氏菌(Edwardsiella tarda)[10]、嗜水氣單胞菌(Aeromonas hydrophila)[7]、點(diǎn)狀產(chǎn)氣單胞菌(Aeromonas punctata)、非霍亂弧菌(Noncholera vibrios)[11]等都可引起蛙類(lèi)敗血癥。
米爾伊麗莎白菌(Elizabethkingia miricola)是伊麗莎白菌屬的重要物種,最早分離于Mir 空間站的冷凝水[12]。最近,有相關(guān)報(bào)道表明,感染米爾伊麗莎白菌的蛙會(huì)出現(xiàn)神經(jīng)系統(tǒng)疾病,造成中樞神經(jīng)系統(tǒng)損傷和類(lèi)似于腦膜炎的癥狀[13]。在多種無(wú)尾目?jī)蓷珓?dòng)物中報(bào)道了米爾伊麗莎白菌感染,導(dǎo)致病蛙出現(xiàn)歪頭和白內(nèi)障等病癥。胡瑞雪等[14]和秦振陽(yáng)等[15]從患歪頭病黑斑蛙(Pelophylax nigromaculatus);Lei等[16]從患有歪頭、白內(nèi)障病癥的棘胸蛙(Quasipaa spinosa)中分離鑒定的致病病原均為米爾伊麗莎白菌。國(guó)外在林蛙(Rana sylvatica)、雨蛙(Hyla viridis)中也有被米爾伊麗莎白菌感染出現(xiàn)歪頭癥病癥的報(bào)道[17]。
針對(duì)牛蛙歪頭、白內(nèi)障病致病菌的研究歷史悠久。陳耀明等[18]首次報(bào)道了腦膜膿毒性黃桿菌會(huì)引起病蛙歪頭和白內(nèi)障病癥。陳會(huì)波等[19]報(bào)道牛蛙歪頭白內(nèi)障病原是黃桿菌IIb 群(Plavobacterium group IIb)。楊春浩等[20,21]發(fā)現(xiàn)嗜水氣單胞菌、藤黃葡萄球菌(Micrococcus luteus)和淺黃假單胞菌(Pseudomonas mendocina)都存在于患有歪頭白內(nèi)障病癥的牛蛙組織,可能都參與了致病作用。有鑒于此,分離并鑒定牛蛙歪頭白內(nèi)障病病原對(duì)于該疾病的預(yù)防和控制至關(guān)重要。2021 年上半年,湖北荊州江陵縣牛蛙養(yǎng)殖場(chǎng)出現(xiàn)大量歪頭、白內(nèi)障癥狀病蛙,通過(guò)分離病原菌、生理生化鑒定、基因序列分析等方法鑒定病原并分析其致病性和耐藥性,為該病害的治療和防治提供了參考資料。
1 材料與方法
1.1 供試牛蛙及主要試劑
試驗(yàn)牛蛙取自湖北省荊州江陵縣牛蛙養(yǎng)殖基地。
抗菌藥物藥敏紙片(批號(hào):20210316)、細(xì)菌微量生化反應(yīng)管(批號(hào):20210422)(杭州微生物試劑有限公司);瓊脂糖凝膠DNA 回收試劑盒(北京全式金生物技術(shù)有限公司);細(xì)菌基因組DNA 提取試劑盒(北京天根生化科技有限公司)。
1.2 方法
1.2.1 病原菌的分離、純化和鑒定
患歪頭、白內(nèi)障癥的牛蛙用70%的酒精進(jìn)行表面消毒后,在實(shí)驗(yàn)室無(wú)菌條件下解剖。用接種環(huán)將眼睛和大腦分離的樣本接種到牛肉膏蛋白胨培養(yǎng)基上,于30℃恒溫培養(yǎng)箱中培養(yǎng)24h,挑取培養(yǎng)基上長(zhǎng)勢(shì)良好的菌落進(jìn)行純化培養(yǎng),并保存?zhèn)溆谩H』罨w涂片,用革蘭氏染色,觀察形態(tài)特征。根據(jù)生化微量反應(yīng)管說(shuō)明進(jìn)行生化鑒定。
提取分離優(yōu)勢(shì)菌基因組DNA,對(duì)菌株進(jìn)行16S rRNA 基因和gyrB 基因PCR 擴(kuò)增。16S rRNA 基因上游引物F 為:5’-AGAGTTTTGATCCTGGCTCAG-3’;下游引物R為:5’-AAGGAGGTGATCCAGCCGCA-3’;gyrB 基因上游引物F 為:5’-GAAGTCATCATGACCGT TCTGCAYGCNGGNGGNAARTTYGA3’;下游引物R 為:5’-AGCAGGGTACGGATGTGCGAGCCRTC NACRTCNGCRTCNGTCAT-3’。反應(yīng)體系為(50 μL):2×PCR Mix 25 μL,上下游引物各1.0 μL,DNA 模板2.0 μL,ddH2O 21 μL。反應(yīng)條件為:95 ℃預(yù)變性3 min,95 ℃變性30 s,53 ℃退火30 s,72℃延伸60 s,循環(huán)34 次,72 ℃延伸7 min。PCR 產(chǎn)物經(jīng)1.0%瓊脂糖凝膠電泳檢測(cè)后回收,送生工生物工程(上海)股份有限公司測(cè)序。測(cè)序得到的基因序列在NCBI中進(jìn)行BLAST 分析比對(duì),從數(shù)據(jù)庫(kù)獲得相關(guān)屬種的16S rRNA 和gyrB 基因序列,找到相似性較高的菌株應(yīng)用MEGA7.0 軟件構(gòu)建系統(tǒng)發(fā)育進(jìn)化樹(shù)。
1.2.2 藥敏試驗(yàn)
藥敏試驗(yàn)采用常規(guī)瓊脂擴(kuò)散(K-B)法。將分離菌培養(yǎng)液在牛肉膏蛋白胨瓊脂培養(yǎng)基表面涂布接種,室溫干燥10 min 后用無(wú)菌鑷子取出藥敏片緊貼于培養(yǎng)基表面,每個(gè)處理3 個(gè)重復(fù),然后將培養(yǎng)基倒置放在30 ℃培養(yǎng)箱中培養(yǎng)24 h,取出觀察并測(cè)定抗生素紙片周?chē)该魅χ睆剑型该魅t表示對(duì)該藥物耐藥,根據(jù)直徑大小判定菌株的耐藥強(qiáng)弱。
1.2.3 病原菌生長(zhǎng)特性研究
調(diào)節(jié)初始液體培養(yǎng)基pH 為3、4、5、6、7、8、9。取甘油保存的菌種在牛肉膏蛋白胨培養(yǎng)基上活化后,挑取單菌落于LB 液體培養(yǎng)基中培養(yǎng)12 h,取300 μL 菌液接種到15 mL 不同pH 液體培養(yǎng)基中,于30 ℃、200 r/min 培養(yǎng)8 h 后在紫外分光光度計(jì)600 nm 處測(cè)定菌液OD 值,表示生物量,每個(gè)梯度做三次重復(fù)。
用NaCl 調(diào)節(jié)培養(yǎng)基鹽度為0%、0.5%、1.0%、1.5%、2.0%、2.5%,取300 μL 菌液接種到15 mL 不同鹽度液體培養(yǎng)基中30 ℃、200 r/min 培養(yǎng)8 h。每個(gè)梯度做三次重復(fù)。
1.2.4 產(chǎn)淀粉酶、蛋白酶檢測(cè)
純化后的菌株在LB 液體培養(yǎng)基中培養(yǎng)至對(duì)數(shù)生長(zhǎng)期,使用打孔器分別在含0.2%可溶性淀粉和含0.2%脫脂奶粉的LB 固體培養(yǎng)基中等間距打四個(gè)孔,其中三孔加入100 μL 菌液,另一孔加等體積LB液體培養(yǎng)基作為對(duì)照,30 ℃培養(yǎng)24 h 后向淀粉培養(yǎng)基中加入碘液,觀察顏色變化和透明圈大小。
1.2.5 病原菌的致病性試驗(yàn)
純化后的分離菌株接種到LB 液體培養(yǎng)基中培養(yǎng)12 h,離心收集菌體,用無(wú)菌生理鹽水將菌株配置成1.0×108CFU/mL 的菌懸液,并將菌液經(jīng)0.3%福爾馬林滅活制備滅活菌液。體質(zhì)量(242±5)g 的健康牛蛙隨機(jī)分為三個(gè)組,每組60 只,每30 只牛蛙暫養(yǎng)在200 L 的養(yǎng)殖箱里,每天換一次水,正常投喂商品飼料,暫養(yǎng)一周后,一組腹腔注射0.2 mL 108 CFU/mL 細(xì)菌,另一組注射等體積經(jīng)福爾馬林溶液滅活的細(xì)菌;第三組腹腔注射0.2 mL 0.9%生理鹽水。記錄30 d 內(nèi)蛙的外觀表現(xiàn)及發(fā)病數(shù)量。
1.2.6 統(tǒng)計(jì)分析
數(shù)值以平均值±標(biāo)準(zhǔn)差表示,用SPSS 14.0 軟件進(jìn)行統(tǒng)計(jì)學(xué)分析。采用單因素方差分析,組間兩兩比較采用卡方檢驗(yàn)(P<0.05)。
2 結(jié)果與分析
2.1 病原菌的菌落形態(tài)及生理生化指標(biāo)
從牛蛙腦組織中分離到多個(gè)形態(tài)特征相似的菌株,經(jīng)純化后篩選到1 株優(yōu)勢(shì)菌株,30℃培養(yǎng)24 h 后觀察形態(tài)特征,進(jìn)行革蘭氏染色。結(jié)果顯示,菌株長(zhǎng)勢(shì)良好,培養(yǎng)24 h 后菌落直徑1~1.5 mm,形狀規(guī)則,不透明,表面光滑,呈淡黃色。顯微鏡觀察呈桿狀,革蘭氏陰性。該菌對(duì)苯丙氨酸、蛋白胨、葡磷胨、葡萄糖、山梨醇、木糖、棉子糖試驗(yàn)反應(yīng)呈陰性,對(duì)尿素、半固體瓊脂、賴(lài)氨酸、鳥(niǎo)氨酸試驗(yàn)呈陽(yáng)性;不能利用葡萄糖酸鹽、枸櫞酸鹽,不能產(chǎn)生硫化氫(表1)。

表1 生化試驗(yàn)結(jié)果Tab.1 Biochemical test results
2.2 分子生物學(xué)鑒定結(jié)果
分離菌株經(jīng)16S rRNA 和gyrB 的特異性引物擴(kuò)增產(chǎn)物送生工有限公司測(cè)序,所得序列經(jīng)拼接后在NCBI 數(shù)據(jù)庫(kù)中進(jìn)行BLAST 比對(duì)分析。結(jié)果表明,分離菌株的16S rRNA 和gyrB 基因序列與Genbank 中其他米爾伊麗莎白菌分離株的同源性高達(dá)99.33%~99.93%,對(duì)序列比對(duì)相似度較高的序列進(jìn)行篩選后,應(yīng)用MEGA7.0 軟件構(gòu)建序列遺傳進(jìn)化樹(shù),分離株與米爾伊麗莎白菌聚為一支,結(jié)果如圖1。該結(jié)果表明,分離菌株為米爾伊麗莎白菌,菌株編號(hào)為Mir-N11,GenBank 注冊(cè)號(hào)CP090369.1。
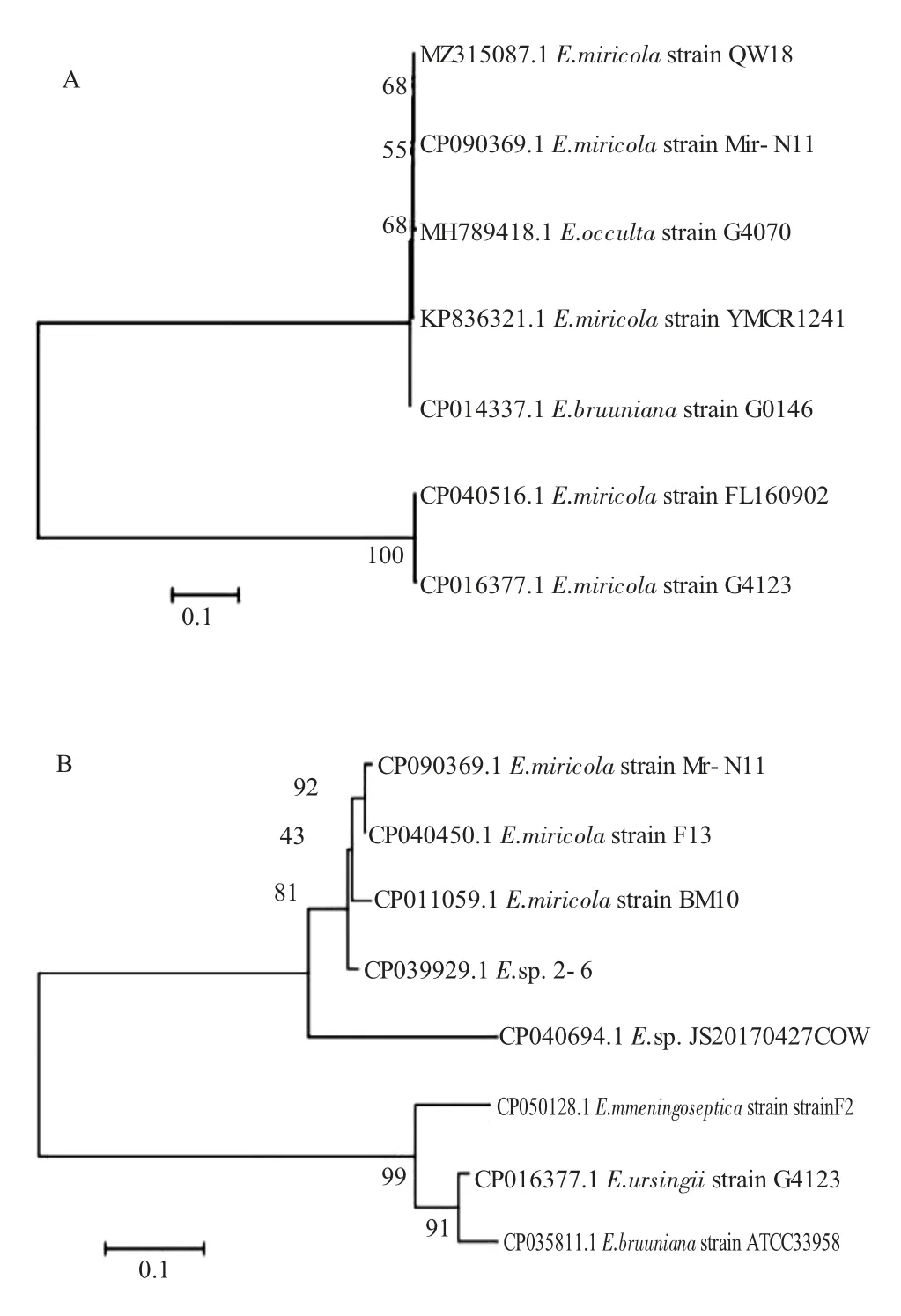
圖1 菌株Mir-N11 的16s rRNA 基因(A)和gyrB 基因(B)序列系統(tǒng)發(fā)育樹(shù)Fig.1 Phylogenetic tree of 16s rRNA gene(A)and gyrB gene(B)of strain Mir-N11
2.3 藥敏試驗(yàn)結(jié)果
菌株E.miricola Mir-N11 對(duì)米諾環(huán)素、紅霉素高度敏感,對(duì)多西環(huán)素、環(huán)丙沙星、氧氟沙星中度敏感,對(duì)哌拉西林、頭孢哌酮、麥迪霉素、羧芐西林、萬(wàn)古霉素敏感,對(duì)克林霉素、卡那霉素、四環(huán)素、氯霉素等不敏感(耐藥)(表2)。
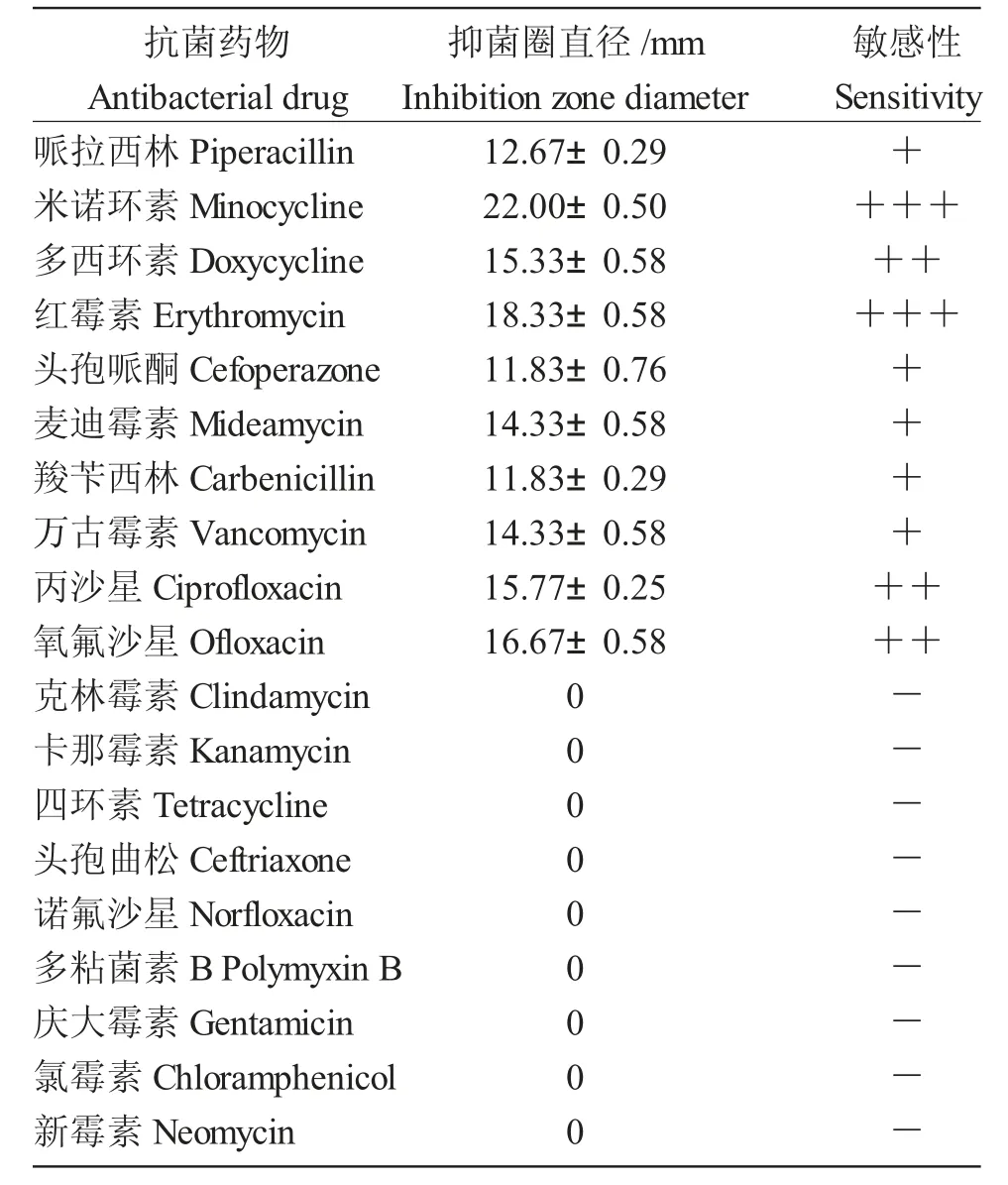
表2 藥敏試驗(yàn)結(jié)果Tab.2 Drug sensitivity test results
2.4 菌株生長(zhǎng)特性研究
菌株Mir-N11 在脫脂奶粉培養(yǎng)基中生長(zhǎng)24 h后能明顯觀察到透明圈,表明Mir-N11 具有較強(qiáng)的產(chǎn)蛋白酶能力(圖2-A);而在淀粉培養(yǎng)基中培養(yǎng)24 h 后用碘液進(jìn)行染色,在菌落周?chē)从^察到透明圈,表明菌株Mir-N11 沒(méi)有產(chǎn)淀粉酶的能力(圖2-B)。菌株Mir-N11 在pH 3.0~9.0 范圍內(nèi)均可生長(zhǎng),最適生長(zhǎng)pH 為6.0(圖3-A);在鹽度0.5%~2.5%范圍內(nèi)均可生長(zhǎng),最適生長(zhǎng)鹽度為0.5%(圖3-B)。

圖2 菌株Mir-N11 產(chǎn)蛋白酶(A)和淀粉酶(B)能力Fig.2 Protease(A)and amylase(B)production capacity of strain Mir-N11
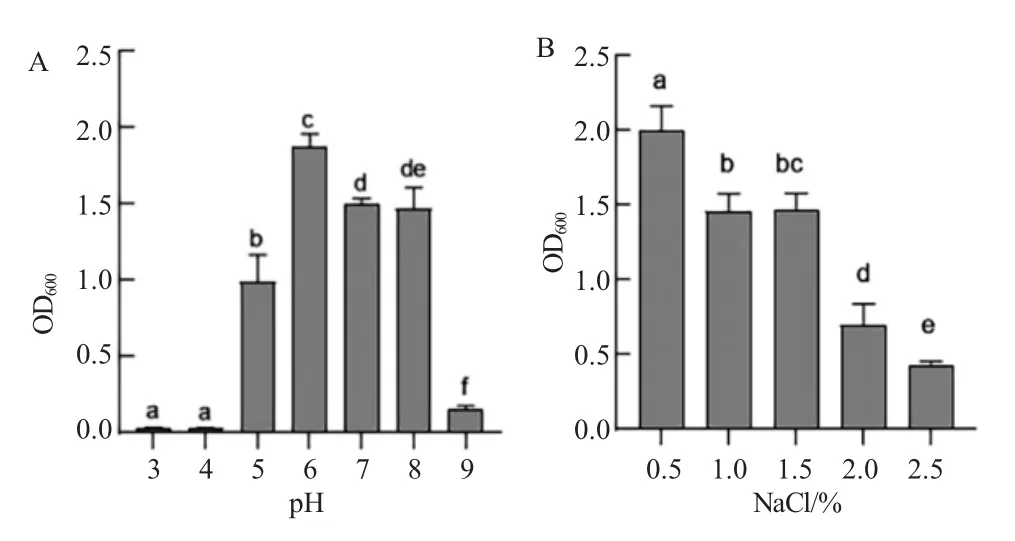
圖3 pH 和鹽度對(duì)菌株Mir-N11 生長(zhǎng)的影響Fig.3 Effects of pH and salinity on the growth of strain Mir-N11
2.5 菌株Mir-N11 致病性驗(yàn)證
活菌注射組的牛蛙在注射后第5 d 開(kāi)始出現(xiàn)不攝食、歪頭、眼球發(fā)白、水中轉(zhuǎn)圈等癥狀,30 d 內(nèi)累計(jì)發(fā)病率為43%,而滅活細(xì)菌注射組和對(duì)照組的牛蛙試驗(yàn)期間未出現(xiàn)發(fā)病(圖4)。進(jìn)一步分離表現(xiàn)明顯癥狀的病蛙腦組織細(xì)菌,將16s rRNA 基因測(cè)序,并在NCBI 上進(jìn)行序列比對(duì),鑒定為米爾伊麗莎白菌,與自然患病蛙細(xì)菌分離鑒定結(jié)果一致,這進(jìn)一步表明,菌株Mir-N11 具有較強(qiáng)的致病能力,是造成此次牛蛙歪頭病的病原菌。
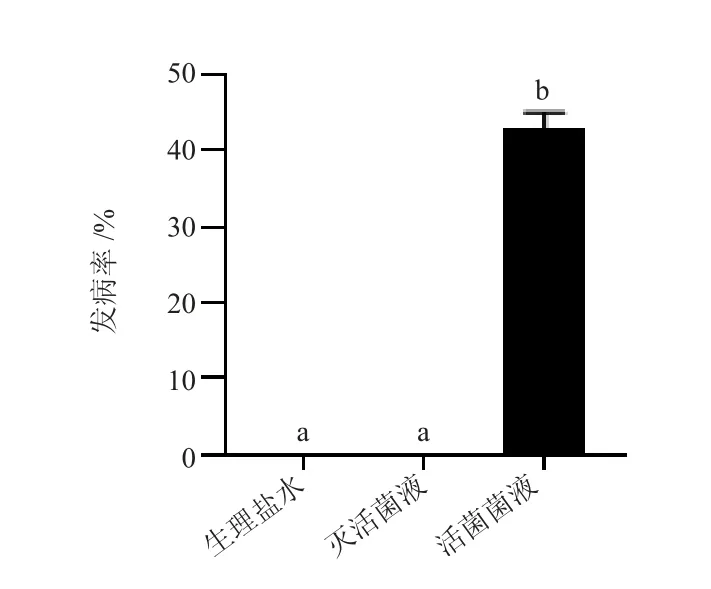
圖4 不同處理組的牛蛙30 d 內(nèi)發(fā)病情況統(tǒng)計(jì)Fig.4 Incidence statistics of bullfrog in different treatment groups within 30 days
3 討論
蛙類(lèi)歪頭、白內(nèi)障病病原的研究倍受關(guān)注。但伊麗莎白菌屬的病原菌分類(lèi)地位及分類(lèi)體系的混亂,很長(zhǎng)時(shí)間以來(lái)沒(méi)有統(tǒng)一蛙歪頭、白內(nèi)障病病原的分類(lèi)地位。陳耀明[18]報(bào)道了腦膜膿毒性黃桿菌會(huì)引起病蛙歪頭和白內(nèi)障病癥。陳會(huì)波等[19]發(fā)現(xiàn),牛蛙歪頭白內(nèi)障病原是黃桿菌。在本研究中,從患有歪頭白內(nèi)障病的牛蛙腦組織中分離得到一株米爾伊麗莎白菌,經(jīng)生理生化、16s rRNA 和gyrB 基因測(cè)序,鑒定其為E.miricola,通過(guò)回歸感染試驗(yàn)證實(shí)了該菌株能引起牛蛙歪頭白內(nèi)障病。
藥敏試驗(yàn)結(jié)果顯示,分離株對(duì)哌拉西林、紅霉素、多西環(huán)素、環(huán)丙沙星、頭孢哌酮等藥物敏感;對(duì)克林霉素、卡那霉素、慶大霉素、氯霉素等耐藥。秦振陽(yáng)等[15]的研究顯示,從患歪頭病黑斑蛙中分離的米爾伊麗莎白菌對(duì)鏈霉素、強(qiáng)力霉素、麥迪霉素等多種藥物敏感,對(duì)諾氟沙星、氧氟沙星、多粘菌素B等耐藥。雷雪平等[16]的研究顯示,從患病棘蛙中分離得到的米爾伊麗莎白菌對(duì)阿米卡星、阿芐西林、紅霉素、慶大霉素、氧氟沙星、四環(huán)素耐藥,但僅對(duì)氟苯尼考敏感。李川北等[22]從患歪頭病黑斑蛙中分離的米爾伊麗莎白菌對(duì)米諾環(huán)素、氟苯尼考、利福平和萬(wàn)古霉素敏感,對(duì)青霉素、氨芐青霉素、頭孢氨芐、頭孢他啶、頭孢克洛等耐藥。造成耐藥表型不一致的原因可能是耐藥基因隱性表達(dá)或不表達(dá)[23],或者細(xì)菌處于異質(zhì)性耐藥[24]。細(xì)菌耐藥性的獲得與生物膜的形成有關(guān)[25]。不同菌株的藥敏差異說(shuō)明不同時(shí)間、不同地域養(yǎng)殖過(guò)程中抗生素的使用差異可能是菌株耐藥性差異的主要原因[26]。
致病菌在生長(zhǎng)繁殖過(guò)程中會(huì)產(chǎn)生多種胞外活性產(chǎn)物,包括胞外蛋白酶、溶血素、明膠酶、脂肪酶、和淀粉酶等[27]。本研究通過(guò)平板打孔法初步測(cè)定了米爾伊麗莎白菌胞外產(chǎn)物的酶活性。結(jié)果表明,分離株Mir-N11 能夠產(chǎn)生胞外蛋白酶,但不產(chǎn)生淀粉酶。胞外產(chǎn)物與病原菌致病作用密切相關(guān)[28],能夠在一定程度上增加病原菌的毒性[29]。張飛等[30]在研究大黃魚(yú)(Pseudosciaena crocea)發(fā)光桿菌(Photobacterium damselae)的致病能力時(shí)發(fā)現(xiàn),胞外蛋白酶與明膠酶、淀粉酶等協(xié)同作用致大黃魚(yú)患病。金珊等[31]發(fā)現(xiàn),溶藻弧菌(Vibrio alginolyticus)的胞外產(chǎn)物中具有明膠酶、淀粉酶、酪蛋白酶、脂肪酶活性及溶血活性,對(duì)大黃魚(yú)有直接致死作用。牟海津等[32]在副溶血弧菌(Vibrio parahaemolyticus)的胞外產(chǎn)物中檢測(cè)到蛋白酶、淀粉酶、卵磷脂酶、幾丁質(zhì)酶活性,表明胞外產(chǎn)物在感染對(duì)蝦的過(guò)程中起至關(guān)重要作用。許兵等[33]研究表明,導(dǎo)致對(duì)蝦紅腿病的毒力因子是副溶血弧菌和溶藻弧菌的胞外蛋白酶。綜上所述,致病菌胞外產(chǎn)物在致病過(guò)程中起著重要作用,不同細(xì)菌的胞外產(chǎn)物也不盡相同,對(duì)于米爾伊麗莎白菌其他胞外產(chǎn)物活性以及在致病過(guò)程中的作用還有待進(jìn)一步研究,以便進(jìn)一步闡明其致病機(jī)理。
結(jié)論:分離得到一株強(qiáng)致病菌,經(jīng)鑒定為米爾伊麗莎白菌。該菌對(duì)牛蛙具有較強(qiáng)的致病性和多重抗耐藥性;可產(chǎn)生胞外蛋白酶,但不產(chǎn)淀粉酶。本研究結(jié)果可為E.miricola 病原流行分析及防治提供參考資料。
猜你喜歡
保健醫(yī)苑(2022年5期)2022-06-10 07:46:38
現(xiàn)代臨床醫(yī)學(xué)(2022年3期)2022-06-06 07:59:40
昆明醫(yī)科大學(xué)學(xué)報(bào)(2022年1期)2022-02-28 07:43:40
天津醫(yī)科大學(xué)學(xué)報(bào)(2021年3期)2021-07-21 09:04:02
科學(xué)大眾(2020年12期)2020-08-13 03:22:22
云南醫(yī)藥(2019年3期)2019-07-25 07:25:10
現(xiàn)代檢驗(yàn)醫(yī)學(xué)雜志(2016年1期)2016-11-12 13:19:40
國(guó)外醫(yī)藥(抗生素分冊(cè))(2016年6期)2016-07-10 11:34:45
中國(guó)衛(wèi)生標(biāo)準(zhǔn)管理(2015年14期)2016-01-15 02:58:37
中國(guó)當(dāng)代醫(yī)藥(2015年17期)2015-03-01 02:03:58
