小麥Tappc3A基因克隆及功能預測
2023-09-02 07:25:16向桂麗烏日娜YamamotoNaoki吳一超廖明莉魏淑紅彭正松楊在君
華北農學報 2023年4期
向桂麗,烏日娜,Yamamoto Naoki,吳一超,蔣 進,廖明莉,魏淑紅,彭正松,楊在君
(1.西華師范大學 生命科學學院,西南野生動植物資源保護教育部重點實驗室,四川 南充 637000;2.西華師范大學 環境科學與工程學院,四川 南充 637000;3.南充市農業科學研究院,四川 南充 637000;4.西昌學院 農業科學學院,四川 西昌 615013)
小麥(TriticumaestivumL.)是典型的C3植物,也是世界絕大多數地區的主要糧食作物之一[1-2]。在過去的幾十年里,以半矮稈小麥育種為代表的“綠色革命”使小麥的產量大幅度提高,在很大程度上保障了過去幾十年全世界的糧食安全[3]。但由于人口的迅速增加以及生物和非生物脅迫等因素的影響,到2050年小麥產量仍需每年增加1.7%左右方可滿足全球的需求[4]。因此,提高小麥的單產水平仍然是小麥育種的根本目標。與其他谷類作物一樣,小麥花器官的發育直接影響其產量,而花器官異常可以為改良小麥產量提供獨特的遺傳資源[5]。三雌蕊小麥(TP)具有3個正常的雄蕊和3個可育的雌蕊,能結3顆種子,具有顯著的穗粒數優勢[6]。從TP與中國春(CS)的雜交后代中篩選出的雄蕊同源轉化為雌蕊突變體(HTS-1)除具有三雌蕊性狀外,其雄蕊全部或部分同源轉化為雌蕊,自然結實率僅為12%左右,而正常雌蕊和完全轉化的雌蕊發育正常,一朵小花中可結3~4粒種子[7]。因此,TP和HTS-1在提高小麥產量和創制雜交小麥中具有較大的潛力。
磷酸烯醇式丙酮酸羧化酶(PEPC;EC4.1.1.31)催化磷酸烯醇式丙酮酸(PEP)發生不可逆的β-羧基化反應,生成草酰乙酸(OAA)和無機磷酸(Pi)。PEPC在植物、藻類和細菌中廣泛存在,但在動物和真菌中缺失[8]。在植物中,PEPC起著變構酶的作用,并被PEPC蛋白激酶磷酸化[9-10]。在C4植物和景天酸代謝途徑(CMA)的植物中PEPC得到了廣泛而深入的研究,因為在光合作用中它是催化大氣中CO2固定初始反應的關鍵酶[11-12]。在非光合細胞和C3植物中PEPC也發揮著重要作用,例如:補充由脂質合成、生物合成和氮同化等過程消耗的三羧酸循環(TCA)中間體,參與果實成熟、種子形成和發芽等生物過程[12-13]。該酶還在保衛細胞的氣孔開放和固氮豆科植物的根瘤中發揮特殊作用[14-15]。基因組分析表明,C3植物中包含一個小的PEPC家族,主要由植物型PEPC(PTPC)和細菌型PEPC(BTPC)2種類型組成[16]。與植物型PEPC不同,細菌型PEPC在其N端不存在絲氨酸磷酸化結構域。
C4植物在低CO2濃度、高溫、強光和干旱脅迫條件下具有更高的光合作用水平、水和氮的利用效率以及更高的生物產量。因此,將C4基因轉入C3作物,以提高C3植物的光合速率和耐熱性。PEPC是C4途徑的第一個關鍵酶,它一直是C3植物的轉化靶點[17-18]。研究表明[19],通過將C4植物玉米(ZeamaysL.)的PEPC基因轉入C3植物小麥中,可以增強轉基因小麥光化學和抗氧化酶活性,上調光合作用相關基因的表達,延遲葉綠素的降解,改變小麥中脯氨酸和其他代謝物的含量,并最終提高其耐熱性。到目前為止,尚無關于PEPC參與小麥器官發育的報道。Yamamoto等[20]研究表明,小麥Tappc3基因在雌蕊中表達量較高,因此,推測其與雌蕊的發育相關。近期,筆者從Yang等[21]利用HTS-1的正常雌蕊(P)、雌蕊化雄蕊(PS)和TP的正常雄蕊(S)為材料構建的RNA-Seq數據庫中篩選出一個在PS和P中表達量異常高的基因,經序列比對分析發現,該基因與節節麥(AegilopstauschiiCoss.)和大麥(HordeumvulgareL.)中編碼PEPC3蛋白的基因同源,由于該基因位于3A染色體上,因此命名為Tappc3A。
在本研究中從HTS-1和三雌蕊近等基因系CM28TP中克隆了Tappc3A基因,并對其表達模式、共表達基因及原核表達后酶的活性進行了分析。結果表明,Tappc3A可能參與小麥雌蕊發育,它在雄蕊中的過量表達可能與HTS-1的雄蕊同源轉化為雌蕊性狀形成有一定的關聯。本研究為進一步闡明Tappc3A基因的功能及其小麥雌蕊發育的調控機制奠定了基礎。
1 材料和方法
1.1 試驗材料
本研究選用三雌蕊小麥CM28TP和小麥雄蕊同源轉化雌蕊突變體HTS-1為試驗材料。CM28TP是小麥三雌蕊突變體(TP)與川麥28(Chuanmai 28,CM28)雜交后與CM28回交7代后獲得的三雌蕊小麥株系[22]。HTS-1是從TP與中國春(Chinese spring,CS)的雜交后代中篩選出的一個雄蕊同源轉化為雌蕊的突變體,其雄蕊部分或完全轉變為雌蕊,因此具有4~6個雌蕊。因此,HTS-1與CM28TP中均含有控制小麥三雌蕊性狀的關鍵基因Pis1。小麥進入孕穗期后,分別取采集長度為0.2~0.5 cm(二棱期至小花分化期),0.5~0.7 cm(雌雄蕊原基分化期)和0.7~1.0 cm(藥隔時期)的3個階段幼穗[23]。同時采集HTS-1的雌蕊(P)、雌蕊化雄蕊(PS)及CM28TP雄蕊(S)。將上述材料浸入RNA保存液(TaKaRa,中國大連)中,置于-80 ℃超低溫冰箱中保存備用。
1.2 總RNA提取和cDNA合成
利用LABGENETMplant RNA isolation Kit(蘭博基因,中國江蘇)分別提取HTS-1和CM28TP 3個階段的幼穗及P、PS、S的總RNA,具體提取方法參照生產商提供的說明書執行。利用1%瓊脂糖凝膠電泳和NaroDrop 2000c微量分光光度計檢測RNA的純度和濃度。以上述提取的RNA為模板,利用PrimeScriptTMRT Reagent Kit(TaKaRa,中國大連)參照說明書提供的方法進行反轉錄,并將得到的cDNA溶液置于-20 ℃的冰箱中保存備用。
1.3 Tappc3A的克隆測序
基于小麥TraesCS3A02G306700.1序列,利用Primer Premier 6.0 設計Tappc3A擴增引物(表1)。以HTS-1和CM28TP藥隔時期幼穗cDNA作為PCR擴增的模板,退火溫度56 ℃。將擴增純化后的PCR產物連接到pMD19-T載體上,導入DH5α感受態細胞中,最后涂布于含100 μg/mLAmp+的LB固體培養基上,37 ℃靜置培養過夜。陽性克隆菌液送至生工生物工程(上海)股份有限公司測序,利用DNAMAN 9.0和Chromas對測序結果進行分析。
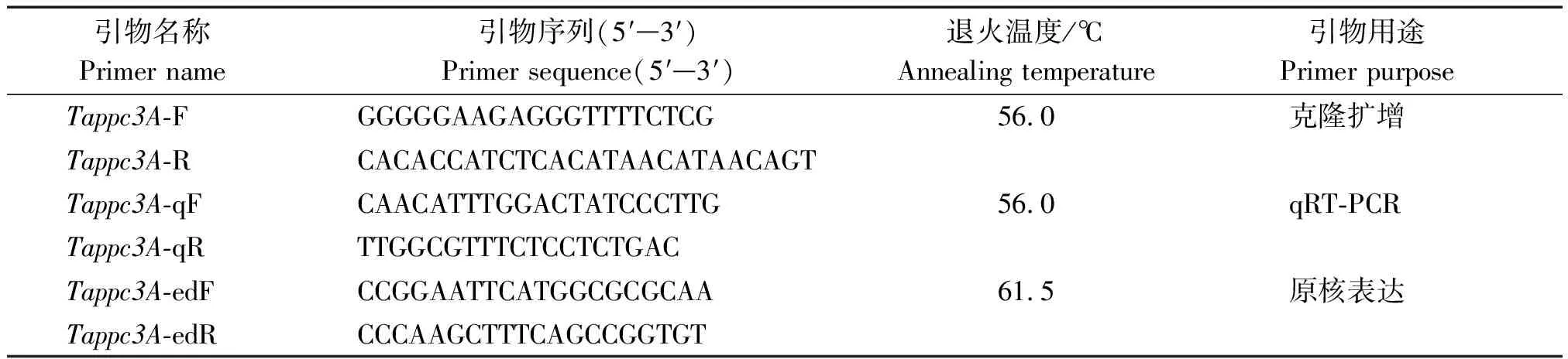
表1 Tappc3A基因克隆、qRT-PCR及原核表達引物序列
1.4 Tappc3A氨基酸序列分析及系統進化樹的構建
獲得的cDNA序列利用NCBI在線軟件ORF Finder(https://www.ncbi.nlm.nih.gov/orffinder/)進行開放閱讀框的分析,DNAMAN 9.0軟件進行氨基酸序列比對;利用NCBI的蛋白結構域數據庫(Conserved domain database,CDD,https://www.ncbi.nlm.nih.gov/cdd)和InterPro在線分析工具(http://www.ebi.ac.uk/interpro/)分析蛋白保守結構域及功能域;使用ExPasy(https://web.expasy.org/protparam/)分析蛋白的分子量、理論等電點和蛋白質的親疏水性;采用NovoPro網站(https://www.novopro.cn/tools/)預測蛋白的二級結構;運用NCBI的Blast(https://blast.ncbi.nlm.nih.gov/Blast.cgi)搜索同源基因,使用ClustalX 1.83軟件進行多重序列比對,再使用MEGA 7.0軟件中的鄰近相連法構建系統發育樹,并進行Bootstrap檢測。
1.5 熒光定量PCR(qRT-PCR)分析
利用Primer Premier 6.0 設計Tappc3A的qRT-PCR引物,引物序列如表1所示。以1.2中的cDNA作為模板,利用CFX 96 PCR儀(Bio-Rad,美國加州)進行qRT-PCR,具體操作參照SsoFastTMEva Green Supermix(Bio-Rad,美國加州)的說明書進行。以小麥Actin基因作為內參(GenBank ID:AB1811991)[20],利用2-ΔΔCt的方法計算各樣品中Tappc3A基因的相對表達量[24]。利用Graphpad軟件作圖。
1.6 重組表達載體pET-28a-Tappc3A的構建及誘導表達
根據Tappc3A基因的ORF序列分別設計帶有EcoR Ⅰ(5′-G↑AATTC-3′)和HindⅢ(5′-A↑AGCTT-3′)限制性內切酶位點的引物(表1)。以1.3中克隆且測序正確的菌液為模板進行菌落PCR,退火溫度61.5 ℃。利用EcoRⅠ和HindⅠ對擴增回收產物和pET-28a質粒進行雙酶切,再利用T4DNA連接酶連接,獲得pET-28a-Tappc3A重組載體。利用熱激法將重組載體轉入大腸桿菌DH5α中,利用PCR和雙酶切進行驗證。最后將驗證正確的菌液送生工生物工程(上海)股份有限公司測序。將測序正確的重組質粒pET-28a-Tappc3A轉化到大腸桿菌BL21(DE3)中,獲得的pET-28a-Tappc3A-BL21重組菌株在LB(含100 μg/mL Kan)液體培養基中,180 r/min,30 ℃培養。培養至OD值0.6左右加入終濃度為0.2 mmol/L的IPTG誘導劑,30 ℃誘導5 h。12 000 r/min,4 ℃離心10 min收集菌液細胞。按照鈺博生物的大腸桿菌總蛋白提取試劑盒提取融合蛋白,通過冰浴超聲得到大腸桿菌總蛋白樣品。按照磷酸烯醇式丙酮酸羧化酶(PEPC)試劑盒(蘇州科銘生物技術有限公司)說明書操作,在340 nm測定NADH減少速率,從而計算PEPC酶活性。
1.7 挖掘與Tappc3A共表達的基因
利用中國春小麥(CS)及其近等基因系17種RNA-Seq數據庫中獲得TPM值,挖掘與Tappc3A共表達的基因[20]。這些共表達基因包括抽穗期和開花期的旗葉,開花早期和晚期的莖、根、雌蕊、雌蕊化雄蕊、雄蕊、子房,孕穗期的莖和穗,種子萌發期的胚芽鞘、胚、根尖以及幼苗期葉片。計算Tappc3A與其他基因之間表達量的Pearson相關系數(PCCs)。將正相關的Pearson相關系數閾值設定為0.7[25]。利用agriGO(http://bioinfo.cau.edu.cn/agriGO/index.php)對與Tappc3A共表達的基因進行GO富集分析。利用bioinformatics在線工具(http://www.bioinformatics.com.cn/)繪制GO富集的氣泡圖。
2 結果與分析
2.1 Tappc3A基因克隆及序列分析
以HTS-1和CM28TP藥隔時期幼穗cDNA作為PCR擴增的模板,克隆獲得一條約3 000 bp的單一條帶,與預期結果相符(圖1-A)。經測序和比對分析后發現從HTS-1和CM28TP中克隆得到的目的基因序列完全一致。克隆得到的Tappc3A長度為2 970 bp,開放性閱讀框(ORF)長度為2 901 bp,編碼966個氨基酸殘基(圖1-B)。通過NCBI上的BlastN比對后發現,該基因序列與其他植物的PEPC基因序列具有77.14%~99.93%的相似性,其中與小麥中國春品種3A染色體上的PEPC基因(XM_044485447.1)具有99.93%的一致性,與二穗短柄草(Brachypodiumdistachyon,XM_014898420.2)和酸棗(Ziziphusjujubavar.spinosa,XM_048471538.1)的PEPC基因分別具有89.77%和78.54%的一致性。因此,將該基因命名為Tappc3A,并將該序列上傳NCBI數據庫(登錄號:OP615936)。
運用DNAMAN 9.0軟件進行氨基酸序列多重比對分析發現,目的基因與中國春(XP_044341382.1)、二穗短柄草(XP_014753906.1)、水稻(Oryzasativa,XP_015630901.1)、高粱(Sorghumbicolor,P15804.2)、黃花菊(Flaveriatrinervia,P30694.2)的氨基酸序列相似度分別為99.69%,91.54%,90.82%,73.17%,81.11%。比對結果顯示(圖2),Tappc3A基因編碼蛋白序列在N端具有保守的絲氨酸(Ser,S)可逆磷酸化位點(SIDAQLR),C端具有植物型PEPC蛋白特征序列(QNTG),氨基酸序列第774位是C3型PEPC的丙氨酸(Ala,A)而非高粱和黃花菊C4型PEPC的絲氨酸(Ser,S),表明小麥Tappc3A蛋白是植物型C3型PEPC。
2.2 Tappc3A蛋白的特征性分析與系統進化樹構建
在CDD數據庫中對Tappc3A結構域進行預測,發現該蛋白在氨基酸序列163—966位具有一個磷酸烯醇式丙酮酸羧化酶(PEPase)保守結構域,屬于磷酸烯醇式丙酮酸羧化酶超級家族成員(圖3-A)。進一步分析顯示,在該結構域中具有2個PEPase活性位點,分別在氨基酸序列第168—179位(PS00781,VLTAHPTQSVRR)和第592—604位(PS00393,VMIGYSDSGKDAG),在這2個活性位點中分別含有與酶活性有關的氨基酸殘基賴氨酸(Lys,K)和組氨酸(His,H),同時具有7個保守的PEPcase亞催化結構域(圖2)。該蛋白質分子式C4917H7778N1356O1442S35,相對分子質量為110.08 ku,理論等電點為6.02,不穩定系數45.18,總平均親水性為-0.358,因此,該蛋白屬于酸性親水性蛋白。二級結構分析預測,該蛋白含有62.84%的α-螺旋、4.35%的β-轉角、5.69%的延伸鏈和27.12%的無規則卷曲(圖3-B、C)。為了進一步了解Tappc3A基因的生物學功能,采用MEGA 7.0軟件N-J構建植物Tappc3A蛋白氨基酸序列的系統進化樹(圖4)。結果表明,相同光合作用途徑且親緣關系近的物種首先聚為一類,Tappc3A基因氨基酸序列與烏拉爾圖小麥(Triticumurartu)、野生二粒小麥(Triticumdicoccoides)、大麥(Hordeumvulgare)等的親緣關系最近,同屬于C3型分支。

圖4 Tappc3A與其他植物PEPC的系統進化樹
2.3 Tappc3A基因qRT-PCR分析
進一步探討克隆得到的Tappc3A基因在CM28TP和HTS-1中的表達是否有差異,通過qRT-PCR對CM28TP和HTS-1 3個階段的幼穗及P、PS、S的表達量進行分析(圖5)。結果顯示,Tappc3A基因在2種材料及P、PS、S中均有表達,但各個時期及P、PS、S表達量有顯著差異。在CM28TP小穗中,Tappc3A的表達量在雌雄蕊原基分化期(0.5~0.7 cm)最高,其次是二棱期至小花分化期(0.2~0.5 cm),表達量最低的是藥隔時期(0.7~1.0 cm)。在HTS-1小穗中,Tappc3A的表達量在二棱期至小花分化期最高,雌雄蕊原基分化期和藥隔時期表達量相近。在二棱期至小花分化期和藥隔時期,HTS-1小穗中Tappc3A的表達量都要顯著高于CM28TP,而在雌雄蕊原基分化期無顯著差異。Tappc3A在P和PS中的表達量顯著高于S的表達量。
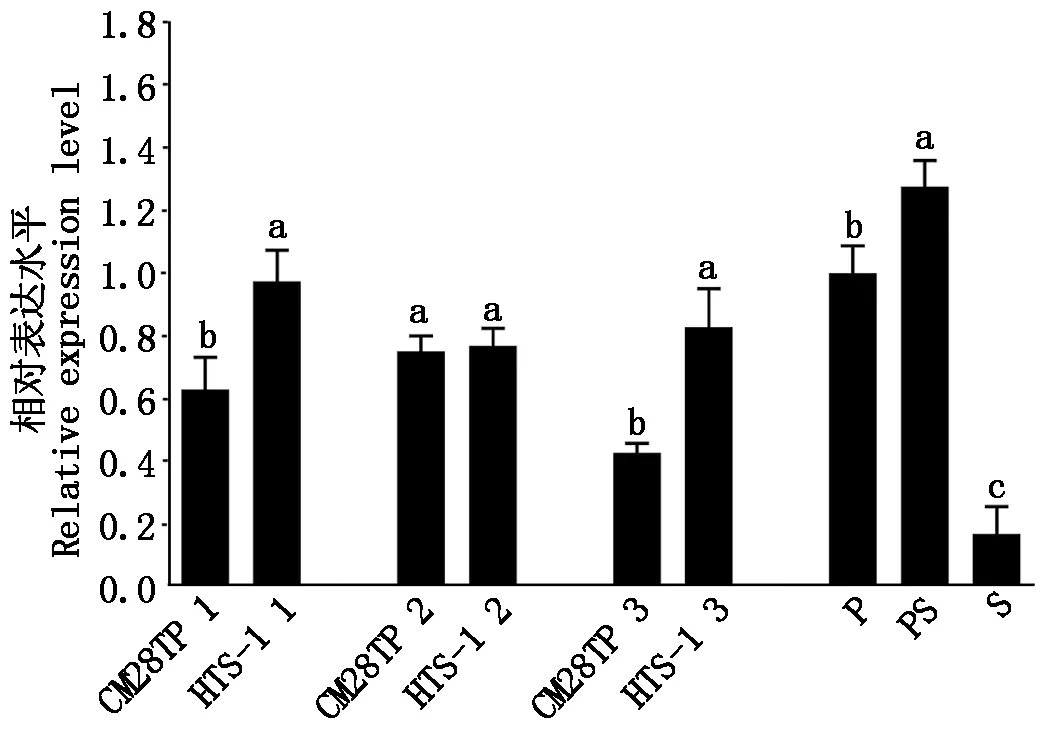
1.二棱期至小花分化期0.2~0.5 cm;2.雌雄蕊原基形成期0.5~0.7 cm;3.藥隔期0.7~1.0 cm;P.雌蕊;PS.雌蕊化雄蕊;S.雄蕊。不同字母的均值表示存在差異顯著(P<0.05)。圖6同。
2.4 重組蛋白誘導表達及PEPC活性分析
構建的pET-28a-Tappc3A重組質粒,經雙酶切和電泳檢測后獲得2條帶,一條大小為5 300 bp左右(pET-28a載體),一條大小為2 900 bp左右(目的條帶),而未經雙酶切的pET-28a-Tappc3A線性質粒只有一條大小8 000 bp左右的條帶,條帶大小與預測的一致(圖6-A)。重組質粒測序結果與目的基因序列一致,證明重組質粒pET-28a-Tappc3A構建成功。將重組質粒轉化到大腸桿菌BL21(DE3)中,活化獲得pET-28a-Tappc3A-BL21重組菌株。對IPTG誘導濃度、誘導時間及誘導溫度進行優化,pET-28a-Tappc3A-BL21在30 ℃、IPTG終濃度為0.2 mmol/L時,誘導5 h后,Tappc3A蛋白表達量最大。表達菌液經超聲破碎后取上清部分,測定融合蛋白中PEPC的活性。結果表明(圖6-B),pET-28a空載體組中是否添加IPTG對PEPC活性影響顯著,且酶活性均較低;與pET-28a空載組相比,pET-28a-Tappc3A組的PEPC活性增強,加入IPTG誘導后,其酶活性進一步得到顯著提升。原核表達分析證明,Tappc3A基因編碼的蛋白能催化磷酸烯醇式丙酮酸(PEP)生成草酰乙酸,并且在IPTG誘導后活性顯著增強。
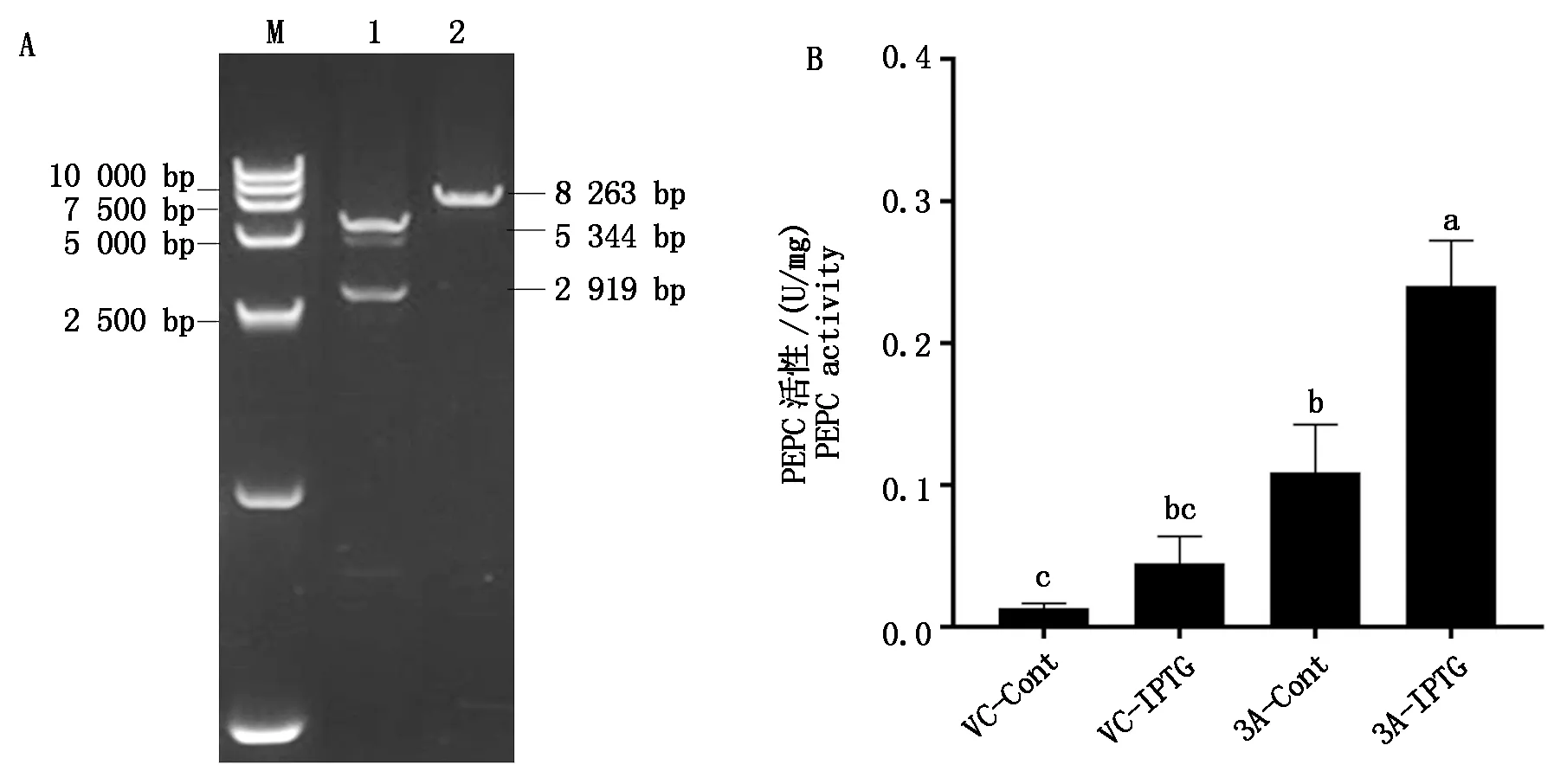
A.重組質粒 pET-28a-Tappc3A的雙酶切鑒定:M.Marker 15000;1.pET-28a-Tappc3A質粒經 EcoRⅠ、Hind Ⅲ雙酶切;2.pET-28a-Tappc3A線性質粒。B.PEPC活性測定:VC.對照組;3A.試驗組。
2.5 與Tappc3A共表達的基因分析
為探究Tappc3A與其他調控花器官發育基因之間是否存在相互關系,從小麥RNA-Seq數據庫中挖掘出與Tappc3A共表達的基因。當Pearson相關系數的閾值設定為0.7時,從小麥17個RNA-Seq數據庫中共篩選到了11 862個基因,其中3 492個基因得到GO注釋信息,當P<0.05時獲得674個富集顯著的GO term。在這些GO term 中有18個GO term 與花器官的發育與分化相關(圖7)。這些GO term主要包括:花器官形成、生殖芽系統發育、胚胎后器官形態發生、花器官形態發生、生殖過程的調節、單體生殖過程、生殖過程、花器官的發育、花器官特性的維持、器官特性的維持、生殖結構發育、生殖系統發育、多細胞生物繁殖過程、分生組織營養期到生殖期的轉變、器官形態發生、花輪發育、胚胎后器官發育和花器官特性規范等。這表明Tappc3A可能與這些基因一起調控了小麥花器官的發育。
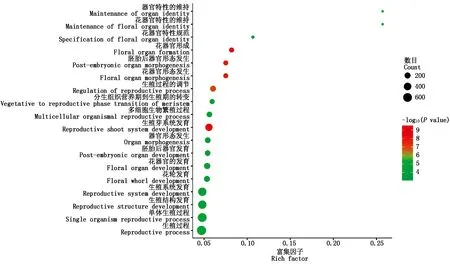
顏色表示富集程度;大小表示基因數量。
3 結論與討論
在C3植物中磷酸烯醇式丙酮酸羧化酶(Phosphoenolpyruvate carboxylase,PEPC)的主要作用是催化磷酸烯醇式丙酮酸(PEP)生成草酰乙酸(OAA)進入三羧酸循環,從而彌補三羧酸循環中消耗的中間產物[26-27]。此外,PEPC在植物的生長發育和逆境適應過程中也發揮了重要的作用,如果實的成熟、種子的萌發和發育、抗旱、抗鹽等[28-31]。到目前為止,除Igawa等[32]報道了細菌型PEPC基因可以加速花粉成熟過程中貯藏物質的積累外,尚無關于PEPC參與植物器官發育的報道。Yamamoto等[20]研究表明,小麥Tappc3基因在雌蕊中表達量較高,因此,推測其與雌蕊的發育相關。本研究以小麥2個特有的雌蕊突變體CM28TP和HTS-1為試驗材料克隆了一個PEPC家族基因Tappc3A,它是Tappc3位于3A染色體上的同源基因,該基因具有典型的磷酸烯醇式丙酮酸羧化酶的活性位點和功能位點,第774位氨基酸為C3植物PEPC典型的丙氨酸。聚類分析結果也表明,Tappc3A屬于C3型PEPC家族。原核表達分析結果表明,構建pET-28a-Tappc3A重組質粒在大腸桿菌中能表現出磷酸烯醇式丙酮酸羧化酶活性,且IPTG誘導后酶活性明顯增強。這表明本研究獲得了具有正常功能的Tappc3A基因。通過對P、PS和S的RNA-Seq數據分析表明Tappc3A在P和PS中的表達量顯著高于S,因此,推測小麥雄蕊同源轉化為雌蕊可能與Tappc3的過量表達相關[21]。序列比對結果分析表明,在CM28TP和HTS-1中克隆得到Tappc3A基因ORF序列及推導氨基酸序列完全相同,這表明,雄蕊同源轉化為雌蕊性狀不是Tappc3A基因序列的差異造成的,可能與其表達量有關。
前期研究表明,小麥PEPC基因在不同組織中的表達具有特異性,如Tappc1a基因在根和生殖器官中表達量較高。Tappc1b基因在幼苗期的葉片中高水平表達。Tappc2的表達具有普遍性,因此,可能是一個看家基因。Tappc3雌蕊等生殖器官中高水平表達。Tappc4在成熟葉片中表達量較高[20]。在蓖麻(Ricinuscommunis)中O′Leary等[29]也觀察到Rcppc3基因在雌蕊的珠被中的表達量高于雄蕊,因此,推測Rcppc3與雌蕊發育相關。本研究重點探討了Tappc3A在小麥幼穗、雌蕊(P)、雄蕊(S)以及雌蕊化的雄蕊(PS)中的表達情況。在幼穗發育的3個階段,除階段二(雌雄蕊原基分化期)外,HTS-1中的Tappc3A基因的表達量均高于CM28TP。尤其是在第三階段(藥隔形成期)HTS-1中的Tappc3A基因表達量約為CM28TP的2倍。在這個階段,雌雄蕊原基已完成分化,雄蕊突起由球狀變為柱狀,且沿中部自頂向下出現微凹縱溝,同時雌蕊原基頂端凹陷分化出2枚柱頭原基。因此,在這個階段,雄蕊和雌蕊發育所需要的物質具有較大的差異。而Tappc3A在雄蕊中的過量表達可能造成蛋白質和脂質代謝混亂,從而導致雄蕊發育異常。已有研究表明,花藥和花粉發育過程中脂質代謝的紊亂會導致花藥角質層、花粉外壁和花藥亞細胞器膜的發育異常,從而引起細胞核雄性不育(GMS)[33]。Tappc3A基因在HTS-1的P、PS及CM28TP的S中的表達模式進一步表明,該基因可能與雌蕊發育相關,因為Tappc3A在P和PS中的表達量顯著高于S,該結果也與之前的RNA-Seq結果一致[21]。共表達分析顯示,Tappc3A基因與花器官形成、花器官形態發生、花器官的發育、花器官特性的維持、器官形態發生、花輪發育和花器官特性規范等花器官的發育與分化相關基因共表達,進一步表明Tappc3A可能參與了小麥花器官的形態發生。此外,研究表明[34],基因表達模式的改變會導致小麥雄蕊心皮化。綜上所述,Tappc3A可能與小麥雌蕊發育相關,其在雌蕊中的過量表達可能與HTS-1的雄蕊同源轉化為雌蕊性狀形成有一定的關聯。本研究為進一步闡明Tappc3A基因的功能及小麥雌蕊發育的調控機制奠定了基礎。
從小麥中克隆得到Tappc3A基因,該基因編碼966個氨基酸,具有典型的磷酸烯醇式丙酮酸羧化酶的活性位點和功能位點,是植物C3型PEPC。構建的pET-28a-Tappc3A重組質粒在大腸桿菌中能表現出磷酸烯醇式丙酮酸羧化酶活性,且IPTG誘導后酶活性顯著增強,這表明Tappc3A基因具有正常的功能。在HTS-1幼穗發育的二棱期至小花分化期和藥隔時期,Tappc3A基因的表達水平都顯著高于CM28TP,且HTS-1的P和PS中的表達量顯著高于CM28TP中的S。共表達分析結果進一步表明,Tappc3A可能參與了小麥花器官的形態發生。因此,推測Tappc3A可能參與小麥雌蕊發育,其在雄蕊中的過量表達可能與小麥雄蕊同源轉化為雌蕊性狀形成有一定的關聯。
