基于Sirt1/p53信號通路探究NK4基因對非霍奇金淋巴瘤細胞增殖和凋亡的機制研究
2023-08-31 02:49:42賁海祥
河北醫學 2023年8期
賁海祥, 徐 娟
(江蘇省如皋市人民醫院血液科, 江蘇 如皋 226500)
惡性淋巴瘤(Malignant lymphoma,ML)是一種血液惡性腫瘤,發生在淋巴結或淋巴結外的淋巴組織[1]。非霍奇金淋巴瘤(Non-Hodgkin's lymphoma,NHL)是一類起源于淋巴系統的癌癥,由惡性B細胞的過度增殖引起,是美國第七大常見惡性腫瘤,通常被認為是預后良好的惡性腫瘤,5年生存率約為70%[2]。但淋巴瘤的發病率在全球范圍內持續增加,對人類健康造成嚴重危害[3]。NHL最常見的類型為彌漫大B細胞淋巴瘤(DLBCL)、濾泡性淋巴瘤(FL)和套細胞淋巴瘤(MCL)[4]。NK4是一種特異性肝細胞生長因子(HGF)拮抗劑,由HGF的N端發夾結構和α鏈的四個Kringle結構域組成。NK4蛋白給藥或NK4基因治療抑制了各種腫瘤類型中的腫瘤生長、侵襲、轉移和血管生成,所述腫瘤類型包括乳腺癌、結腸癌、肺癌、胰腺癌、間皮瘤、前列腺癌、胃癌和腦癌[5~7]。另外,去乙酰化酶sirtuin-1(Sirt1)是一種NAD+依賴性Ⅲ類組蛋白去乙酰化酶,其功能上與細胞代謝相關,并被視為代謝傳感器[8],可調節干細胞、細胞增殖、凋亡、DNA修復、自噬和腫瘤發生,Sirt1在許多癌癥中上調,包括前列腺癌、白血病和皮膚T細胞淋巴瘤腫瘤細胞[9]。沉默或抑制Sirt1的表達水平會激活下游p53蛋白,并隨后加速或推動細胞衰老[10]。目前,NK4對NHL疾病的調控機制尚鮮有研究,尤其是NK4是否通過介導Sirt1/p53信號通路調控腫瘤細胞的生物學行為仍未可知。因此,在這項研究中,我們旨在探究NK4基因對NHL細胞增殖、凋亡的調節機制。
1 材料與方法
1.1細胞培養:人類非霍奇金淋巴瘤細胞株(Jurkat和Raji)及正常B細胞系均購自美國典型培養物保藏中心(ATCC,Rockville,MD,USA)。Jurkat和Raji細胞在補充有10% FBS的RPMI 1640培養基中生長,B細胞維持在添加10%人血清、100U/mL青霉素、100μg/mL鏈霉素、2mM L-谷氨酰胺(Invitrogen,Carlsbad,CA,USA)的Iscove s改良培養基中(NABI Biopharmaceuticals,Boca Raton,FL,USA)。所有細胞系都在37℃,5% CO2條件下培養。
1.2細胞轉染:pcDNA3.1-NK4、pcDNA3.1-NC均購自Invitrogen(Carlsbad,CA,USA),Sirt1激動劑SRT1720購自碧云天生物科技有限公司(上海,中國)。oe-NK4/NC組細胞處理步驟如下:將細胞以1×106細胞/mL的密度置于6孔板上,按照操作說明書,使用LiPofectamine 2000 (11668-019,Invitrogen,Carlsbad,CA,USA)將50 nM pcDNA3.1-NK4/NC轉染到Raji細胞(培養至融合度80%以上)中,轉染48h。oe-Sirt1+SA/DMSO組處理步驟如下:在培養基中加入給定濃度的SIRT1激動劑SRT1720 (1moL/L)或者等量的DMSO,再轉染pcDNA3.1-NK4 48h后進行后續實驗。
1.3qRT-PCR(實時熒光定量聚合酶鏈反應):從人淋巴瘤細胞系或正常外周淋巴細胞中提取總RNA TRIzol試劑(Invitrogen),并使用反轉錄第一鏈cDNA合成試劑盒(K1622; Thermo Fisher Scientific)進行反轉錄。根據制造商的說明,使用Maxima SYBR Green/ROX qPCR master mix(K0223;Thermo Fisher Scientific)進行qRT-PCR檢測NK4 mRNA水平,β-actin用于歸一化。用于qRT-PCR的引物如下,NK4 F:5′-GTGAATACTGCAGACCAATGTGCTA-3′和NK4 R:5′-GGTCAAATTCATGGCCAAATTC-3′;β-actin F:5′-TGGCACCCAGCACAATGAA-3′,R:5′-CTAAGTCATAGTCCGCCTAGAAGCA-3′。使用2-△△Ct比較Ct方法計算。
1.4CCK-8檢測細胞增殖:細胞增殖通過細胞計數試劑盒-8(CCK8)分析(C0039,碧云天生物科技有限公司,北京,中國)。將不同組的細胞以2×105細胞/孔的密度接種到96孔板中,并在37℃含5% CO2的濕潤培養箱中孵育隔夜。在第0小時、第12小時、第24小時、第48小時和第72小時收集細胞。去除培養基并用無血清基本培養基替換后,向每個孔中加入10μL CCK 8溶液并孵育1h,之后將100 μL樣品上清液轉移到96孔微孔板中,并根據光密度波長為450nm。
1.5流式細胞術檢測細胞凋亡:細胞被胰蛋白酶消化并在冷PBS中洗滌兩次。在1000g離心5min后,收集細胞并濃縮至每孔1×106個細胞。將0.1mL細胞懸浮液與5μL FITC綴合的膜聯蛋白V混合,在室溫下避光孵育15min,然后與5μL PI 混合并孵育5min。然后使用流式細胞儀(FACSCanto Ⅱ,BD Biosciences,US)分析樣品。
1.6Transwell法檢測細胞遷移、侵襲:根據生產商說明,使用24孔Transwell小室(BD,Biosciences)檢測細胞遷移和侵襲能力。侵襲實驗時,先按1∶8的比例將50mg/L Matrigel膠稀釋后鋪于小室底部;遷移實驗時不鋪膠。24孔板下室加入完全培養基600μL,取各組細胞懸液200μL加入Transwell小室的上室,在37℃下孵育24h后,使用棉簽去除上室中的細胞。遷移或侵襲細胞用甲醇在4℃固定30min,用0.1%結晶紫溶液在37℃染色20min,用倒置顯微鏡(Nikon Corporation,Japan)進行觀察。細胞實驗獨立重復3次。
1.7Western blot:用含有磷酸酶和蛋白酶抑制劑的RIPA裂解緩沖液(基爾頓生物科技上海有限公司,上海,中國)獲取細胞裂解液,使用BCA蛋白質測定試劑盒(PICPI23223; Thermo Fisher Scientific)對樣品蛋白濃度進行檢測。進行加熱和變性后,使用10%(對于高分子量蛋白質)或15%(對于低分子量蛋白質)十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳分離蛋白質樣品(25μg/泳道),并轉移到硝酸纖維膜(Millipore,Billerica,MA,USA)上。在室溫下用5%脫脂牛奶封閉1h后,將膜與一抗在4℃下單獨孵育過夜。一抗信息如下:Sirt1(ab189494,1∶1000,80kDa)、p53(ab32389,1∶10000,44kDa),GAPDH(ab9485,1∶2500,37kDa)所有抗體均購自Abcam(Cambridge,MA,USA)。用含Tween-20的Tris緩沖鹽水洗滌后,將膜與辣根過氧化物酶(HRP)偶聯的山羊抗兔二抗(1∶1000; ab150077)一起在室溫下孵育1h。在Tanon 5200化學發光成像系統(上海天能科技有限公司,上海,中國)上觀察到蛋白質條帶。使用ImageJ軟件(National Institutes of Health,Bethesda,MD,USA)分析條帶密度,GAPDH條帶用作內參。
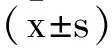
2 結 果
2.1NK4基因在NHL細胞低表達:為評估NK4基因在NHL細胞中的表達水平,我們使用qRT-PCR檢測NHL細胞系(Jurkat和Raji)及正常B細胞(normal)中NK4 mRNA的表達。結果顯示,與正常B細胞相比,NK4 mRNA水平在NHL細胞系中顯著下降(圖1,均P<0.01),其中,NK4表達水平在Raji細胞中最低。因此在后續的研究中,我們采取Raji細胞作為研究對象。

圖1 NK4基因在NHL細胞低表達
2.2上調NK4基因表達水平抑制NHL細胞生長進展:為探索NK4細胞在NHL細胞中的作用,我們使用pcDNA3.1-NK4及其陰性對照pcDNA3.1-NC轉染至Raji細胞中。qRT-PCR結果顯示,轉染pcDNA3.1-NK4組(oe-NK4組)NK4 mRNA水平顯著上升,轉染成功(圖2A,P<0.001)。隨后,我們分別采取CCK-8和流式細胞術探究細胞增殖和凋亡,結果顯示,與轉染pcDNA3.1-NC組(oe-NC組)相比,上調NK4表達水平顯著抑制NHL細胞增殖、促進細胞凋亡(圖2B-C,均P<0.001)。此外,在NHL細胞中上調NK4表達水平后,NHL細胞遷移和侵襲能力顯著下降(圖2D,均P<0.01)。以上結果表明,上調NK4基因表達水平抑制NHL細胞生長進展。

圖2 上調NK4基因表達水平抑制NHL細胞生長進展
2.3NK4基因抑制Sirt1/p53信號通路:Western blot檢測結果發現,與正常B細胞相比,Sirt1蛋白表達水平在Raji細胞中顯著上調(圖3,P<0.001),而p53蛋白表達水平顯著下降(圖3,P<0.001)。而在Raji細胞中過表達NK4基因后,Sirt1蛋白表達水平顯著下降,p53水平顯著上升(圖3,均P<0.001)。以上結果表明,NK4基因抑制Sirt1/p53信號通路。

圖3 NK4基因抑制Sirt1/p53信號通路
2.4激活Sirt1/p53信號通路部分逆轉過表達NK4對NHL細胞生長進展的抑制:為進一步確定NK4/Sirt1/p53在NHL細胞生長進展中發揮的作用,在上調NK4表達的Raji細胞中,同時添加Sirt1激活劑SRT1720上調Sirt1的表達。結果顯示,SRT1720顯著上調Raji細胞中Sirt1蛋白表達水平,顯著降低p53蛋白表達水平(圖4A,均P<0.01)。本研究還觀察到Sirt激活劑顯著促進NHL細胞增殖、抑制細胞凋亡(圖4B-C,均P<0.01)。此外,在NHL細胞中添加Sirt激活劑后,NHL細胞遷移和侵襲能力顯著下降(圖4D,均P<0.01)。以上結果表明,激活Sirt1/p53信號通路部分逆轉過表達NK4對NHL細胞生長進展的抑制。

圖4 激活Sirt1/p53信號通路部分逆轉過表達NK4對NHL細胞生長進展的抑制
3 討 論
NHL覆蓋了約90%的影響淋巴結、脾、骨髓和免疫系統及其他器官的淋巴瘤。NK4基因療法可抑制多種腫瘤的生長、侵襲、轉移和血管生成[5]。在不同癌癥的實驗模型中,NK4基因治療抑制了Met受體的激活,這與抑制腫瘤的侵襲和轉移有關。類似地,NK4基因治療抑制腫瘤血管生成,從而抑制血管生成依賴性腫瘤生長[11]。因此,在本研究中筆者大膽假設NK4基因或許對NHL的發生發展發揮著關鍵作用。結果顯示,與正常B細胞相比,NK4基因在NHL細胞系(Jurkat和Raji)中低表達。這提示,差異表達的NK4或許對NHL細胞的生物學功能扮演了重要角色。接下來,我們通過過表達NK4來探究其對Raji生物學行為的影響。結果顯示,上調NK4基因表達水平能夠明顯抑制NHL細胞生長進展。這暗示NK4在NHL細胞中作為抑瘤基因發揮著重要功能。
已有的研究證明,Sirtuin家族具有延緩細胞衰老和延長壽命方面的功能[10]。Sirt1有多種底物,如NF-κB、Ku70、p53、E2F1、TFR2、PPARγ、p73、BcL-XL、MyoD、AR、cI AP2和FOXO家族的轉錄因子。p53作為Sirt1底物的最具代表性的底物,被認為是一種腫瘤抑制因子。有研究報道,Sirt1表達與彌漫大B細胞淋巴瘤的不良預后相關。Ismail等的研究顯示p53的表達水平反映了侵襲性淋巴瘤亞型中有關基因突變的積累。因此,Sirt1/P53通路與抑瘤基因發揮功能密切相關。基于此,在本研究中,結果發現,與正常B細胞相比,Sirt1蛋白表達水平在NHL細胞中顯著上調,而p53蛋白表達水平顯著下降。這提示Sirt1/P53通路在NHL細胞中異常激活。然而,過表達NK4能夠抑制Sirt1/P53通路活性。另外,通過利用Sirt1/P53通路激活劑SRT1720筆者觀察到,激活Sirt1/p53信號通路能夠部分逆轉過表達NK4對NHL細胞生長進展的抑制作用。因此,NK4可能通過抑制Sirt1/p53通路從而抑制NHL細胞增殖、誘導NHL細胞凋亡。
總之,我們的結果提供了NHL中NK4過表達可能對削弱細胞生長進展具有十分重要的作用,這表明NK4在NHL進展中作為一種抗癌基因發揮作用,并可能作為一種新的潛在治療生物標志物影響下游Sirt1/p53信號通路。但本研究中不足之處在于僅在Raji細胞中對該機制進行驗證,未充分考慮該機制是否存在于其它NHL細胞系中。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
小讀者(2021年2期)2021-03-29 05:03:48
火花(2019年12期)2019-12-26 01:00:28
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
人大建設(2019年12期)2019-05-21 02:55:32
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
學苑創造·A版(2015年11期)2016-01-14 09:03:27
