睡蓮屬植物花粉形態研究
2023-08-31 09:53:52戴忠良徐迎春劉春貴孫春青
西北植物學報 2023年7期
曹 婧,戴忠良,徐迎春,劉春貴,徐 婷,孫春青,*
(1 江蘇丘陵地區鎮江農業科學研究所,江蘇句容 212400;2 南京農業大學,南京 210095;3 江蘇里下河地區農業科學研究所,江蘇揚州 225000)
睡蓮科(Nymphaeaceae)有8~9屬約100個種,其中睡蓮屬(Nymphaea)有50余種[1]。睡蓮屬植物根據心皮附屬物的有無可分為Nymphaea亞屬、Brachyceras亞屬、Anecphya亞屬、Lotos亞屬和Hydrocallis亞屬[2]。其中,Nymphaea亞屬包括8種,在北半球的溫帶地區分布;Brachyceras亞屬包括14~16個種,在熱帶和亞熱帶地區分布;Anecphya亞屬包括7~10個種,主要在澳大利亞和新幾內亞分布;Lotos亞屬包括3~4個種,在熱帶地區分布;Hydrocallis亞屬包括14個種,在熱帶地區分布[3]。盡管有些睡蓮亞屬的種數量少,但是睡蓮屬植物在生活史、心皮形態、繁育系統、傳粉生物學等方面表現出顯著的多樣性[4-8]。
目前,從形態學[9]、解剖學[10]和分子生物學[3]已對睡蓮屬植物的系統分類和進化關系進行了相關研究,但仍存在互相矛盾的觀點[2],因此,有必要從孢粉學角度研究睡蓮屬植物的系統進化關系。更重要的是,睡蓮屬處于被子植物系統進化的關鍵位置[11],并且睡蓮屬植物在生態和形態特征上具有多樣性,因此,睡蓮屬植物孢粉學的特征對于研究早期被子植物生殖特征進化至關重要。但是先前的研究僅對睡蓮屬植物的數種花粉特征開展研究。例如,楊梅花等僅對4種睡蓮的花粉形態進行研究,結果表明雪白睡蓮(N.candida)的花粉形態特征在睡蓮屬植物中較原始[12]。
同樣地,Zini等僅對Hydrocallis亞屬的3個種花粉粒發育特征和超微結構進行研究,發現Hydrocallis亞屬花粉的花粉壁較光滑,花粉壁無顆粒、棒狀、疣狀等紋飾[10]。現有睡蓮屬植物孢粉學研究缺乏多個亞屬關鍵的花粉形態特征資料,因此需要對睡蓮屬植物未報道的花粉形態特征進行研究,并對其進行全面比較分析。此外,耐寒睡蓮與熱帶睡蓮跨亞屬雜交的母本大多選擇香睡蓮塊莖類型的耐寒睡蓮[13],因此本研究對香睡蓮的花粉活力進行評價,因為在睡蓮雜交組合中母本自交結實率高更易獲得跨亞屬雜交種[14],而花粉生活力可直接影響自交結實率,以期為耐寒睡蓮與熱帶睡蓮的跨亞屬雜交提供理論依據。
本研究利用掃描電鏡對4個亞屬的18種或品種睡蓮花粉進行形態觀察,測量和分析了花粉的形態特征,采用離體萌發法對‘彼得’的花粉活力進行測定,研究結果可為睡蓮屬植物的系統進化提供證據,也可為睡蓮雜交育種親本的選擇提供依據。
1 材料和方法
1.1 供試材料
供試材料均保存在江蘇丘陵地區鎮江農業科學研究所水生植物資源保存圃。本試驗選擇4個亞屬的18種或品種睡蓮的具體性狀特征見表1。2021年9月,待睡蓮陸續開放,收集供試睡蓮的成熟新鮮花粉。
1.2 方 法
1.2.1 睡蓮花粉形態學研究將睡蓮花藥分別置于2.5%戊二醛(用pH 7.2的0.1 mol/L的磷酸緩沖液配制)溶液中4 ℃固定4 h固定8 h以上。
用生理鹽水進行3次沖洗,每次10 min;再經50%乙醇→60%乙醇→70%乙醇→80%乙醇→90%乙醇→100%乙醇梯度脫水,每個處理15~20 min,將樣品置于4 ℃過夜。
再將脫水過夜處理的花粉進行乙酸異戊酯置換、二氧化碳臨界點干燥[15-17]。脫水和干燥后用雙面膠將花粉均勻地粘在金屬臺上,真空噴金、鍍膜后用Hitachi S-3000N型掃描電鏡(日本Hitachi公司)進行觀察。
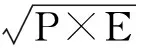

運用系統聚類組間聯接的分析方法,根據平方歐氏距離測量,最終獲得18種或品種睡蓮屬植物花粉聚類分析圖,從而推斷其系統進化規律以及它們的親緣關系[20]。
1.2.3 睡蓮花粉生活力研究采用離體培養法對睡蓮品種‘彼得’進行花粉生活力測定。用吸管吸取2~3滴柱頭液于載玻片作為培養液,選擇第2天開放的耐寒睡蓮花朵,于上午9:00用鑷子沾取花粉播種于載玻片的柱頭液上,后將載玻片放入墊有濕潤濾紙的培養皿中,置于恒溫光照培養箱,28 ℃培養4~6 h。
顯微鏡下觀察花粉萌發情況,以花粉管長度大于或者等于花粉粒長度作為花粉管成功萌發的標準[21]。
隨機選擇3個視野,統計花粉粒數量及萌發數量,每個視野隨機抽取3粒萌發的花粉粒,測量花粉管長度。
1.3 數據統計分析
數據采用Excel 2003進行統計和分析,圖片處理采用Adobe Photoshop CS5。
2 結果與分析
2.1 花粉粒形態特征
利用掃描電鏡對18種或品種睡蓮屬植物的花粉形態進行觀察,發現18種或品種睡蓮屬植物的花粉都是單粒花粉。
如圖版Ⅰ所示,18種或品種睡蓮屬植物花粉粒極軸/赤道軸比值范圍為:0.62~0.89,形態均為近球體或扁球體,極面觀為圓形或近圓形,赤道面觀為圓形、長圓形或近圓形;其中,花粉粒極軸長度范圍:15.1~27.9 μm,赤道軸長度范圍:24.5~32.8 μm,巨花睡蓮花粉粒最大,體積指數為30.3;埃及白花粉粒最小,體積指數為19.2;其余花粉粒體積指數在26左右(表2)。此外,埃及白睡蓮、喀麥隆睡蓮、印度紅睡蓮、墨西哥黃睡蓮的花粉形態具有異極性。例如:墨西哥黃睡蓮花粉的遠極面光滑,而近極面為瘤狀紋飾。
埃及藍睡蓮的萌發溝為單槽萌發溝,而其他17種睡蓮屬植物都為環槽萌發溝,且與赤道平行(圖版Ⅰ和Ⅱ)。
栽培品種‘達爾文’和‘佛羅里達’為遠級環槽類型,其他15個種或品種屬于赤道環槽或者近赤道環槽。
香睡蓮、‘達爾文’、‘火冠’、‘亞克’、‘佛羅里達’和‘征服者’的花粉萌發溝較淺。Brachyceras亞屬的藍星睡蓮萌發溝內有環形嵴。
18種或品種睡蓮屬植物花粉外壁紋飾存在一定差異,類型也比較豐富(表3),大部分為小顆粒狀、瘤狀和小柱狀,少數具有長條狀、微波紋起伏狀和網紋狀(圖版Ⅰ、Ⅱ、Ⅲ)。Lotos亞屬喀麥隆睡蓮、埃及白睡蓮的花粉外壁表面均較為平滑,附屬物少;而印度紅睡蓮的花粉外壁表面可觀察到顆粒狀紋飾。巨花睡蓮、小花睡蓮和睡蓮子午蓮的花粉外壁為顆粒狀凸起紋飾。
藍星睡蓮和一些具有香睡蓮塊莖的種和品種(香睡蓮、‘亞克’、‘火冠’)花粉外壁具有顆粒狀凸起、棒狀和刺狀紋飾,只是紋飾的密度和高度有差異。
2.2 聚類分析
聚類分析結果如圖1所示,18種或品種睡蓮屬植物可分為4個類群。第1類群有香睡蓮、白睡蓮、墨西哥黃睡蓮和睡蓮子午蓮4個原生種以及‘寬瓣白睡蓮’、‘達爾文’、‘火冠’、‘亞克’、‘佛羅里達’、‘征服者’和‘彼得’7個栽培種,其花粉主要為圓球形,幾乎每個品種都有多個外壁紋飾類型,P/E值在0.70~0.84之間。第2類群有藍星睡蓮、小花睡蓮和埃及藍睡蓮3個原生種,花粉為扁球形,其P/E值在0.70~0.73之間,體積指數相近(22.0~23.0),均有瘤狀附屬物。第3類群僅有巨花睡蓮,花粉形態為圓球形,P/E值為0.85,外壁紋飾為微波紋起伏狀,極軸、赤道軸等各項指標數據均較大。第4類群有印度紅睡蓮、喀麥隆睡蓮和埃及白睡蓮3個原生種,其P/E值在0.62~0.89之間,這3個原生種的體積指數、溝長遠小于其他種(品種),表面紋飾單一,與其他3個類群又明顯區別。
2.3 花粉生活力
‘彼得’花粉在28 ℃恒溫光照培養箱生長6 h后,花粉萌發比例高達95%,如圖2所示,睡蓮花粉粒生長出花粉管,平均長度約為260 μm,花粉管生長狀態好。因為睡蓮授粉后花粉是落到柱頭液中,經過長期進化選擇,柱頭液內含豐富的氨基酸、微量元素[21],為睡蓮花粉發育提供了最有利的條件,使得花粉具有較好的育性。
3 討 論
3.1 睡蓮屬植物花粉形態多樣性及分類學意義
18種或品種睡蓮屬植物的花粉形態具有豐富的多樣性。花粉形態的聚類分析結果可將18種或品種睡蓮屬植物分為4個類群,這一結果與基于核糖體DNA的ITS序列[3]和葉綠體基因[2]所建立的系統分類一致。18種或品種睡蓮屬植物的花粉萌發溝除了埃及藍睡蓮是單槽的,其余均為環槽形的,并且環槽形花粉溝的位置在花粉遠極端和赤道間變化,而以前的研究結果都認為睡蓮屬植物的花粉萌發溝都是環槽類型的[22-23]。Brachyceras亞屬的3個種都有明顯區別于其他種的花粉形態特征,這為這些種的鑒定提供了依據。Anecphya亞屬的澳大利亞巨睡蓮的花粉體積最大,可能由于其所處的生態環境影響所致,這些特征也是鑒定這些種的可靠依據。Lotos亞屬的花粉外壁紋飾較光滑,且花粉形態差異小,這與前人的研究結果[24]一致,可以利用花粉形態特征從睡蓮屬植物中將Lotos亞屬區分開來。Nymphaea亞屬中具有香睡蓮塊莖的種和品種的花粉外壁紋飾相似度較高,一般都具有棒狀、刺狀和顆粒狀紋飾,只是紋飾的密度和高度有差異,這一特征可以區分大多數睡蓮屬植物中具有香睡蓮血統的種和品種。根據上述特征,我們推斷Nymphaea亞屬的栽培品種‘亞克’、‘火冠’、‘佛羅里達’和‘征服者’都可能是香睡蓮的雜交品系。此外,以Brachyceras亞屬藍星睡蓮、Nymphaea亞屬香睡蓮為代表,其花粉粒表面小顆粒突出且數量多,且都具有棒狀、刺狀紋飾,這些特征可對花粉起到較好的保護作用,有利于結實和發育,因此這些種作為親本雜交成功率大。
3.2 睡蓮屬植物花粉形態特征的系統進化趨勢
花粉的形態特征,特別是花粉外壁紋飾、萌發溝等特征,對于了解睡蓮屬植物系統演化、親緣關系及系統分類均有重要意義。花粉外壁表面紋飾特點可以反映植物進化的一般規律[25],花粉外壁表面較光滑的較為原始,其次為顆粒狀凸起,再到棒狀、刺狀,最終演化為條紋狀及網狀[26]。本研究中,Lotos亞屬中喀麥隆睡蓮、埃及白睡蓮和印度紅睡蓮的花粉形態相似,但三者的花粉外壁紋飾不同,其中喀麥隆睡蓮、埃及白睡蓮的花粉外壁表面均較為平滑,附屬物少,說明它們在睡蓮屬植物中較為原始、進化程度較淺,而印度紅睡蓮的花粉外壁表面可觀察到有顆粒狀附屬物存在,說明有進化發展的趨勢;Zini等[23]發現Hydrocallis亞屬的3個種花粉壁較光滑,表明Hydrocallis亞屬和Lotos亞屬的種在睡蓮屬植物中最原始,并且這2個亞屬的種可能有共同的起源[24]。Anecphya亞屬巨花睡蓮、Brachyceras亞屬的小花睡蓮、Nymphaea亞屬的睡蓮子午蓮的花粉外壁都有顆粒狀凸起,Brachyceras亞屬的藍星睡蓮、Nymphaea亞屬的香睡蓮花粉外壁的顆粒狀凸起更加明顯,且藍星睡蓮和香睡蓮粉外壁在顆粒狀凸起的基礎上演變出棒狀和刺狀紋飾,說明睡蓮屬植物是不斷進化的,其中Nymphaea亞屬的香睡蓮在睡蓮屬植物中進化程度較高。此外,Nymphaea亞屬中的墨西哥黃睡蓮花粉的遠極面是光滑的,而近極面具有顆粒狀凸起,說明墨西哥黃睡蓮可能是睡蓮屬植物進化中的過渡類型。
關于睡蓮屬植物環槽萌發溝的起源一直有爭議,現有兩種假說。第一種假說認為環槽萌發溝起源于萍蓬草屬(Nuphar)的單槽萌發溝[27]。本研究中埃及藍睡蓮的萌發溝為單槽萌發溝,而其他17種睡蓮屬植物都為環槽萌發溝,根據第一種假說,筆者推斷埃及藍睡蓮的萌發溝為睡蓮屬植物的原始形態,而環槽萌發溝是睡蓮屬植物的進化形式。第二種假說認為睡蓮屬植物環槽萌發溝是單孔形式的,環槽是單孔結構的擴展,花粉粒的整個遠極端半球可看作是單孔的修飾部分[27]。根據第二種假說,如果遠極端半球可以看作是修飾蓋,那遠極端與近極端的超微結構不同[28]。在本研究中,Nymphaea亞屬的白睡蓮和墨西哥黃睡蓮的遠極面和近極面的超微結構不同,這一結果支持了第二種假說,但本研究其他睡蓮屬植物不存在這種現象。睡蓮屬植物萌發溝的進化歷程可能為:遠極單槽→遠極環槽→赤道環槽[24]。根據這一進化歷程,本研究中埃及藍睡蓮的萌發溝屬于最原始類型,Nymphaea亞屬白睡蓮和睡蓮子午蓮屬于較原始類型,其余原生種萌發溝都是較進化類型(赤道環槽)。此外,栽培品種中‘佛羅里達’和‘達爾文’屬于遠極環槽,可能是由于品種間雜交的原因。
3.3 睡蓮屬植物花粉形態特征的親緣關系分析
花粉形態特征的穩定性強,可用于植物種間親緣關系分析[29]。18種或品種睡蓮屬植物的聚類分析結果表明:Nymphaea亞屬和Brachyceras亞屬中的種和栽培種間的親緣關系較近;Lotos亞屬中印度紅睡蓮和喀麥隆睡蓮親緣關系較近,而它們與埃及白睡蓮的親緣關系較遠;Nymphaea亞屬和Brachyceras亞屬間親緣關系較近,許多育種者已經獲得了這2個亞屬的雜交種[13,30];Brachyceras亞屬和Anecphya亞屬親緣關系較遠,但是我們已經得到了這2個亞屬的雜交種,說明利用花粉形態特征分析睡蓮屬植物的親緣關系有一定的誤差。Nymphaea亞屬和Anecphya亞屬、Lotos亞屬與其他3個亞屬間的親緣關系遠,至今無育種者獲得這些亞屬間的雜交種。
綜上所述,花粉形態特征,特別是花粉萌發溝類型、外壁紋飾等特征可為睡蓮屬植物種間分類、亞屬間以及種間親緣關系、系統進化規律提供關鍵的證據。但孢粉學的研究對分析睡蓮屬演化、親緣關系及系統分類具有一定的局限性,仍需通過形態學、解剖學和分子生物學等手段深入探討和分析,以便更深層次地了解睡蓮屬植物系統演化過程。
猜你喜歡
數學小靈通·3-4年級(2024年2期)2024-05-15 02:02:28
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:36
瘋狂英語·新策略(2019年10期)2019-12-13 08:43:28
當代陜西(2019年10期)2019-06-03 10:12:04
數學小靈通·3-4年級(2017年9期)2017-10-13 08:10:54
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
