滇水金鳳花斑形成相關基因IuMYB114和IuMYB36的克隆及表達分析
2023-08-28 06:01:54陶宇蝶孟丹晨黃美娟魏春梅趙秋燕周敏瞿素萍黃海泉
山東農業科學 2023年2期
陶宇蝶 孟丹晨 黃美娟 魏春梅 趙秋燕 周敏 瞿素萍 黃海泉


摘要:MYB基因作為植物中最龐大的一個基因家族,在花斑及色素形成、生長發育等過程中發揮著重要作用。本研究以滇水金鳳花器官為材料,獲得2個MYB基因,分別命名為IB114和IuMYB36,其cDNA分別為315 bp和876 bp,分別編碼104個和151個氨基酸。生物信息學分析顯示:二者均不具有內含子且均為親水性不穩定蛋白,IuMYB114基因屬于MYB超家族,推測IuMYB36屬于新的R2R3-MYB轉錄因子亞組;IuMYB114和IuMYB36的氨基酸序列與其他物種的同源性均在64%和50%左右;二者均分別與各自的同源序列聚在一起,且處在兩個不同分支。qRT-PCR分析發現兩個基因在滇水金鳳斑區和非斑區中均有表達,但在斑區表達量顯著高于非斑區,IuMYB114和IuMYB36基因斑區表達量分別為非斑區的12.83倍和9.88倍,推測兩個基因在滇水金鳳花斑形成中發揮了重要的調控作用。本研究結果為后續探討滇水金鳳花斑形成的分子調控機理以及進行鳳仙花花色改良等方面研究提供了一定的理論依據。
關鍵詞:滇水金鳳;花斑形成;MYB基因;基因克隆;表達分析
中圖分類號:S681.I:Q78 文獻標識號:A 文章編號:1001-4942(2023)02-0001-08
花色是被子植物花器官的重要表型性狀之一,在提高植物抗逆性、吸引傳粉者、維持繁衍進化基石等方面發揮著重要作用。花斑一般指在花瓣或花萼上位置、大小和形態基本固定的斑塊,是被子植物一個獨特的表型特征,是人工選擇創新、植物進化和自然選擇的綜合結果,能極大地豐富植物的觀賞價值,是現代花卉業的重要研究領域。因此,揭示花斑形成的分子機理對花色改良具有重要意義。
植物花斑的形成是由于花色素基因在花瓣或其他成花部位上差異表達,且調控基因可以調節花色素合成催化酶基因在不同區域的表達水平,進而導致花斑的形成。MYB基因作為植物中最龐大的一個基因家族,在花斑及色素形成、生長發育等過程中發揮著重要作用。已在多種植物中發現調節花色索苷合成及與色斑形成相關的MYB轉錄因子,且以R2R3類型為主,也有單一R重復結構的MYB轉錄因子。如AuMYB75、At-MYB90、AtMYB113參與花青素的生物合成;在蝴蝶蘭中,PeMYB2調控唇瓣的全紅著色,PeMYB11調控唇瓣的紅色斑點,PeMYBI2調控唇瓣紋理的形成;R2R3-MYB轉錄因子LhMYB6、IhMYB12和IhMYB12-Lat分別在百合的凸起式斑點、飛濺式斑點和非斑區表達;cjMYBI基因可能在紅色山茶品種的花色苷合成途徑中起關鍵作用;FtMYB23提高了苦蕎原花青素的合成與積累;在月季紅花突變體中,RhMYB基因正調控花青素的合成。劉盛雨研究發現,MYB114轉錄因子可能在血橙果實成熟過程中介導藍光對多甲氧基黃酮和花青素的合成調控;Yao等提出PyMYB114與已報道的PyMYB10有著色增強的效應,并在草莓果實和煙草葉片中得到驗證:趙英提出SmMYB36的同源基因首次出現在洛磯山耬斗菜中,過表達SmMYB36的轉基因材料可以提高丹參酮類物質而抑制酚酸類物質。但關于MYB36基因調控花斑形成的相關研究尚未見報道。
滇水金鳳(Impatiens uliginosa)為中國特有的鳳仙花科鳳仙花屬一年或多年生草本植物,花色艷麗,以紅色為主,花形獨特,旗瓣圓形,花期長,具有較高的園林應用價值。目前關于滇水金鳳的研究主要集中于種子萌發特性、花發育調控基因克隆、花色調控基因及金屬元素測定等方面,尚未見關于其花斑的研究報道。本研究從斑色系滇水金鳳花瓣中克隆得到IuMYB114和IuMYB36基因,并分析了其序列結構、系統發育和表達模式,為研究滇水金鳳花斑形成的分子調控機理以及鳳仙花花色改良等提供一定的基礎數據和理論依據。
1材料與方法
1.1試驗材料
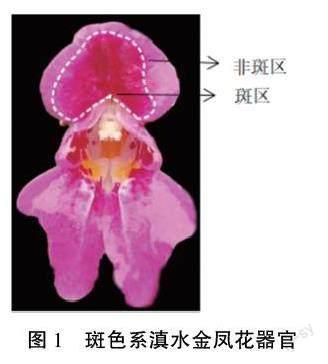
供試斑色系滇水金鳳于2020年10月取自昆明市尋甸縣清水海,于滇水金鳳盛花期采集樣品(圖1),迅速放于液氮中,并帶回實驗室保存于-80℃冰箱中備用。
1.2總RNA的提取與IuMYB114和IuMY836基因的克隆
以滇水金鳳整個花器官為試驗材料,采用RNA提取試劑盒(OMEGA)提取總RNA;RNA逆轉錄采用逆轉錄試劑盒(北京全式金生物技術股份有限公司),-20℃保存備用。根據轉錄組數據設計擴增全長的特異性引物(表1)。以RNA逆轉錄合成的cDNA第一鏈作為模板進行IuMYB114和IuMYB36基因全長cDNA擴增。回收產物并進行克隆,送上海生工生物工程有限公司測序。
1.3滇水金鳳花斑IuMYB114和IuMYB36序列分析
采用ExPasy-ProtParam在線軟件進行基因編碼蛋白質的基本理化性質分析:運用TMHMM2.0對蛋白進行跨膜結構域的預測分析:利用NCBI的CDD工具預測目的基因結構域:運用SMART對蛋白結構域進行分析,蛋白質的二級結構預測采用SOPMA在線軟件進行,蛋白質的三級結構預測采用SWISS-MODE在線軟件進行:運用NCBI中的Protein BLAST工具尋找同源序列,借助NCBI BLAST、DNAMAN和MEGA 11進行序列分析和系統進化樹構建。
1.4IuMYB114和如MYB36基因的表達模式
按照前述方法分別提取滇水金鳳旗瓣斑區和非斑區2個部位(圖1)的RNA,反轉錄合成cD-NA,設計qRT-PCR引物(表2),以滇水金鳳Ac-tin1作為內參基因,進行qRT-PCR試驗。每個樣本設置3個生物學重復。將非斑區的表達水平設為對照,與斑區的進行比較。
2結果與分析
2.1滇水金鳳IuMYB114和IuMYB36基因的克隆
通過PCR擴增,獲得luMYB114與IuMYB36基因的cDNA全長,分別為315 bp和876 bp(圖2),分別編碼104個和151個氨基酸。對這兩個MYB基因的內含子和外顯子進行分析發現兩者均不具有內含子。
2.2IuMYB114和IuMYB36基因的序列結構分析
luMYB114、IuMYB36蛋白均為親水性不穩定蛋白,均不具有跨膜結構域,均屬于MYB超家族蛋白成員(圖3、表3)。預測目的基因結構域發現IuMYB114具有MYB轉錄因子典型的SANT結構域,IuMYB36不具有保守結構域(圖4)。
2.3IuMYB114和IuMYB36基因編碼蛋白的二、三級結構預測
luMYB114和luMYB36蛋白的二級結構均以無規則卷曲為主,說明其結構不穩定。蛋白質三級結構預測結果(圖5)顯示,luMYB114是以轉錄因子WER為模型,屬于R2R3型亞類成員,覆蓋率為53.19%,序列相似性為47%:IuMYB36是以涂層蛋白為模型,覆蓋率為27.08%,序列相似性為33%。
2.4luMYB114和luMYB36的氨基酸序列同源性分析及系統進化分析
分別將luMYB114和IuMYB36的氨基酸序列與其它植物的氨基酸序列進行對比,結果(圖6)顯示,IuMYB114與芝麻(XP_011100166.1)、歐洲李(XP_034208044.1)等物種的MYB同源性較高,同源性均達64%左右;luMYB36與常綠越橘(KAH7854734.1)、黃芩(AKA59789.1)和藍果樹(KAA8518714.1)等的同源性較高,同源性均在50%左右。通過構建系統進化樹,發現luMYB114和IuMYB36均分別與各自的同源序列聚在一起,各處在兩個分支,與luMYB114親緣關系最近的是旱金蓮,而與IuMYB36親緣關系最近的是除蟲菊(圖7)。
2.5如IuYB114和IuMYB36基因在滇水金鳳旗瓣中的表達模式分析
通過對兩個基因在滇水金鳳旗瓣不同部位的表達分析(圖8)發現,luMYB114與luMYB36在斑區和非斑區均有表達,但斑區表達量顯著高于非斑區,分別為非斑區的12.83倍和9.88倍,與轉錄組分析結果相符。推測IuMYB114和IuMYB36可能在盛花期存在一定的正調控作用從而使花斑顯現;另外,兩個基因的表達模式和表達量相似,推測兩者在斑區具有一定的協同作用。
3討論與結論
MYB轉錄因子集特異性和多樣性于一體,在花色素中發揮著重要作用,是花斑形成的關鍵基因。最早從玉米中克隆出第一個調節花色素合成的單一型MYB基因——C/基因,隨后從巨峰葡萄中克隆出WMYBA基因,能誘導葡萄果實上紅紫色斑點的產生。近幾年,關于MYB轉錄因子組織表達特異性導致花斑出現的機理逐漸成為研究熱點,但未見其在滇水金鳳花斑形成中作用的相關研究報道。
本研究成功從滇水金鳳花器官中克隆得到2個cDNA全長分別為315 bp和876 bp、分別編碼104個和151個氨基酸的MYB基因,命名為IuMYB114和IuMYB36。經生物信息學分析,IuMYB114和IuMyB36均沒有內含子,這與從蓖麻莖稈中克隆的RcMyB114、從丹參中克隆的SmMyB36分析結果一致,且二者均屬于親水不穩定蛋白,二級結構均以不穩定的無規則卷曲為主:luMYB114三級結構以轉錄因子WER為模型,屬于R2R3型亞類,且具有典型的MYB轉錄因子SANT結構域,這與在甘藍型油菜中克隆的RFBnMYB114的分析結果一致;IuMYB36不具有保守結構域,與丹參SmMYB36基因的分析結果一致,推測其屬于新的R2R3-MYB轉錄因子亞組。同源性分析發現,滇水金鳳的luMYB114與芝麻(Sesamum,indicum)和歐洲李(Prunus dulcis)等的MYB同源性均達64%左右,IuMYB36與常綠越橘(Vaccinium,darrowi)、黃芩(Scutellaria playfairii)等的MYB同源性均達50%。左右;系統進化分析發現IuMYB114和IuMYB36各處在兩個分支。
植物花斑的出現主要依賴花青素在時間和空間上的受限沉積,由于MYB基因是調節花青素合成和積累的關鍵因子,研究MYB基因對花斑形成的作用具有重要意義。研究表明,在牡丹中R2R3-MYB轉錄因子PsMYB12協同WD40和bHLH轉錄因子,共同調控牡丹花瓣底部花斑的形成;文心蘭中得到的OgMYB1基因通過控制OgCHI和OgDFR兩個結構基因的表達進而調控花斑的形成:外源aMYB基因對中國水仙花色素苷代謝途徑產生了影響,且誘導了結構基因的表達,使其花瓣發生了顏色的變化:PcMYB114是導致梨果皮紅色突變的關鍵基因:ATMYB114作為激活劑參與了擬南芥中原花青素、類黃酮或花青素的生物合成;PyMYB114與PybHLH3、PyERF3之間相互作用形成調控復合體,促進梨果實花青素的積累;MdMYB114是蘋果花青素生物合成和運輸的積極調節因子。結合以往研究,推測IuMYB114可能與滇水金鳳的花色苷生物合成有關。另外,有研究表明,CsMYB36可以調節黃瓜的黃綠色果皮著色;SIMYB36-1基因過表達后對番茄根系內皮層凱氏帶形成具有影響;MIB36控制擬南芥側根原基邊界,從而調控根從增殖到分化的轉變。本研究發現luMyB114和IuMYB36在滇水金鳳旗瓣斑區和非斑區均有表達,但在斑區的表達量顯著高于非斑區,這與MYB114L在‘鳳丹牡丹花色變化過程中的表達分析結果一致。推測IuMYB114可能在滇水金鳳盛花期的斑區中起一定的正調控作用,而IuMYB36可能在斑區顏色形成中發揮著一種新功能,從而對花斑的形成具有一定調控作用:因二者表達模式和表達量相似,推測它們具有一定的協同作用,但具體功能尚有待進一步試驗驗證。
本研究結果可為后續進行IuMYB114和IuMYB36基因的功能研究、探究其對滇水金鳳花斑形成的調控機制以及鳳仙花花色改良等提供一定的理論依據。
