蛋雞免疫及營養性免疫調控機制的研究進展
2023-08-27 15:35:38康佳佳高振東何洋施紅梅劉永王坤葛長榮
江蘇農業科學 2023年14期
康佳佳 高振東 何洋 施紅梅 劉永 王坤 葛長榮

摘要:幾十年來,我國蛋雞產業都在穩定發展,標準化規模化的發展趨勢一直處于畜牧業前列水平。尤其是自主創新品種,近幾年成為畜牧業中的領軍者。再加上政府出臺的各項扶持政策,使得蛋雞產業處于我國乃至世界領先地位,但是免疫力與生產性能兩者無法同時達到最佳水平,是導致蛋雞產業停滯不前的根本原因之一。目前為止,關于禽類免疫方面的系統介紹較為匱乏。因此,全面總結禽類免疫特征,了解禽類最新免疫進展,試圖從中找到免疫力與生產性能同時達到最佳水平的平衡點,持續高效發展蛋雞產業,具有十分重要的實際經濟價值。本文概述了蛋雞免疫、免疫性指標、免疫抑制性疾病的作用機理及影響因素,簡單介紹了自身免疫調控,詳細闡述了營養性免疫調控機制與策略,旨在為未來蛋雞行業更加高效、綠色經濟、穩步發展提供新的參考。
關鍵詞:蛋雞;免疫指標;免疫抑制性疾病;營養性免疫調控機制;營養性免疫調控策略
中圖分類號:S852.4文獻標志碼:A
文章編號:1002-1302(2023)14-0032-11
我國蛋雞產業發展迅速,綜合生產能力顯著增強,消費市場不斷拓寬,進入一個資本驅動、生態驅動、新技術驅動的全新時代[1]。但受新冠肺炎、禽流感以及“雙碳”目標的影響,2021年禽蛋產量約為2 682.6萬t,比2020年約下降9.0%。隨著疫情逐漸好轉,預計2022年雞蛋整體供需相對平衡、禽蛋產量銷售情況將有所提升[2]。然而,由于育種弊端,蛋雞生產力和免疫力之間始終呈負相關,產蛋量最大化的育種目標導致商品蛋雞免疫力大幅下降[3-4]。蛋雞免疫力低下嚴重影響了蛋雞的健康,威脅到蛋雞養殖業的持續發展。如2020年10月至2021年2月暴發的全球性禽流感事件,導致大量免疫力低下蛋雞患病被捕殺,致使蛋雞產蛋量迅速下降,供不應求[5]。一系列問題的出現,讓筆者逐漸意識到免疫力、生產性能、經濟效益三者直接相關。如何高效解決由于免疫能力不足而阻礙禽類生產性能發揮,降低經濟效益的現象,成為當代急需解決的一個棘手問題。通過基因篩選培養出新的優良品種,從根本上解決免疫力的問題,是一個不錯的辦法,但持續時間過長。因此,本研究重點介紹了蛋雞的免疫及營養性免疫調控機制,旨在表明在優良免疫相關基因高度選擇的前提下,再依靠蛋雞完整的免疫系統與正常的免疫功能,進行營養性免疫調控,是有效預防和治療疾病的關鍵,能保證蛋雞產能最大化。
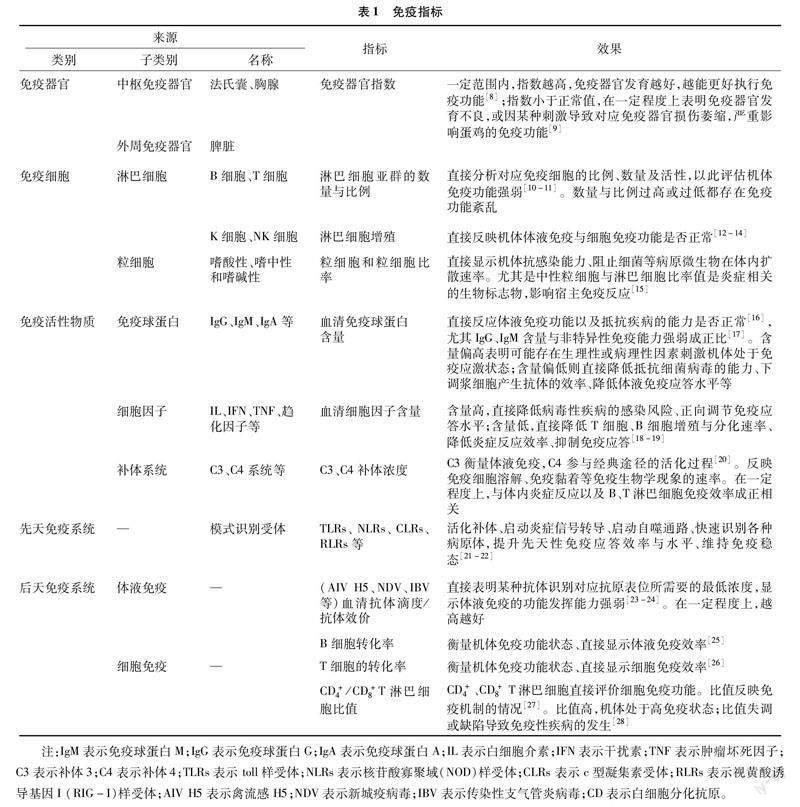
1 蛋雞的免疫
1.1 免疫指標
免疫系統是一個整體,與機體其他系統相互協調,具有識別和排除抗原性異物、維持機體內環境穩定和生理平衡的功能[6]。遺傳、營養、環境、養殖場生物安全措施等與家禽飼養管理相關的各個方面,都會影響雞群的健康,而免疫系統的完整性是有效預防和對抗疾病的關鍵所在。因此,可將免疫系統作為蛋雞健康的指標來反映蛋雞場的管理是否合理(表1)[7]。
1.1.1 免疫器官、免疫細胞指標及其影響因素
如表1所示,免疫器官指標主要是胸腺、脾臟、法氏囊指數。免疫細胞指標主要是B、T、K、NK等淋巴細胞的數量及比例,嗜中性、嗜酸性、嗜堿性粒細胞的比率等。受環境溫度和食物成分影響,免疫器官和免疫細胞指標會有變化,在一定程度上可以反映機體免疫能力。例如,受金霉素與博落回提取物的共同刺激,6~17周齡的海蘭褐蛋雞,胸腺指數、脾臟指數、法氏囊指數分別增長3.4%~9.6%、 1.5%~11.26%、0.69%~22.64%,直接提升免疫器官質量,促進免疫細胞分化,提升免疫能力[29]。14周齡伊莎褐育成雞,24 ℃環境下,淋巴細胞(169.93×109/L)、粒細胞(24.56×109/L)、粒細胞比率(12.10%)相比于其他溫度含量最高、免疫性能最強[30];7日齡海蘭褐公雛對照組皮下注射 100 mg/mL 的滅菌生理鹽水,試驗組皮下注射等量刺五加多糖(acanthopanax senticosus polysaccharide,ASPS),21 d后CD4+/CD8+T淋巴細胞比值分別為0.84、1.10,顯著增長30.95%,表明免疫細胞含量增長,可能存在間接促進脾臟免疫功能發揮,進一步增強蛋雞免疫的效果[31]。
1.1.2 免疫因子指標及其影響因素
血清中免疫抗體除了IgA、IgG、IgM等體液免疫的重要組成物質外,還存在IL、IFN、TNF等細胞因子,正向調節免疫與炎癥反應。營養物質可直接促進免疫因子含量提升,如20周齡海蘭褐蛋雞飼喂1.0%女貞子粉,促使血清IgG、IgA、IgM由原來的0.21、0.12、0.12 g/L,分別提升33.33%、33.33%、16.67%[32];呂新彪在基礎日糧中添加1.5%二至丸,結果發現72周齡海蘭灰蛋雞血清中IgA、IgG、IgM整體含量提升30.4%、60.86%、27.90%,IL-1β、IL-2、IL-6 分別提升13.54%、73.43%、27.01%,直接增強蛋雞免疫性能[33]。
1.1.3 其他免疫指標及其影響因素
激活補體系統(complement system,CS)后形成抗菌蛋白復合物和多種信號分子,進一步刺激免疫應答。研究證實,雞先天性免疫系統分泌的可溶性蛋白、甘露糖結合凝集素(mannose binding lectin,MBL),結合絲氨酸蛋白酶經過凝集素途徑激活補體系統,直接激活免疫反應[34];飼糧中添加2 000 mg/kg苜蓿粗多糖,30周齡海蘭褐產蛋雞體內C3(230.09 μg/mL)增長50.53%,直接介導炎癥反應,增強B淋巴細胞、T淋巴細胞免疫,提升免疫效果[35]。抗體滴度直接衡量某種抗體識別特定抗原的能力。研究表明,30周齡的海蘭褐商品蛋雞,AIV H5、NDV的抗體滴度分別為log2 6.87、log2 6.27,在基礎日糧中添加0.3 g/kg的大蒜素,促使對應抗體滴度整體含量提升22.27%、41.47%,直接增強海蘭褐蛋雞對AIV H5、NDV的免疫水平[36];同時也有研究顯示,20周齡羅曼褐殼蛋雞IBV、NDV的抗體滴度是0.65、497.78 ng/L,在基礎日糧中添加8%的金針菇菌渣,促使相應抗體滴度含量提升22.27%、41.47%,直接增強羅曼褐殼蛋雞對IBV、NDV的免疫水平[37]。模式識別受體(pattern recognition receptor,PRR)是先天性免疫反應中免疫受體的代表,表明細胞損傷和疾病相關分子模式的宿主特征分子,使先天性免疫反應高效表達[38]。研究證實,清道夫受體(scavenger receptors,SR)CD36通過識別外來或修飾過的自我抗原,啟動不同細胞激活機制,在巨噬細胞和B細胞中高表達,調控B細胞自噬,影響免疫[39];另有研究表明,RLRs家族中的LGP2(laboratory of genetics and physiology 2),對病毒有一定的抑制作用,LGP2可以識別并與IBV的RNA結合;LGP2作為細胞質中的RNA傳感器,遇到病毒刺激時,能快速識別致病RNA,產生抗病毒免疫反應;并通過反式激活反應RNA結合蛋白TRBP(trans-activation response RNA-binding protein)的協同作用,抑制IBV在細胞內的增殖,防止感染[29]。
目前大量研究常常將蛋雞免疫器官中的法氏囊、胸腺、脾臟等免疫器官指數;免疫細胞中T、B淋巴細胞、各種粒細胞等含量占比;免疫活性物質IgA、IgG、IgM等免疫球蛋白;IL、IFN、TNF等細胞因子含量;補體系統、抗體滴度、模式識別受體等其他信號分子作為反應蛋雞免疫能力強弱、是否健康的指標。一旦免疫系統紊亂、免疫力喪失,就會產生免疫抑制性疾病。
1.2 免疫抑制性疾病
病原體以宿主免疫器官或免疫細胞為靶點,使免疫系統發生非暫時性或難以修復性損害的疾病稱為免疫抑制性疾病。根據不同影響因素,免疫抑制性疾病分為不同類型,見表2。飼料自身營養不足、管理不當引發霉變以及營養不合理引發應激,是造成免疫抑制性疾病的重要因素之一。
目前,研究顯示人們常采用大量藥物治療免疫抑制性疾病,導致病菌耐藥性顯著增加、雞體內藥物殘留量逐漸增多,影響人類健康。對此,后續急需開展采取各種免疫調控措施解決疾病問題的研究。
2 免疫調控
免疫調控包括自身免疫調控和營養性免疫調控。自身免疫調控是先天獲得的,依賴遺傳因素的好壞。篩選優良個體,可從根本上解決免疫力的問題,但過程極其繁瑣、復雜。營養性免疫調控依賴營養物質提高蛋雞的免疫能力,這個過程在實際工作中比較好操作,已經有大量的研究成果。
2.1 自身免疫調控
家禽品系與抗病力密切相關[57]。通過基因修飾和篩選,保留含有優良生長性狀和抗病基因的雞種,是免疫調控策略的根本。機體中相關基因直接調控免疫。如野生型p53誘導的磷酸酶1基因(wild-type p53-induced phosphatasel,Wip1)正向調控T細胞、B細胞的發育及功能發揮。Sun等研究證實,Wip1基因限制p38 MAPK通路正向調控髓質胸腺上皮細胞(medullary thymic epithelial cell,mTEC)成熟、內穩態和再生,進而促進T細胞增殖分化,提升免疫功能[58]。Yi等研究表明,在p53依賴性凋亡能力增強的條件下,Wip1基因敲除小鼠會出現B細胞前期發育受損,導致骨髓、外周血、脾臟等B細胞數量減少,直接影響B細胞功能發揮[59]。MHC抗病基因與機體易感性和抗性相關,是典型的抗性遺傳基因,影響先天免疫[60]。IFN-γ可激活巨噬細胞,增強MHC-Ⅰ、MHC-Ⅱ分子能力,阻斷病毒復制,清除細胞內病原體[61]。IL-2基因可促進脾臟T細胞增殖, 增加NK細胞活性;IL-7可誘導B細胞、T細胞活化[62]。
細胞中部分miRNA可以通過靶向免疫相關分子或病毒基因影響病毒復制。研究表明,干擾素調節因子1(interferon regulatory factor 1,IRF1)是miR-23b 的一個潛在靶點,兩者具有相反表達模式,即miR-23b過表達與IRF1敲除均能增強 ALV-J 的復制,表明miR-23b可能通過靶向IRF1在ALV-J復制中發揮重要作用[63];同時也有研究顯示,細胞miRNA靶向IBDV基因組中vp1和vp2基因構成重組病毒載體,轉染到DF-1細胞中,抑制IBDV復制[64]。許多miRNAs參與宿主-病原體的相互作用,調節免疫基因和通路的表達。研究發現,新城疫病毒疫苗株感染雞巨噬細胞microRNA表達譜中差異基因多富集在RIG-I樣受體信號通路、NOD樣受體信號通路、絲裂原活化蛋白激酶(MAPK)信號通路等與免疫相關的信號通路中,調控免疫反應[65];類似研究表明,經過GO富集分析(gene ontology)后,與雞流感病毒感染相關的miR-34c、miR-1b、miR-1a等差異基因在AIV H5N3感染后上調。miR-206等在甲型流感病毒感染后下調,與免疫功能調節直接相關[66];另有研究證實,細胞miRNA中差異基因主要參與免疫相關細胞因子-細胞因子受體相互作用的調節。如miR-34a上調,促進INF-γ產生的輔助性T淋巴細胞1(T helper lymphocyte 1,Th1)分化,促進IL-6、IL-17表達,調控免疫[67]。細胞miRNA可以成為未來自身免疫調控的研究重點。
細胞中lncRNAs調控干擾素刺激基因(interferon-stimulated genes,ISGs)表達,干擾和抑制病毒感染,限制病毒復制,參與先天性免疫應答[68-69];在CD4+T細胞向TH1或TH2亞群分化狀態下,STAT4、T-box等TH1相關轉錄因子誘導部分TH1特異性lncRNAs的表達,STAT6調節TH2特異性lncRNAs的表達。lncRNA在調節適應性免疫中發揮著關鍵作用[69]。
sncRNA在宿主-病毒相互作用中鮮有報道。miRNA、lncRNA以及兩者之間的相互作用可作為免疫方面新的前沿研究領域。
各種免疫球蛋白直接特異性識別、結合抗原,產生免疫應答。MHC編碼生成的主要組織相容性抗原介導特異性免疫應答;ISGs編碼生成包括IFN糖蛋白在內的一系列蛋白,抑制病毒DNA復制,降低活性,刺激特異性免疫[70]。
綜上所述,機體在DNA、RNA、蛋白水平構成關系互作網,相互作用,影響機體自身免疫調控功能的正常發揮。
2.2 營養性免疫調控
利用各種營養調控機制,在特定營養物質需要期,結合不同類營養物質成分的獨特優點,與機體各個系統之間相互作用,維持自身正常生理動態平衡與穩定生理功能的調控,即營養性免疫調控。
2.2.1 營養調控機制
一定生理狀態下,營養物質轉運體介導營養物質跨過血漿膜被吸收利用,不同類型和數量的細胞因子通過選擇性作用于白細胞上不同的細胞因子受體來提升對營養物質的吸收利用率[71]。營養物質主要通過以下幾種調控機制對機體免疫能力進行調控。
免疫反應前:一些營養物質在宿主體內重新分配,作為限制性營養底物抵抗病原微生物。如生物素是雞體內病菌生長的第一限制性營養底物,鐵僅次于生物素[72]。發生急性感染時,鐵離子和鋅離子離開血液循環,蓄積于肝臟和肝外組織中,以此來限制病原微生物的利用。雞體內受到刺激的巨噬細胞也可分泌抗生物素蛋白,使感染部位的病原體發生生物素饑餓,抵制病原入侵[73-74];營養物質為免疫系統提供底物,滿足其代謝需求,促進免疫器官、免疫細胞的形成與發育,調節胚胎時期就進行的影響機體免疫功能正常發揮的發育活動[75]。如維生素A、維生素D、維生素C、維生素E、維生素B6、維生素B12、葉酸以及微量元素鋅、鐵、銅、硒等已經被證實對免疫系統的完整性起重要作用[76],通過免疫營養互作效應來調控免疫[77]。營養不足時,機體對疾病的抵抗能力嚴重下降。
免疫反應中:首先,營養可減少免疫反應中的細胞損傷。免疫反應過程中,激活免疫細胞的同時,釋放具有破壞作用的分子活性氧(reactive oxygen species,ROS)。其中的超氧化物通過酶的轉化引起廣泛的細胞損傷;自由基、過氧化物會對細胞造成永久性損傷甚至導致動物死亡[78]。而外源性抗氧化劑維生素C、維生素E與內源性抗氧化防御系統協同,保護細胞膜免受正常代謝過程中產生的自由基,以及接觸毒素和污染物而造成的損害[79]。而且使用動物模型的早期研究已經建立了維生素E缺乏與免疫功能損傷之間的明確關系。雞的淋巴細胞增殖抑制可通過補充維生素E來逆轉[80]。因此常使用維生素C、維生素E、谷胱甘肽[81]等營養類抗氧化劑保護細胞免受ROS的傷害。其次,影響腸道動力。Ponsuksili等研究發現,蛋雞腸道miRNAs(miR-143-3p、miR-21、miR-16等)參與JAK-STAT信號通路和細胞信號轉導,調節先天免疫和適應性免疫。miR-126-3p、miR-214、miR-24-3p、miR-726-5p和miR-29b-3p等,介導能量代謝和線粒體功能的分子途徑,滿足腸道對蛋雞在生產期間不斷變化的需求[53]。由此發現,miRNAs在營養物質刺激下調節免疫的同時,維持腸道穩態。同時也有研究顯示,蛋雞腸道微生物菌群,寄生于胃腸道黏膜表面,彼此競爭營養物質,通過短鏈脂肪酸(short chain fatty acids,SCFAs)產生調節性T細胞,IFN-α、IFN-β、IL-22等抗炎細胞因子和IgA、IgG等免疫球蛋白,結合腸道-肺、腸道-腦、腸道-皮膚、腸道-肝等調節渠道,利用不同免疫機制進行免疫調節的同時,維持腸道穩態環境,影響腸道動力[56,82]。除此之外,營養調節白細胞內信號傳導及特定因子表達。維生素E調控白細胞活素的釋放量,通過減少白細胞與抗體接觸,降低蛋雞H5-AI、H9-AI等抗體滴度,提高前列腺素E2(prostaglandin E2,PGE2)、IL-1、TNF-α等含量,促進特定細胞因子、炎性分子信號轉導和表達調控免疫[83]。最后,日糧調控體內激素環境水平,影響免疫。洋蔥、大蒜提取物能產生類似胰島素的影響,刺激體內胰島素、生長激素、胰高血糖素的分泌,起代謝調節作用[84];飼料中基本營養素蛋氨酸、精氨酸、賴氨酸、異亮氨酸缺乏促使雞血漿三碘甲狀腺原氨酸(triiodothy-ronine,T3)升高,蛋氨酸缺乏降低胰島素樣生長因子2(insulin like growth factor2,IGF-2)水平[85]。由于白細胞血漿膜表面有不同的激素受體,如胰島素、胰島素樣生長因子、胰高血糖素等,營養變化造成的激素變化必然會影響到白細胞對營養物質的吸收和利用,影響免疫效率。提示營養物質對免疫可能起至關重要的作用。
有效利用各種營養調控機制是發揮營養物質調控免疫的基礎,確定各特殊階段的營養需要量是充分發揮營養物質調控免疫的關鍵。
2.2.2 營養需要期
免疫系統是一個擴散的、遍布全身的、動態的系統,隨時都在發生各種信號傳遞、代謝過程、生化反應,影響機體對營養物質的利用,使營養需要量發生改變,影響代謝平衡。目前很難通過經典的試驗測定方法檢測出發生免疫反應時的營養需要量,只能根據免疫特點確定各自重要的營養需要期[71]。(1)先天性免疫系統的重要營養需要期:感染初期,先天性免疫系統被激活,對各類病原菌起全身性炎性反應,刺激分泌急性期蛋白(acute phase protein,APP)和NO等物質影響代謝;炎性反應后,蛋雞處于代償性生長階段,由于快速生長的需要,營養需要量快速增加[86];先天性免疫系統對抗原沒有記憶效應,當再次接觸到相同抗原時,激活這個系統所需要的營養需要量與初次接觸抗原時的需要量是相似的。(2)適應性免疫系統的重要營養需要期:初級免疫反應中淋巴細胞發育對適應性免疫系統功能發揮起重要作用。即淋巴細胞對應的發育期為其重要的營養物質需要期。B細胞、T細胞的不同發育特點導致胚胎期第5、6天至出生后3周內是T細胞的重要營養物質需要期,胚胎期第15天到出生后6周內是B細胞的重要營養物質需要期;淋巴細胞激活即淋巴細胞初次接觸特異性抗原,以及記憶性淋巴細胞再次接觸相同抗原[71]。需大量營養物質滿足其分化增殖、產生抗體等,屬于重要的營養物質需要期。
在免疫反應的特殊階段,需要額外的營養供給。如何使免疫能效最大化,則需參照各種營養成分的免疫特殊性,進行綜合考慮,確定最佳的配比。
2.2.3 不同類營養物質對蛋雞的免疫調控
各種營養成分有獨特的免疫作用,不同類營養物質相互協作,共同參與蛋雞的免疫調控。
2.2.3.1 飼料營養水平
能量影響免疫器官發育、激素分泌、免疫功能正常發揮。攝入不足或過量都會造成免疫器官萎縮、腺體激素分泌不足、內分泌功能異常、免疫功能紊亂、抗體能力下降[87]。但適當能量限飼又可以在一定程度上提高肉仔雞脾臟免疫器官指數[88]。因此未來研究可以多關注適當能量與最佳免疫效率之間的關系。
蛋白質和氨基酸促進體內免疫系統發育、酶和抗體生成。蛋氨酸為家禽第一限制性氨基酸,若缺乏,直接限制了半胱氨酸形成谷胱甘肽(glutathione,GSH)的效率,導致淋巴細胞數量及增殖指數下降,甚至影響法氏囊發育,降低IL-2含量[89]。GSH是免疫應激時的必需氨基酸,促進樹突狀細胞、T淋巴細胞以及IL-12增殖,刺激嗜中性粒細胞、NK細胞活性,抑制細胞產生ROS,保護免疫細胞不受損傷[90]。谷氨酰胺(glutamine,Gln)是各種免疫細胞的重要能源物質。促進淋巴細胞增殖,刺激產生 IL-2,增加腸黏膜分泌型IgA(secretory IgA,sIgA)分泌,提升免疫細胞活性,增強新城疫HI抗體滴度水平[91]。精氨酸誘導IGF-I、免疫調節因子NO,刺激淋巴細胞、IL-2分泌,參與受損組織的修復,刺激巨噬細胞發揮吞噬功能,提高免疫反應效率[92]。蘇氨酸是IgG的主要成分,刺激免疫球蛋白、淋巴細胞、抗體生成,提高特異性免疫功能[93];研究表明,添加蘇氨酸的量由0.47%增長到0.77%,42周齡信陽綠殼蛋雞體內IgG(1.24 g/L)、IgM(0.047 g/L)、IgA(0.416 g/L)顯著增長23.39%、65.96%、16.83%[94];類似研究顯示,日糧添加0.7%蘇氨酸可直接提高雛雞ND抗體效價[95]。因此蘇氨酸可能直接影響體液免疫應答水平。
維生素在一定程度上都能改善免疫效果。維生素A促進淋巴細胞分化生成T細胞和B細胞,增強巨噬細胞的吞噬作用,缺乏時嚴重影響免疫[96];維生素D尤其是維生素D3,調控T細胞分化,誘導B細胞增殖分化,介導特異性免疫反應。在體內轉化成25-OH-D3和1,25-(OH)2-D3啟動抗菌反應[97]。10-9~10-7 mol/L濃度的1,25(OH)2D3協同2 mmol/L濃度丁酸刺激雞HD11巨噬細胞系可使禽β-防御素(avian β-defensin,AvBD)尤其是AvBD9基因表達量提升4 900倍,從而提升免疫能力[98];維生素E促進免疫器官發育、T淋巴細胞生成,作為免疫佐劑與硒協同影響免疫功能,抑制干擾免疫反應的前列腺素生成[99]。另有研究表明,52周齡的天府肉種雞,在槲皮素+維生素E混合組合刺激下,INF-γ、IL-2基因在脾臟中的表達量由1.33、0.56顯著提升到5.79和1.50,直接促進 INF-γ、IL-2表達,調控免疫反應[100]。維生素C可緩解熱應激對蛋雞免疫功能的抑制,促進免疫器官發育、淋巴細胞產生,維持吞噬細胞的吞噬能力,促使免疫蛋白中血清總蛋白(total protein,TP)、γ球蛋白等、補體(CD3、CD4等)含量增加,提高蛋雞NDV抗體滴度,提高抗體效價,提升免疫力[101-102];VB9可顯著提高IgA、IgM、sIgA、IL-10等血清免疫指標,改善腸道屏障功能[103]。提示無論是脂溶性還是水溶性維生素可能對蛋雞免疫都起關鍵性作用。
礦物質作為輔助因子,優化激素功能,調節免疫。適量鋅可產生促炎細胞因子(IL-1β、IL-6和TNF-α),調節巨噬細胞、嗜中性粒細胞吞噬功能,維持NK細胞活性,促進T細胞正常發育、功能正常發揮,促進B細胞抗體生成、避免程序性死亡[104]。錳促進上皮細胞發育、修復、維持,促進生長,改善蛋殼品質,降低蜂窩織炎,加強對ND等傳染性疾病的抵抗力。硒抗氧化,促進抗體生成,增強T細胞反應。鐵促進T細胞增殖,調控淋巴細胞產生細胞因子。鈣是蛋殼的主要組成成分,但同時也增強免疫效果。在促鈣化激素VD3、甲狀旁腺素(parathyroid hormone,PTH)、降鈣素(calcitonin,CT)等刺激下會促進腸道鈣吸收,增加IgG、IgA等的含量[97,105]。高氟飼糧抑制黏蛋白2(mucin 2,MUC2)、sIgA生成,降低黏膜免疫系統功能,影響炎癥反應,降低抵抗力[106]。將氨基酸與微量元素螯合形成氨基酸螯合物,具備更加優良的化學穩定性,更高的生物學效價,可被蛋禽更高效利用。如羥基蛋氨酸鋅(hydroxymethionine chelated zinc,MHA-Zn)可顯著降低脾臟促炎因子IL-8 mRNA的表達,提高蛋雞生產性能,蛋品質和免疫機能[107];40mg/kg蛋氨酸錳可顯著提高IgA、IgG,補體分子C4的含量[108]。將微量元素與小肽螯合,形成微量元素小肽螯合物,如小肽螯合鐵、銅、錳、鋅和納米硒能顯著提高IgM、IgG、TP、白蛋白(albumin,ALB)等免疫指標的含量,增強免疫效果[109]。未來可進一步探索礦物質螯合物對免疫的作用機理。
2.2.3.2 飼料添加劑
植物提取物、微生態制劑等飼料添加劑,添加少量但作用顯著,節省飼料成本的同時,可提高家禽生產性能、免疫效果、保證動物體健康,改善蛋品質。
研究表明,日糧中添加300 mg/kg復合植物提取物,能顯著提高蛋雞的產蛋水平,增加IgM、IgG、超氧化物歧化酶(superoxide dismutase,SOD)、谷胱甘肽過氧化物酶(glutathione peroxidase,GSH-Px)含量,改善機體免疫功能及抗氧化能力[110]。0.5%的生姜粉和100 mg/kg的姜油可顯著提高肉仔雞胸腺、法氏囊指數、ND抗體效價,增加淋巴細胞、粒細胞數量[111]。0.03%的紫蘇籽提取物,可顯著提高蛋雞脾臟指數,提高IgA、IgG、IgM、IL-2含量[112]。0.4 g/kg的大蒜素,可促進免疫器官的發育,提高IgA、IgG、IgM含量,尤其提高AIV病毒、NDV的抗體滴度,促進體液免疫的發揮[36]。添加一定量大青葉正丁醇提取物可抑制LPS降低中性粒細胞CXC趨化因子受體(CXC-chemokine receptor,CXCR)CXCR1、CXCR2、L-選擇素(CD62L)和IL-8的表達水平,激活髓過氧化物酶,提升中性粒細胞的遷移能力[113]。一定量的甜菜堿,能提高營養物質消化、吸收、利用率的同時,增加淋巴細胞的數量,增加免疫器官質量[114]。8%的金針菇莖廢料或金針菇菌渣等可促進sIgA、IgG、IL-2、IL-4、IL-6、TNF-α 分泌,提升相關免疫抑制性疾病的抗體滴度,代替抗生素類藥物提前調控蛋雞的免疫應答和血清免疫,提高免疫調節和抗氧化活性[37]。5%的苜蓿草粉促進蛋黃著色的同時,可降低ND、AI抗體水平,提高蛋雞免疫力[115]。200 mg/kg 萬壽菊提取物,其中有效成分葉黃素提升太行雞蛋黃顏色同時,可增加IgA、IgM、IgG含量,增強免疫能力[116]。基礎日糧中添加400 mg/kg的竹葉黃酮飼喂1日齡的愛拔益加肉雞14~42 d時,INF-γ、IL-2基因在脾臟中的表達量由1.90、0.41,提升35.26%、48.78%,直接提高血清細胞因子INF-γ、IL-2水平,增強其免疫調節功能[117]。一定量的復方中草藥添加劑可促進雛雞免疫器官、免疫細胞生長發育,刺激機體免疫應答,誘導免疫細胞分泌免疫球蛋白,延長抗體高峰期[118-119]。由此發現,添加一定量的植物提取物對蛋雞免疫性能的提升起著至關重要的作用。
飼料中添加微生態制劑,提升蛋雞生產性能的同時,促進有益菌成為優勢菌群,調節腸道平衡,促進免疫器官生長發育,免疫細胞增殖,增強機體免疫能力,降低死淘率,延緩抗體滴度下降,提高抗體效價水平[120]。研究表明,添加一定量的益生菌微生態制劑,可激活吞噬細胞活性。促進腸道上皮內淋巴細胞產生細胞因子,提升IgA水平,增強免疫應答能力[121];類似研究發現,添加1%的復合微生物制劑,能顯著提高IgG、IgA含量,增強蛋雞免疫功能[122];除此之外,日糧中添加100 g有效活菌為1×1010 CFU/g的屎腸球菌會比添加同等量枯草芽孢桿菌、凝結芽孢桿菌等更能有效提升京紅1號蛋雛雞ND、H5-Re4等抗體值,提高抗體水平[123]。
蛋雞飼料中添加一定量低聚木糖,可顯著提高血漿IgA、IgM、TNF-α、IL-2的濃度[124]。添加 3 ku 的殼寡糖,可增加蛋雞脾臟TNF-α、IL-2的基因表達量,增強免疫力[125]。添加0.2%的復合酸化劑,可顯著增加海蘭褐蛋雛雞生長后期的IgG含量,促進免疫性能的提高[126]。添加0.1 g/kg的抗菌肽可顯著提高IgG、T-AOC含量,降低血尿素氮(blood urea nitrogen,BUN)含量,調節免疫功能,提高抗氧化能力[127]。部分研究顯示,添加1.0 mg/kg富硒蚯蚓粉(selenium enriched earthworm powder,SEP)可顯著提高血清中溶菌酶的吞噬活性,提高蛋雞IgG、IL-2含量,正向調節 IFN-γ 生成[128];類似研究表明,聯合添加0.3%的蟲草粉和0.1 mg/kg的酵母硒,可顯著提高蛋雞產蛋高峰時期IL-2、IFN-γ水平,總抗氧化能力(total antioxidant capacity,T-AOC)、總超氧化物歧化酶(total superoxide dismutase,T-SOD)活性,同時顯著降低血清中的丙二醛(malondialdehyde,MDA)含量,這些指標與免疫功能高低顯著相關[129]。由此可見,未來關于飼料添加劑的研究應更加關注物質間的配伍作用,使免疫效果最大化。
自身免疫調控是根本,從基因角度增強免疫能力;而營養免疫調控是關鍵,從日常飲食中增強免疫。營養免疫調控更省時省力,能通過多種調節機制調節免疫系統,有效提高蛋雞免疫力,是最綠色、健康、便捷的方式。
3 結論及展望
免疫是影響蛋雞產業持續健康穩步發展的重要因素。了解免疫抑制性疾病的特點、掌握蛋雞免疫基礎知識、運用恰當的調控策略,能使機體高效發育的同時,保持完善的免疫機能,有效解決機體藥物耐受及殘留問題。目前研究多集中在營養免疫調控上,研究成果也能解決部分商品蛋雞免疫力下降的問題,在一定程度上維持蛋雞產業可持續發展。但未來仍需注意以下問題:(1)如何通過有效搭配使每種營養素調控免疫的效率最佳,從而解決蛋雞不同生長階段因生理特征不同而感染不同疾病的問題。(2)達到優良免疫性能下的營養需要還沒有成為國際飼養標準中營養需要的衡量標準。至于上述存在的問題,隨著研究的不斷深入,終將能夠實現不同階段不同狀態針對性處理,滿足最佳免疫力的同時,有最高的生產性能和效益,制定出更加全面的飼養標準。而這無疑不再一次推動免疫營養學的發展,使中國蛋雞養殖業又更進一步。
參考文獻:
[1]馬 驥. 中國蛋雞養殖業的發展及對策[J]. 北方牧業,2019(3):11-12.
[2]孫從佼,于愛芝,佘惠靈,等. 2021年蛋雞產業發展情況、未來發展趨勢及建議[J]. 中國畜牧雜志,2022,58(3):210-215.
[3]彭夢華,謝曉鵬 .動物的營養與免疫[J]. 江西飼料,2014(1):1-3,7.
[4]張春艷. 動物疫病與免疫系統及營養的關系[J]. 養殖技術顧問,2012(1):238.
[5]左麗霞,楊 景. 新形勢下蛋雞產業高質量發展關鍵點[J]. 今日畜牧獸醫,2021,37(4):42-43.
[6]陳淑增,楊 翀,邱丹纓. 病原生物學與免疫學[M]. 2版.武漢:華中科技大學出版社,2015.
[7]羅天海,Yegani M,Butcher G,等. 免疫抑制正威脅著家禽的健康和福利[J]. 國外畜牧學(豬與禽),2005,25(4):51-55.
[8]Madej J P,Stefaniak T,Bednarczyk M.Effect of in ovo-delivered prebiotics and synbiotics on lymphoid-organs morphology in chickens[J]. Poultry Science,2015,94(6):1209-1219.
[9]Chen J,Zhu X Q,Yang L,et al. Effect of Glycyrrhiza uralensis Fisch polysaccharide on growth performance and immunologic function in mice in Ural City,Xinjiang[J]. Asian Pacific Journal of Tropical Medicine,2016,9(11):1078-1083.
[10]楊樹寶,張桂山,徐 晶,等. 不同日齡雞喉黏膜組織中T淋巴細胞亞群的分布特點[J]. 動物醫學進展,2021,42(11):19-23.
[11]Nace G,Evankovich J,Eid R,et al. Dendritic cells and damage-associated molecular patterns:endogenous danger signals linking innate and adaptive immunity[J]. Journal of Innate Immunity,2011,4(1):6-15.
[12]伍芳芳. 猴頭菇多糖的結構表征、免疫調節活性及其機理研究[D]. 廣州:華南理工大學,2018.
[13]王麗娜,吳 波,丁國棟,等. 金針菇菇腳對肉雞生產性能及免疫功能的影響[J]. 菌物研究,2013,11(2):120-123.
[14]張 萍,劉海俠,江善祥.赤靈芝多糖對雞淋巴細胞增殖的影響[J]. 中國家禽,2015,37(20):54-56.
[15]馬成才,周 英,許天齊,等. 免疫治療6周后中性粒細胞淋巴細胞比率及乳酸脫氫酶對非小細胞肺癌預后預測價值[J]. 臨床軍醫雜志,2022,50(3):246-250.
[16]方磊涵,王 振,王 留,等. 中藥復方多糖對肉仔雞生長性能和免疫功能的影響[J]. 中國獸醫雜志,2018,54(9):56-59.
[17]孫甜甜. 枸杞粗多糖對肉仔雞生長性能、抗氧化及免疫功能的影響[D]. 長春:吉林農業大學,2019.
[18]Espinosa V,Rivera A. Cytokines and the regulation of fungus-specific CD4 T cell differentiation[J]. Cytokine,2012,58(1):100-106.
[19]Chu W M.Tumor necrosis factor[J]. Cancer Letters,2013,328(2):222-225.
[20]黃俊祥. 氟中毒對肉仔雞血細胞數、血清溶菌酶和補體C3、C4含量的影響[J]. 現代牧業,2020,4(2):30-33.
[21]He Y N,Xie Z W,Dai J L,et al. Responses of the Toll-like receptor and melanoma differentiation-associated protein 5 signaling pathways to avian infectious bronchitis virus infection in chicks[J]. Virologica Sinica,2016,31(1):57-68.
[22]Bhattacharya A,Eissa N T.Autophagy as a stress response pathway in the immune system[J]. International Reviews of Immunology,2015,34(5):382-402.
[23]孫秋艷,韓偉濤,王彩霞. 靈芝多糖對雞新城疫疫苗抗體滴度的影響[J]. 山東畜牧獸醫,2012,33(5):18-19.
[24]方磊涵,王 振,劉詩柱,等. 中草藥復方多糖對肉雞腸道菌群及免疫功能的影響[J]. 中國飼料,2019(9):71-75.
[25]張彤瑤,高雅松,張愛龍,等. 金針菇菇腳對肉雞T、B淋巴細胞免疫功能的影響[J]. 中國農業大學學報,2016,21(4):86-94.
[26]尹艷軍,郝延剛. 凍干乳酸菌對肉雞生長性能和免疫功能的影響[J]. 飼料工業,2015,36(10):36-40.
[27]Takeuchi A,Badr M E,Miyauchi K,et al. CRTAM determines the CD+4cytotoxic T lymphocyte lineage[J]. The Journal of Experimental Medicine,2016,213(1):123-138.
[28]Husnain M,Mackrides N,Vega F,et al. CD+4/CD+8 immunophenotype switching as a marker for intraocular and CNS involvement in mycosis fungoides[J]. Leukemia & Lymphoma,2019,60(5):1308-1311.
[29]Wang K L,Cui P F,Ni R Q,et al. Chicken-derived pattern recognition receptor chLGP2 inhibits the replication and proliferation of infectious bronchitis virus[J]. Frontiers in Microbiology,2022,12:810215.
[30]林曉艷. 不同飼養環境對蛋雞健康的影響[D]. 泰安:山東農業大學,2018.
[31]張英楠,徐 晶,張桂山,等. 刺五加多糖對雛雞脾臟中CD+4和CD+8T淋巴細胞定位分布的影響[J]. 中國獸藥雜志,2021,55(9):44-49.
[32]閆先峰,唐偉麗. 女貞子對蛋雞生產性能·血清生化和免疫性能的影響[J]. 安徽農業科學,2021,49(18):96-98.
[33]呂新彪. 二至丸對產蛋后期蛋雞免疫和抗氧化功能及生產性能的影響[D]. 邯鄲:河北工程大學,2021.
[34]Idowu P A,Idowu A P,Zishiri O T,et al. Activity of mannose-binding lectin on bacterial-infected chickens—a review[J]. Animals,2021,11(3):787.
[35]辛小青. 飼糧中添加苜蓿粗多糖對蛋雞生產性能、蛋品質及免疫指標的影響[D]. 北京:中國農業科學院,2016.
[36]尹 典,葉美怡,李敬雙,等. 大蒜素對蛋雞免疫功能、蛋品質和生產性能的影響[J]. 飼料研究,2021,44(13):61-63.
[37]陳盛絮,李 冰,潘秋明,等. 金針菇菌渣對蛋雞生產性能、鈣代謝、免疫性能和抗體滴度的影響[J]. 中國飼料,2018(14):88-92.
[38]Kogut M H,Lee A,Santin E. Microbiome and pathogen interaction with the immune system[J]. Poultry Science,2020,99(4):1906-1913.
[39]He C F,Wang S,Zhou C K,et al. CD36 and LC3B initiated autophagy in B cells regulates the humoral immune response[J]. Autophagy,2021,17(11):3577-3591.
[40]馬 暢,孟凡生. 家禽免疫抑制疾病致病因子的研究進展[J]. 家禽科學,2015(11):51-54.
[41]黎 敏,徐丘璠,陳禮斌,等. 雞幾種重要免疫抑制性疾病的流行現狀和防控[J]. 養禽與禽病防治,2021(10):25-32.
[42]卞 璐. 雞常見細菌性疾病的特點及防治分析[J]. 中國畜禽種業,2021,17(7):190-191.
[43]趙桂春. 雞腸道寄生蟲感染情況調查[J]. 畜牧獸醫科學(電子版),2021(1):36-37.
[44]陳 琳. 雞常見細菌性疾病分類及預防治療[J]. 吉林畜牧獸醫,2022,43(1):70,72.
[45]蔡 莉,王 芹,李 方,等. 飼養管理不當引起家禽免疫抑制的原因[J]. 養殖技術顧問,2012(10):41.
[46]沈 霞,霍清合. 引起家禽免疫抑制的因素及防治對策[J]. 家禽科學,2014(1):45-46.
[47]谷風柱. 構建家禽堅強免疫體系的解決方案[J]. 中國家禽,2011,33(1):33-34.
[48]唐彩琰,邵建忠,Dvm R B. 免疫系統受抑制是否影響家禽的生長[J]. 國外畜牧學(豬與禽),2017,37(11):59-60.
[49]張浩輝. 淺析飼料營養對動物免疫機能的影響[J]. 今日畜牧獸醫,2018,34(7):60.
[50]張 建,馬 飛. 雞霉菌毒素中毒的防治措施[J]. 獸醫導刊,2021(3):111-112.
[51]Verwoolde M B,van den Biggelaar R H G A,de Vries Reilingh G,et al. Innate immune training and metabolic reprogramming in primary monocytes of broiler and laying hens[J]. Developmental and Comparative Immunology,2021,114:103811.
[52]常金萍. 蛋雞應激產生的原因、危害及預防[J]. 現代畜牧科技,2019(8):106-107.
[53]Ponsuksili S,Hadlich F,Reyer H,et al. Genetic background and production periods shape the microRNA profiles of the gut in laying hens[J]. Genomics,2021,113(4):1790-1801.
[54]薛玉霞. 家禽飲水免疫十要點[J]. 鄉村科技,2014(3):30.
[55]關 蘊,焦培榮,劉 紅. 家禽免疫失敗的原因分析及防治措施[J]. 養禽與禽病防治,2019(12):16-19.
[56]Chen C J,Wu G H,Kuo R L,et al. Role of the intestinal microbiota in the immunomodulation of influenza virus infection[J]. Microbes and Infection,2017,19(12):570-579.
[57]Cheng H H,Kaiser P,Lamont S J.Integrated genomic approaches to enhance genetic resistance in chickens[J]. Annual Review of Animal Biosciences,2013,1:239-260.
[58]Sun L N,Li H R,Luo H Y,et al. Phosphatase Wip1 is essential for the maturation and homeostasis of medullary thymic epithelial cells in mice[J]. Journal of Immunology,2013,191(6):3210-3220.
[59]Yi W W,Hu X L,Chen Z Y,et al. Phosphatase Wip1 controls antigen-independent B-cell development in a p53-dependent manner[J]. Blood,2015,126(5):620-628.
[60]Kaufman J. Innate immune genes of the chicken MHC and related regions[J]. Immunogenetics,2022,74(1):167-177.
[61]Yang X,Arslan M,Liu X J,et al. IFN-γ establishes interferon-stimulated gene-mediated antiviral state against Newcastle disease virus in chicken fibroblasts[J]. Acta Biochimica et Biophysica Sinica,2020,52(3):268-280.
[62]Huo S S,Zhang J L,Fan J H,et al. Co-expression of chicken IL-2 and IL-7 enhances the immunogenicity and protective efficacy of a VP2-expressing DNA vaccine against IBDV in chickens[J]. Viruses,2019,11(5):476.
[63]Li Z H,Chen B,Feng M,et al. MicroRNA-23b promotes avian leukosis virus subgroup J (ALV-J) replication by targeting IRF1[J]. Scientific Reports,2015,5:10294.
[64]Zhou L Y,Zheng S J. The roles of microRNAs (miRNAs) in avian response to viral infection and pathogenesis of avian immunosuppressive diseases[J]. International Journal of Molecular Sciences,2019,20(21):5454.
[65]Mu J Q,Liu X X,Yu X B,et al. Cellular microRNA expression profile of chicken macrophages infected with Newcastle disease virus vaccine strain LaSota[J]. Pathogens,2019,8(3):123.
[66]Kemp V,Laconi A,Cocciolo G,et al. miRNA repertoire and host immune factor regulation upon avian coronavirus infection in eggs[J]. Archives of Virology,2020,165(4):835-843.
[67]Li T,Song Y P,Bao X Y,et al. The mediation of miR-34a/miR-449c for immune cytokines in acute cold/heat-stressed broiler chicken[J]. Animals,2020,10(11):2168.
[68]Poynter S J,DeWitte-Orr S J.Fish interferon-stimulated genes:the antiviral effectors[J]. Developmental and Comparative Immunology,2016,65:218-225.
[69]Meng X Y,Luo Y Z,Anwar M N,et al. Long non-coding RNAs:emerging and versatile regulators in host-virus interactions[J]. Frontiers in Immunology,2017,8:1663.
[70]Schoggins J W.Interferon-stimulated genes:what do they all do?[J]. Annual Review of Virology,2019,6(1):567-584.
[71]蘇記良,楊 琳,佟建明. 家禽免疫系統的營養需要[J]. 中國家禽,2007,29(7):48-50,53.
[72]Klasing K C,喬 娜(譯). 雞的營養和免疫系統[J]. 中國家禽,2008,30(20):25-29.
[73]潘家強,葉承榮,王小龍. 營養調控對家禽免疫功能的影響[J]. 畜牧與獸醫,2005,37(7):58-60.
[74]王 斌. 家禽的營養與免疫[J]. 動物科學與動物醫學,2003,20(9):60-62.
[75]Humphrey B D. 家禽免疫系統營養需求的研究現狀與趨勢[J]. 林又米,譯. 中國家禽,2011,33(5):4-8.
[76]Gombart A F,Pierre A,Maggini S. A review of micronutrients and the immune system-working in harmony to reduce the risk of infection[J]. Nutrients,2020,12(1):236.
[77]楊小軍,姚軍虎,咼于明. 畜禽免疫營養研究進展及其互作評價[J]. 飼料工業,2011,32(2):61-64.
[78]Isnaini I,Permatasari N,Mintaroem K,et al. Oxidants-antioxidants profile in the breast cancer cell line MCF-7[J]. Asian Pacific Journal of Cancer Prevention,2018,19(11):3175-3178.
[79]Maggini S,Beveridge S,Sorbara P J P,et al. Feeding the immune system:the role of micronutrients in restoring resistance to infections[J]. CAB Reviews:Perspectives in Agriculture,Veterinary Science,Nutrition and Natural Resources,2008,3:98.
[80]Wu D Y,Lewis E D,Pae M,et al. Nutritional modulation of immune function:analysis of evidence,mechanisms,and clinical relevance[J]. Frontiers in Immunology,2019,9:3160.
[81]劉世龍,朱曉萍,秦江帆,等. 谷胱甘肽對畜禽生長性能和機體健康的影響[J]. 廣東畜牧獸醫科技,2020,45(2):24-26.
[82]Abaidullah M,Peng S W,Kamran M,et al. Current findings on gut microbiota mediated immune modulation against viral diseases in chicken[J]. Viruses,2019,11(8):681.
[83]Zhang X H,Zhong X,Zhou Y M,et al. Dietary RRR-α-tocopherol succinate attenuates lipopolysaccharide-induced inflammatory cytokines secretion in broiler chicks[J]. The British Journal of Nutrition,2010,104(12):1796-1805.
[84]Rabinowitch H D,Currah L. Allium crop science:recent advances[M]. UK:CABI Publishing,2002.
[85]Carew L B,McMurtry J P,Alster F A. Effects of methionine deficiencies on plasma levels of thyroid hormones,insulin-like growth factors-Ⅰ and-Ⅱ,liver and body weights,and feed intake in growing chickens[J]. Poultry Science,2003,82(12):1932-1938.
[86]焦玉萍,徐向陽,崔海軍. 雞日糧營養水平與免疫機能[J]. 河南畜牧獸醫,2002,23(6):8-10.
[87]官麗輝,劉海斌,張立永,等. 不同能量水平日糧對塞北烏骨雞消化器官發育、肌肉脂肪酸相對含量及免疫器官指數的影響[J]. 河南農業科學,2017,46(7):106-109.
[88]劉路路. 能量限制對三黃雞生長性能、免疫特性及脾臟基因表達的影響[D]. 鄭州:河南農業大學,2016.
[89]劉怡琳. 低蛋氨酸對產蛋后期蛋雞生產性能和抗氧化能力的影響[D]. 洛陽:河南科技大學,2017.
[90]Rodrigues C,Percival S S. Immunomodulatory effects of glutathione,garlic derivatives,and hydrogen sulfide[J]. Nutrients,2019,11(2):295.
[91]李秀群. 氨基酸對動物營養免疫的影響[J]. 現代畜牧科技,2015(1):28-29.
[92]葉 婷,田科雄. 精氨酸對機體免疫系統的影響及相關機理[J]. 飼料博覽,2010(7):11-14.
[93]畢 曄,潘曉花,辛海瑞,等. 蘇氨酸在家禽營養中的研究進展[J]. 中國畜牧獸醫,2017,44(8):2326-2332.
[94]Li J M,Zhang X Y,Yuan C,et al. Effects of dietary L-threonine levels on antioxidant capacity,digestive enzyme activities,and antibody production of Xinyang green-shell laying hens[J]. Journal of Applied Poultry Research,2016,25(3):422-427.
[95]刁華杰,王芳芳,夏九龍,等. 日糧營養素對動物機體免疫的影響[J]. 黑龍江畜牧獸醫(下半月),2016(8):180-183.
[96]劉海港,韓 杰,洪 宇,等. 日糧營養素對動物免疫功能影響研究進展[J]. 飼料工業,2009,30(15):14-16.
[97]邱凌云. 飼養密度和飼糧添加25-羥基維生素D3對蛋雞生產性能、蛋品質和腸道健康的影響[D]. 雅安:四川農業大學,2019.
[98]張 龍. 禽宿主防御肽基因進化及其受丁酸和1,25(OH)2D3誘導表達研究[D]. 雅安:四川農業大學,2017.
[99]黨曉鵬. 蛋雞配合飼料中維生素E不同添加量的目標功能定位[J]. 今日畜牧獸醫,2015(10):38-40.
[100]Amevor F K,Cui Z F,Ning Z F,et al. Synergistic effects of quercetin and vitamin E on egg production,egg quality,and immunity in aging breeder hens[J]. Poultry Science,2021,100(12):101481.
[101]王天元. 維生素C和黃芪多糖對熱應激蛋雞蛋品質及免疫力的影響[D]. 太谷:山西農業大學,2016.
[102]布 英. 維生素C在蛋雞養殖中的應用[J]. 中國動物保健,2021,23(3):89,106.
[103]武笑天,溫 曌,李銳瑞,等. 葉酸對產蛋雞血液免疫指標和腸道屏障功能的影響[J]. 中國畜牧雜志,2021,57(8):174-181.
[104]成廷水. 氨基酸鋅對蛋雞免疫和抗氧化功能的調節作用及其應用研究[D]. 北京:中國農業大學,2004.
[105]Attia Y A,Al-Harthi M A,Abo El-Maaty H M.Calcium and cholecalciferol levels in late-phase laying hens:effects on productive traits,egg quality,blood biochemistry,and immune responses[J]. Frontiers in Veterinary Science,2020,7:389.
[106]Miao L P,Zhu M K,Li H Y,et al. Dietary high sodium fluoride impairs digestion and absorption ability,mucosal immunity,and alters cecum microbial community of laying hens[J]. Animals,2020,10(2):179.
[107]齊 茜,李佶隆,馬淑雪,等. 羥基蛋氨酸鋅對產蛋后期蛋雞生產性能、蛋品質和免疫相關基因表達的影響[J]. 動物營養學報,2018,30(12):4939-4946.
[108]苗灑灑,洪作鵬,朱明坤,等. 蛋氨酸錳對蛋雞抗氧化能力、脂質代謝、免疫功能和腸道形態的影響[J]. 中國畜牧雜志,2021,57(11):173-177,185.
[109]曲湘勇,彭燦陽,蔡 超,等. 不同水平有機微量元素對產蛋雞生產性能、血液生化及免疫指標的影響[J]. 中國家禽,2017,39(20):27-31.
[110]陳祥宇,朱亞昊,劉 萌,等. 復合植物提取物對蛋雞生產性能、蛋品質、免疫功能和抗氧化能力的影響[J]. 動物營養學報,2021,33(6):3271-3279.
[111]張桂鳳. 生姜對肉雞生產性能、抗氧化性能及肉品質影響的研究[D]. 泰安:山東農業大學,2010.
[112]時藝霖.紫蘇籽提取物對蛋雞產蛋高峰后期生產性能和免疫功能的影響[D]. 洛陽:河南科技大學,2015.
[113]Wu B B,Wang L Y,Jiang L L,et al. N-butanol extract from Folium isatidis inhibits the lipopolysaccharide-induced downregulation of CXCR1 and CXCR2 on human neutrophils[J]. Molecular Medicine Reports,2018,17(1):179-185.
[114]趙 晨,李海林,王明陽,等. 甜菜堿在家禽抗應激方面的作用研究進展[J]. 畜牧獸醫科學,2020(22):158-159.
[115]孫耀慧. 日糧中添加苜蓿草粉對蛋雞蛋品品質及免疫功能的影響[J]. 畜禽業,2015(6):18-19.
[116]陳祥宇,孫二東,陳一凡,等. 萬壽菊提取物對太行雞生產性能、蛋品質、免疫功能和抗氧化能力的影響[J]. 動物營養學報,2021,33(12):7160-7168.
[117]Shu G,Kong F L,Xu D,et al. Bamboo leaf flavone changed the community of cecum microbiota and improved the immune function in broilers[J]. Scientific Reports,2020,10(1):12324.
[118]張 健,楊亮宇,白文順. 復方中草藥添加劑對云南武定雛雞生長性能及免疫功能的影響[J]. 黑龍江畜牧獸醫,2015(5):79-81.
[119]李 維,張 蕓,龔 俞,等. 復方中草藥添加劑對種雞產蛋后期蛋品質的影響[J]. 江蘇農業科學,2021,49(7):165-170.
[120]鹿曉慧,謝全喜,侯楠楠,等. 微生態制劑對蛋雞生產性能及免疫機能的影響[J]. 現代畜牧獸醫,2021(6):53-55.
[121]Al-Khalaifah H S. Benefits of probiotics and/or prebiotics for antibiotic-reduced poultry[J]. Poultry Science,2018,97(11):3807-3815.
[122]李 偉. 飼料中添加復合微生物制劑對蛋雞生產性能與免疫機能的影響[J]. 中國飼料,2019(19):58-61.
[123]王國君,李文斌,向 榮.不同微生態制劑對京紅1號蛋雛雞生長性能和抗體水平的影響[J]. 養殖與飼料,2020(3):17-20.
[124]Ding X M,Li D D,Bai S P,et al. Effect of dietary xylooligosaccharides on intestinal characteristics,gut microbiota,cecal short-chain fatty acids,and plasma immune parameters of laying hens[J]. Poultry Science,2018,97(3):874-881.
[125]王紅衛,孫敏敏,孟 曉,等. 不同分子質量殼寡糖對蛋雞生產性能、腸道微生物及脾臟白細胞介素-2和腫瘤壞死因子-α基因表達的影響[J]. 動物營養學報,2013,25(11):2660-2667.
[126]沈家鯤,曹巖峰,梁先偉,等. 復合酸化劑對海蘭褐蛋雛雞生長、免疫性能和血清抗氧化能力的影響[J]. 中國家禽,2017,39(8):48-51.
[127]周 芬,吳義景,楊家軍,等. 抗菌肽對蛋雞生產性能、蛋品質及血液生化指標的影響[J]. 中國飼料,2021(13):43-46.
[128]Sun X F,Yue S Z,Qiao Y H,et al. Dietary supplementation with selenium-enriched earthworm powder improves antioxidative ability and immunity of laying hens[J]. Poultry Science,2020,99(11):5344-5349.
[129]顧動地,鄧在雙,任 曼,等. 蟲草粉聯合酵母硒對蛋雞產蛋高峰中期產蛋性能、蛋品質、抗氧化性和免疫功能的影響[J]. 中國畜牧獸醫,2021,48(7):2404-2414.
收稿日期:2022-10-14
基金項目:國家蛋雞產業技術體系建設專項(編號:CARS-40-S25);云南省西疇縣烏骨雞產業科技特派團項目(編號:202104BI090020)。
作者簡介:康佳佳(1996—),女,河南登封人,碩士,研究方向為動物遺傳育種與繁殖。E-mail:153730248@qq.com。
通信作者:葛長榮,博士,教授,研究方向為動物遺傳育種與繁殖。E-mail:gcrzal@126.com。
