干旱脅迫下外源內生真菌對燕麥種子萌發及幼苗生長的影響
2023-08-09 09:33:28李曉婷李立軍張永平郭云飛
灌溉排水學報 2023年7期
關鍵詞:影響
李曉婷,李立軍,張永平,郭云飛,韓 雪
(內蒙古農業大學 農學院,呼和浩特 010019)
0 引 言
【研究意義】干旱是制約我國農業可持續發展的主要非生物脅迫因素之一[1],尤其在作物生長早期會抑制種子萌發和降低出苗率[2],制約植物的生產力[3]。外源物質在提高種子萌發,促進作物生長和緩解干旱脅迫方面已有諸多研究,如外源生長物質[4-6]、礦物質元素[7]和納米顆粒[8-10]等。植物內生菌廣泛存在于植物體內,因其與宿主植物的相容性、再感染的便捷性和不同接種方式,較非生物外源物質更有利于農業的可持續發展[11-12]。因此,探究干旱脅迫下外源內生菌下種子萌發及幼苗生長的變化,對促進旱作區作物穩產高產具有重要意義。【研究進展】植物內生菌是存在于植物體內且不會引起明顯癥狀或對宿主造成損害的微生物類群[13]。作為新型的微生物資源,植物內生菌不僅具有較高的生態適應性,還能提高作物應對非生物脅迫的能力[14-15]。植物內生菌與植物共生體能提高宿主植物對干旱脅迫的耐受性,通過分泌有益的植物激素和其他代謝物促進植物生長,提高植物滲透調節和抗氧化能力[16-17]。在干旱脅迫下感染內生真菌的種子的發芽率高于未感染種子,且感染內生真菌的植物生物量和株高均優于未感染真菌植株。沙生梭梭的部分內生菌能夠提高種子的發芽率[18],并促進植物生長。內生菌的定殖能提高種子活力,改善干旱和重金屬鎘等非生物脅迫下種子發芽[19-20]。當前,內生真菌作為外源物質的研究主要集中在增強植物的抗逆性方面,而干旱條件下外源內生真菌如何提高種子萌發和改變根系空間結構的相關研究仍然較少。
燕麥(Avena sativaL.)是優質的糧飼兼用作物[21],作為內蒙古地區的特色作物,多種植在農牧交錯的干旱半干旱區,由于年均降水量偏少且分布不均,連年受到干旱的影響。【切入點】干旱缺水影響燕麥整個生育期的生長發育,尤其是在生長發育前期,表現為出苗率和成苗率低,造成該區燕麥產量較低。外源物質浸種是提高種子萌發的技術手段,而關于外源內生真菌對干旱脅迫下種子萌發和幼苗生長及生理特性變化的報道較少。【擬解決的關鍵問題】本研究利用PEG-6000 模擬干旱脅迫,結合不同內生真菌發酵液處理,探究正常水分和干旱脅迫下外源內生真菌對種子萌發和幼苗形態指標變化特征的影響,進一步揭示外源內生真菌提高植物耐旱特性的生理機制,以期為農業生產中外源植物內生真菌的合理應用提供理論依據。
1 材料與方法
1.1 試驗材料
供試內生真菌為鐮刀菌屬 O-2(Fusarium equiseti,登錄號 ON495943)和鏈隔孢屬 O-66(Alternaria pomicola,登錄號ON500608),由內蒙古農業大學燕麥產業研究中心從農牧交錯區燕麥中分離純化獲得,保存于-80 ℃超低溫冰箱。菌種預先在馬鈴薯葡萄糖培養基中(PDA)(馬鈴薯葡萄糖培養基20 g,瓊脂22 g,蒸餾水1 000 mL)培養7 d。供試內生真菌發酵培養基為Czapek 培養基(NaNO33 g、K2HPO41 g、MgSO4·7H2O 0.5 g、KCl 0.5 g、FeSO40.01 g、蔗糖30 g、蒸餾水1 000 mL)。供試作物為燕麥(Aνena satiνaL.),品種為燕科2 號。
1.2 內生真菌發酵液的培養和燕麥種子表面消毒
將在PDA 培養基上培養7 d 的內生真菌用打孔器打出直徑5 mm 的菌餅,用接種針挑取菌餅分別接種到250 mL 的錐形瓶中,每個錐形瓶含有100 mL 的Czapek 培養基,培養基中放置3 個菌餅,在恒溫振蕩箱中28 ℃、160 r/min 黑暗條件下培養7 d,調節發酵液的吸光度值(OD600nm)為1(約為1×108CFU/mL)。
燕麥種子的表面消毒在超凈工作臺中進行。選取籽粒飽滿、大小均勻的燕麥種子,置于75%乙醇中浸泡1 min,無菌水沖洗2 次,再用3%的NaClO溶液消毒5 min,接著用無菌水沖洗5 次,然后將種子置于無菌濾紙上晾干備用。
1.3 試驗設計
1.3.1 燕麥種子萌發試驗
試驗采用水培養基法進行種子萌發試驗。設2個水分處理,分別為正常水分(WW)和12%的PEG-6000 干旱脅迫處理(HD);向培養皿添加100μL 的O-2、O-66 內生真菌發酵液,以添加等量Czapek 培養基為對照(CK),共計3 種添加液,合計6 個處理,每個處理重復3 次。將消毒后的燕麥種子置于不同處理的培養皿中,每個培養皿(直徑90 mm)放置燕麥種子30 粒,將培養皿置于人工氣候箱中培養(光照25 ℃、16 h,黑暗20 ℃、8 h,濕度70%)。以種子胚芽長達到燕麥種子的1/2 為萌發標準,逐日觀察種子發芽情況并記錄種子發芽數,待連續2 d 種子發芽數不變視為種子發芽結束,持續觀察7 d。
1.3.2 干旱脅迫下外源內生真菌對燕麥幼苗影響的試驗
將表面消毒過的燕麥種子均勻置于消毒并含有2 層用無菌水浸潤濾紙的發芽盒(13 cm×19 cm×12 cm)中培養,待種子發芽5~7 d 后,選取長勢均勻一致的幼苗用海綿固定在35 孔育苗盤上,每孔1 株苗,同時用無菌的刀片割破燕麥的根系表皮,移栽到含有無菌1/4 的Hoagland 營養液的水培盒(1.25 L)中,水培盒大小與發芽盒一致,周圍用黑色膠帶裹纏,避免根系透光。每隔4 d 更換1 次營養液,并逐次提高Hoagland 營養液質量濃度。將幼苗置于光照充足、恒溫的育苗室中培養,并保持16 h 光照、8 h 黑暗,溫度分別控制在25、20 ℃,光溫交替周期循環。
試驗采用二因素隨機區組設計。將經過水培的燕麥苗分成3 組進行內生真菌處理,分別為對照(CK,添加10 mL 的Czapek 培養基)、O-2 和O-66內生真菌處理(分別添加10 mL 內生真菌發酵液)。在幼苗培養15 d 后進行模擬干旱脅迫處理,每個內生真菌處理的燕麥苗分為2 組,一組為Hoagland 營養液處理(WW),另一組為添加12%的PEG-6000的Hoagland 營養液(HD),共6 個處理,每個處理重復3 次。試驗期間定期隨機調整各處理燕麥幼苗的位置,并每隔4 d 更換1 次營養液。干旱脅迫15 d 后結束,隨即進行各項指標的測定。
1.4 外源內生真菌在植株體內的檢測
在種子萌發試驗結束后,取不同水分處理及外源內生真菌處理下的燕麥幼苗胚根檢測內生真菌的定殖情況。將燕麥幼苗的根系碎片經過無菌水沖洗后,置于95%乙醇內進行表面消毒10 s,然后在5%的NaClO 溶液浸泡30 s,再用無菌蒸餾水沖洗5次,使用無菌濾紙擦凈水分,無菌剪刀修剪根系兩端,使之成為約0.5 cm 的片段,然后將根段置于直徑90 mm 含有PDA 培養基的培養皿內,每個培養皿內放置15 個根片段,共重復3 次,在28 ℃培養箱內黑暗培養7 d,觀察根系中的內生真菌情況,并統計各內生真菌處理下根中的內生真菌侵染率。
1.5 測試指標及方法
根據許文博等[22]、王歡等[23]的方法計算種子發芽勢和發芽率,計算式分別為:
種子發芽結束后,從每個處理中隨機選取10 株燕麥,測定燕麥胚根長和胚芽長。
水培試驗結束后,每個處理隨機選取15 株燕麥幼苗測量株高;隨機選取9 株燕麥的根系用根系掃描儀(Epson Expression 10000XL,日本愛普生)和根系形態與結構分析應用系統WinRHIZO Pro2005b測定根系總長度、根系投影面積、根系總表面積、根系總體積和根尖數;取20 株燕麥幼苗置于牛皮紙袋中,105 ℃殺青30 min,85 ℃烘干至恒質量后稱量并記錄冠層和根干質量。
根系活力采用TTC 法測定[24],超氧化物歧化酶(SOD)采用氮藍四唑(NBT)光還原法測定[25],過氧化物歧化酶(POD)采用愈創木酚法測定[25],過氧化氫酶(CAT)采用過氧化氫法測定[26],丙二醛(MDA)采用硫代巴比妥酸法測定[27]。
1.6 數據分析
使用Excel 2019 整理數據并繪圖,采用IBM SPSS Statistics 25 進行方差分析,運用Origin 2021對幼苗形態指標進行主成分分析(PCA)。
2 結果與分析
2.1 外源內生真菌在燕麥根中的侵染率
2 個接種外源內生真菌處理的燕麥根系在各水分處理下均能發現內生真菌,如圖1 所示,圖中同一水分處理間不同外源內生真菌添加處理上不同小寫字母表示添加不同外源內生真菌處理的該項指標差異顯著,下同。在正常水分(WW)處理下,接種內生真菌根系侵染率均達到60%以上,干旱脅迫下燕麥幼苗根系的侵染率顯著低于正常水分處理(P<0.05),但侵染率均在40%以上,但添加外源內生真菌處理間的差異不顯著。
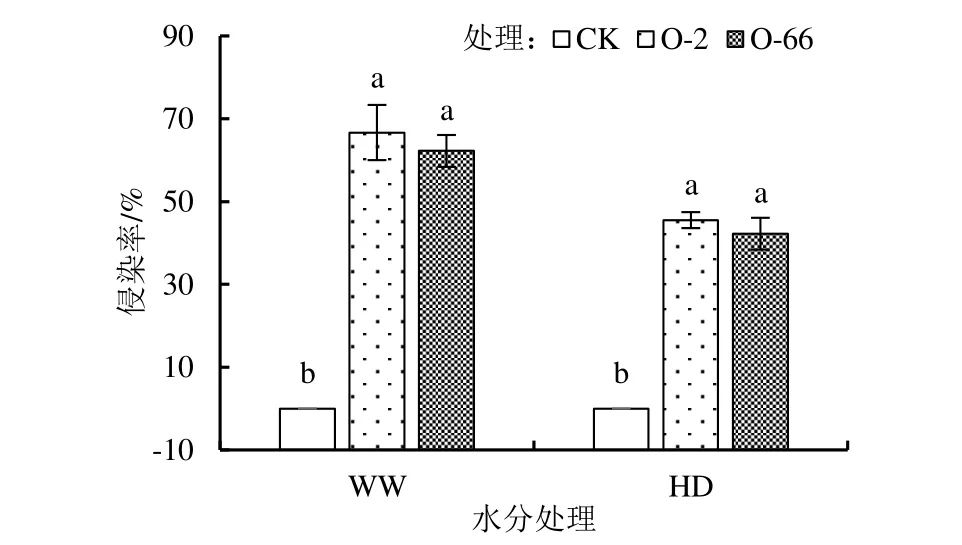
圖1 外源內生真菌在燕麥胚根中的侵染率Fig.1 Infection rate of exogenous endophytic fungi in oat radicle
2.2 外源內生真菌對不同水分處理下燕麥種子萌發的影響
外源內生真菌對干旱脅迫下燕麥發芽率和發芽勢的影響如圖2 所示。由圖2 可知,在正常水分處理(WW)下,添加外源內生真菌O-2、O-66 處理均提高了燕麥種子的發芽率,但與CK 差異不顯著;在干旱脅迫下(HD),添加外源內生真菌處理顯著提高了種子發芽率(P<0.05),分別較CK 提高了48.94%和44.68%(圖2(a))。正常水分處理(WW)下,添加外源內生真菌處理的燕麥種子發芽勢顯著高于CK;干旱脅迫下(HD),僅添加外源內生真菌O-66 處理較CK 顯著增加了41.18%(P<0.05)。水分和添加內生真菌均極顯著的影響種子的發芽率和發芽勢(P<0.001),二者的交互作用對種子發芽率達顯著水平(P<0.05),且極顯著的影響種子發芽勢(P<0.001)(表1)。

表1 水分處理和添加外源內生真菌對燕麥種子萌發指標雙因素方差分析Table 1 Two-factor variance analysis of oat seed germination indexes under different water treatments and exogenous endophytic fungi

圖2 不同水分處理下添加外源內生真菌對燕麥種子發芽率和發芽勢的影響Fig.2 Effects of exogenous endophytic fungi on germination rate and germination potential of oat seeds under different water treatments
2.3 外源內生真菌對不同水分處理下燕麥胚根長和胚芽長的影響
外源內生真菌對不同水分處理下燕麥胚根長和胚芽長的影響如圖3 所示。由圖3 可知,在正常水分(WW)和干旱脅迫處理下(HD),添加外源內生真菌均顯著增加了燕麥胚根長和胚芽長(P<0.05)。O-2、O-66 處理的胚根長在正常水分處理下(WW)分別是CK 的1.13 倍和1.19 倍,在干旱脅迫下(HD)分別是CK 的1.15 倍和1.36 倍。O-2、O-66 處理的胚芽長在正常水分(WW)處理下較CK 分別增加了14.65%和31.23%,而在干旱脅迫(HD)下分別較CK 增加了17.93%和42.45%。水分處理和添加外源內生真菌均極顯著影響胚根長和胚芽長(P<0.001),而二者交互作用對各指標的影響不顯著(表1)。
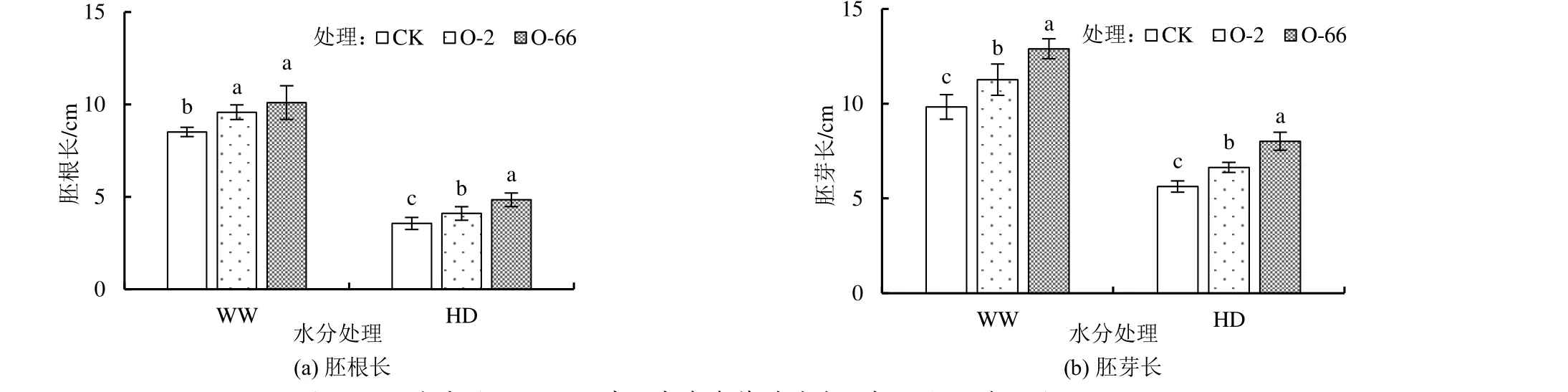
圖3 不同水分處理下添加外源內生真菌對燕麥胚根長和胚芽長的影響Fig.3 Effects of exogenous endophytic fungi on radicle and germ length of oat under different water treatments
2.4 外源內生真菌對不同水分處理下燕麥幼苗表型指標的影響
外源內生真菌對不同水分處理下燕麥幼苗形態指標的影響如圖4 所示。由圖4 可知,在正常水分處理下(WW),O-66 處理顯著提高了燕麥的株高、冠層干質量和根干質量,分別較CK 增加了21.41%、27.92%和30.63%(P<0.05)。在干旱脅迫下(HD),添加內生真菌O-2、O-66 均顯著提高了燕麥株高、冠層干質量和根干質量(P<0.05)。干旱脅迫(HD)下,CK 的株高和總干質量(冠層干質量+根干質量)分別較正常水分(WW)下降低了10.26%和34.63%,而 O-2 處理的株高和總干質量降低了 9.33%和33.69%,O-66 處理的株高和總干質量降低了8.59%和29.28%。故干旱脅迫(HD)下,O-2 處理燕麥根冠比顯著增加(P<0.05)。由表2 可知,干旱對燕麥各指標的影響均達到極顯著水平,添加內生真菌處理對燕麥株高、冠層干質量和根干質量的影響極顯著(P<0.001),對根冠比的影響顯著(P<0.05),而水分與添加內生真菌的交互作用對各指標的影響不顯著。
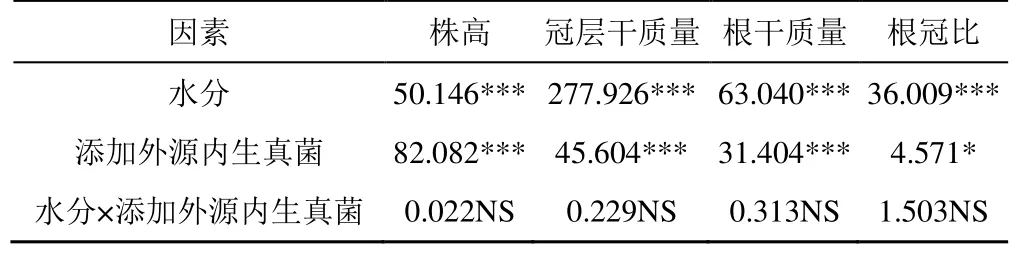
表2 外源內生真菌和水分處理對燕麥幼苗表型指標的雙因素方差分析Table 2 Two-factor variance analysis of exogenous endophytic fungi and water treatment on phenotypic indexes of oat seedlings

圖4 不同水分處理下添加外源內生真菌對燕麥幼苗表型指標的影響Fig.4 Effects of exogenous endophytic fungi on morphological indexes of oat seedlings under different water treatments
2.5 外源內生真菌對不同水分處理下燕麥幼苗根系的影響
外源內生真菌對不同水分處理下燕麥幼苗根系指標的影響如表3 所示。由表3 可知,正常水分處理下(WW),添加外源內生真菌O-2、O-66 處理均顯著提高了根系總長度、根系投影面積、根系總表面積和根尖數,而對根系總體積影響顯著的為O-66 處理。在干旱脅迫下(HD),添加外源內生真菌O-66處理顯著促進了根系總長度、根系投影面積、根系總表面積、根系總體積和根尖數的增加,分別較CK增加了20.40%、35.47%、31.84%、36.94%和59.24%。雙因素方差分析表明(表3),水分脅迫和內生真菌處理均對根系總長度、根系投影面積、根系總表面積和根系總體積有極顯著影響(P<0.001),而水分處理對根尖數的影響顯著(P<0.05),內生真菌對根尖數的影響達到極顯著水平(P<0.001)。水分和內生真菌的交互作用對根系總表面積(P<0.001)和根系總體積(P<0.01)的影響極顯著,對根系投影面積的影響達到顯著水平(P<0.05)。

表3 外源內生真菌對不同水分脅迫下的燕麥根系指標的影響Table 3 Effects of exogenous endophytic fungi on morphological indexes of oat seedlings under different water treatments
2.6 不同處理燕麥形態指標的主成分分析(PCA)
對各處理燕麥種子萌發及幼苗形態指標進行主成分分析(PCA)如圖5 所示,PC1 和PC2 分別解釋了總體方差的78.4%和9.6%,PC1 很好地把不同水分處理分開,WW 水分處理出現在第2、第3 象限,HD 水分處理出現在第1、第4 象限,根冠比與HD 水分處理關系密切,而其他指標均與WW 水分處理關系密切(圖5(a));對不同外源內生真菌處理下的燕麥指標進行主成分分析(圖5(b))發現,PC1 和PC2 分別解釋了總體方差的71.7%和13.3%,PC1 和PC2 分別將3 個處理明顯分離,添加外源內生真菌O-2 處理的根冠比最大,添加外源O-66 處理與根系結構指標、發芽率、發芽勢、干物質積累量等指標的關系密切,CK 出現在第4 象限,與各指標的距離均較遠。
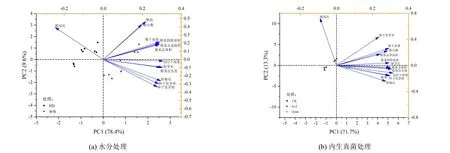
圖5 不同水分處理和不同外源內生真菌處理燕麥各形態指標的主成分分析Fig.5 Principal component analysis of oat indexes under different water and different exogenous fungal treatments
2.7 外源內生真菌對不同水分處理下燕麥生理特性的影響
內生真菌對不同水分處理下燕麥幼苗生理特性的影響如圖6 和表4 所示。干旱脅迫下(HD),添加外源內生真菌O-2、O-66 處理的根系活力顯著高于CK(P<0.05),分別增加了23.54%和15.34%。添加外源內生真菌處理的SOD、POD、CAT 活性均高于CK,在干旱脅迫下均顯著高于CK(P<0.05),而2 個添加外源內生真菌處理間差異不顯著。正常水分(WW)下,各處理的丙二醛(MDA)量變化差異不顯著;干旱脅迫下,添加外源內生真菌處理顯著降低了MDA 量(P<0.05),與CK 相比,O-2、O-66 處理分別降低了10.33%和9.27%。雙因素方差分析(表4)表明,水分和外源內生真菌交互作用顯著(P<0.05)影響對燕麥POD 活性和CAT 活性(P<0.05)。
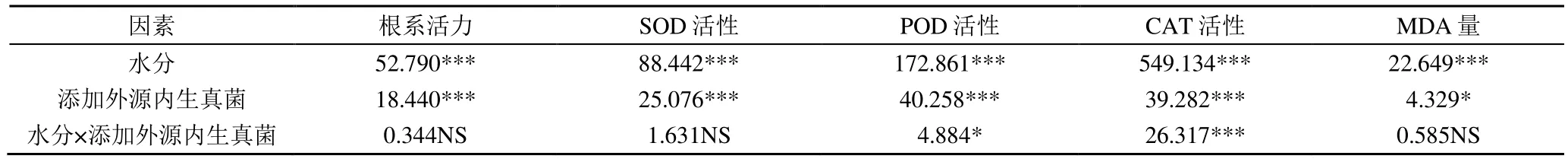
表4 水分和添加外源內生真菌對燕麥幼苗各指標雙因素方差分析Table 4 Two-factor variance analysis of indexes of oat seedlings under different water treatments and exogenous endophytic fungi
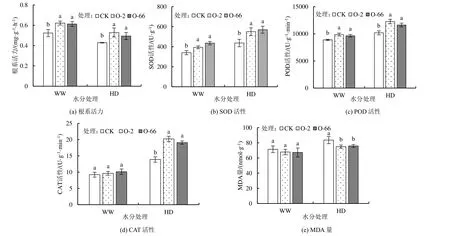
圖6 不同水分處理下添加外源內生真菌對燕麥生理指標的影響Fig.6 Effects of exogenous endophytic fungi on physiological indexes of oat under different water treatments
3 討 論
3.1 外源內生真菌對種子萌發的影響
種子萌發是植物生命周期中關鍵的階段之一[28],會受到多方面因素的影響[29-30]。內蒙古農牧交錯區是我國燕麥作物的主栽區,播種時期常會面臨風大雨少、土壤墑情差的情況,直接影響種子萌發。本研究采用PEG-6000 模擬干旱脅迫下接種內生真菌對種子萌發的影響,發現外源內生真菌O-2、O-66 均能定殖在燕麥的胚根上,干旱脅迫下接種外源內生真菌均提高了燕麥種子的發芽率和發芽勢,這說明外源內生真菌侵染種子后,增強了種子對脅迫的耐受性[31],緩解了干旱脅迫對種子萌發的抑制作用。本研究參與試驗的內生真菌分別是鐮刀菌屬(Fusariumsp.)和鏈格孢屬(Alternariasp.)對種子萌發均有積極的作用,這二類內生真菌的功能也從Qiang 等[32]和Khan 等[33]的研究中證實。內生真菌侵染后提高了草甸羊茅種子的發芽率,而對高羊茅種子的影響不顯著[34],甚至接種內生真菌鐮刀菌(Fusariumsp.)會降低種子發芽率[35]。這表明內生真菌對不同作物的種子萌發均有一定的促進或抑制作用,這可能與內生真菌與作物互作產生的代謝產物有關。在自然界中,內生真菌廣泛存在于植物中,經過長期的進化后一部分可能與作物互利共生,而另一部分在特定環境下會對寄主產生不利影響[36-37]。本研究發現接種內生真菌會顯著促進燕麥胚根和胚芽的生長。內生真菌對燕麥種子萌發的積極作用,可能是由于提高了種子的滲透調節能力,或產生生長調節物質,打破干旱脅迫對種子的休眠作用,提高種子活力,促進了種子萌發[38-39]。
3.2 外源內生真菌對燕麥水培幼苗生長的影響
本研究發現干旱脅迫會抑制燕麥幼苗的生長,而添加外源內生真菌O-2、O-66 處理的燕麥株高、冠層干質量和根干質量均顯著高于CK,故接種外源內生真菌會緩解干旱脅迫對燕麥生長的影響,這與Waqas 等[40]的研究一致,內生真菌在干旱脅迫下顯著提高了植株生物量和相關生長參數。干旱脅迫會破壞質膜系統,打破滲透平衡,抑制植物的生長。抗氧化酶可以清除活性氧物質,本研究發現,干旱脅迫下接種內生真菌均顯著增加了SOD、POD、CAT 酶活性,降低了MDA 量,這與Yogendra 等[17]的研究一致,表明外源內生真菌的定殖能提高抗氧化保護酶活性,抑制活性氧物質的累積,有效緩解干旱脅迫對細胞膜的損傷,提高燕麥的抗旱能力。
本研究中干旱脅迫顯著降低了根干質量和根系活力,而添加外源內生真菌提高了根干質量和根系活力,同時提高了根系總長度、總表面積、總體積和根尖數,說明接種內生真菌能改變根系結構,其原因可能是內生真菌最初先定殖在根中并產生促進根系生長的調節物質[41-42],提高了根系活力,也可能是由于內生真菌發酵液中的物質能夠間接影響植物的生長[43],這與Martinuz 等[44]和Verma 等[45-46]的研究結果一致。內生真菌與作物的生長存在一定的同步性,且會提高作物的耐旱性。植物根中的內生真菌可能會促進碳水化合物的積累,或改變根系生理生態,從而減輕干旱脅迫的影響[47]。
本研究中,外源內生真菌的應用緩解了干旱脅迫對燕麥種子萌發和幼苗生長的影響,且2 種外源內生真菌均對燕麥生長有積極的作用,該結果為外源內生真菌提高作物耐旱性的應用提供了理論基礎。同時,后續即將開展內生真菌對燕麥在萌發和幼苗生長階段植株代謝物變化、內源激素及滲透調節物質等方面的研究,從多角度揭示內生真菌提高燕麥耐旱性的機理。
4 結 論
1)干旱脅迫抑制了燕麥種子的萌發,而外源內生真菌處理提高了種子的發芽率,且能促進胚根和胚芽的伸長。
2)外源內生真菌的應用緩解干旱脅迫對植株的不利影響,提高了抗氧化酶活性,降低植株體內丙二醛量,改善根系結構,提高根系活力,促進幼苗生物量的累積。其中,添加外源內生真菌O-66 處理燕麥的抗旱效果較好。
(作者聲明本文無實際或潛在的利益沖突)
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
知識經濟·中國直銷(2016年3期)2016-02-27 16:15:49
現代檢驗醫學雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學低年級版)(2011年3期)2011-01-01 00:00:00
