陸地棉開(kāi)花整合子調(diào)控機(jī)制解析
2023-08-01 11:27:36陳麗婷閻媛媛
關(guān)鍵詞:植物
陳麗婷, 閻媛媛
(河北農(nóng)業(yè)大學(xué)農(nóng)學(xué)院,華北作物種質(zhì)資源研究與利用教育部重點(diǎn)實(shí)驗(yàn)室,河北省作物種質(zhì)資源重點(diǎn)實(shí)驗(yàn)室,河北 保定 071000)
棉花起源于亞熱帶地區(qū),是重要的纖維原料作物。野生棉生育期長(zhǎng)達(dá)180 d,其對(duì)熱量的需求限制著栽培區(qū)域。通過(guò)不斷的馴化培育,早熟棉生育期縮短至100 d左右,使得棉花栽培區(qū)域擴(kuò)張到46°N,且使糧棉輪作成為可能,極大地提高了土地利用率和經(jīng)濟(jì)效益[1]。早熟棉馴化的關(guān)鍵在于其對(duì)光周期及溫度的適應(yīng),使花芽分化和開(kāi)花提前。喻樹(shù)迅等[1]研究發(fā)現(xiàn),棉花早熟性是包含多個(gè)農(nóng)藝性狀(生育期、現(xiàn)蕾期、開(kāi)花期、吐絮期、第1果節(jié)位、霜前花率等)的綜合復(fù)雜性狀,其中現(xiàn)蕾期、開(kāi)花期是決定棉花早熟的關(guān)鍵時(shí)期。目前,種質(zhì)資源狹窄是早熟棉育種工作的瓶頸,挖掘調(diào)控棉花開(kāi)花相關(guān)的重要基因和解析開(kāi)花調(diào)控機(jī)制是發(fā)揮分子育種優(yōu)勢(shì)、加快早熟棉種質(zhì)創(chuàng)新的重要基礎(chǔ)。
植物開(kāi)花事件始于花芽分化的起始,標(biāo)志著植物從營(yíng)養(yǎng)生長(zhǎng)進(jìn)入生殖生長(zhǎng),是外因和內(nèi)因共同決定的結(jié)果。來(lái)自光周期途徑、春化途徑、環(huán)境溫度途徑、赤霉素途徑、自主途徑和年齡途徑的信號(hào)共同促進(jìn)開(kāi)花整合子基因LFY(LEAFY)、SOC1(SUPPRESSOR OF OVEREXPRESSION OF CO 1)和FT(FLOWERING LOCUS T)的轉(zhuǎn)錄,進(jìn)而激活花分生組織決定子,啟動(dòng)花芽分化[2]。
FT及其同系物TSF(twin sister of FT)屬于磷脂酰乙醇胺結(jié)合蛋白(phosphatidyl ethanolaminebinding proteins, PEBP),被視作植物“成花素”[3-4],受光周期和溫度的調(diào)控。擬南芥FT和TSF、番茄的SFT(single flower truss)和水稻的Hd3a(heading date 3a)[5-7]均在維管束韌皮部伴胞細(xì)胞中產(chǎn)生,隨后被運(yùn)輸?shù)巾敹朔稚M織(shoot apical meristem,SAM),與bZIP轉(zhuǎn)錄因子FD(flowering locus D)形成復(fù)合物,激活花分生組織特異性基因的表達(dá),包括AP1(APETALA1)和LFY。擬南芥FT基因研究的比較清楚,過(guò)表達(dá)FT能促進(jìn)擬南芥開(kāi)花,而其突變體明顯比野生型植株開(kāi)花晚[3]。水稻中FT同源蛋白Hd3a與GF14c和OsFD1形成激活復(fù)合體,促使水稻開(kāi)花,顯著縮短營(yíng)養(yǎng)生長(zhǎng)期[7]。Mcgarry等[8]發(fā)現(xiàn),在棉花中異位表達(dá)FT基因能促使其開(kāi)花,并影響花序。將GhFT1基因在擬南芥中異源表達(dá),促進(jìn)早花,并能部分挽救f(wàn)t-10突變體晚花表型[9];將其在煙草中異位表達(dá),同樣促進(jìn)煙草開(kāi)花,此外,呈現(xiàn)基部生長(zhǎng)更多的側(cè)生芽、葉片的形態(tài)發(fā)生改變等表型[10]。
SOC1基因編碼MADS蛋白,具有典型MIKC類(lèi)MADS-box轉(zhuǎn)錄因子的結(jié)構(gòu)特征,是關(guān)鍵的開(kāi)花調(diào)控和花器官?zèng)Q定因子。其整合了來(lái)自多個(gè)途徑的信號(hào),包括光周期途徑、春化途徑、赤霉素途徑和自主途徑[11]。在擬南芥中,SOC1受FLC(FLOWERING LOCUS C)、FT和CO基因調(diào)控[11-12]。超表達(dá)GhSOC1基因可以促進(jìn)擬南芥開(kāi)花,棉株株高變矮,其通過(guò)直接調(diào)控AP1/FUL-like基因促進(jìn)開(kāi)花[13]。
在擬南芥中,LFY作為植物花分生組織的決定子,整合了來(lái)自光周期途徑和赤霉素途徑的信號(hào)[14-15],同時(shí)也與其他MADS蛋白共同調(diào)控花發(fā)育程序[16-17]。從金魚(yú)草中克隆得到LFY基因的同系物FLO(FLORICAULA),并發(fā)現(xiàn)其影響花分生組織形成。擬南芥LFY基因不僅決定花分生組織,而且也影響植物從營(yíng)養(yǎng)生長(zhǎng)向生殖生長(zhǎng)的轉(zhuǎn)變,控制植物開(kāi)花時(shí)間。長(zhǎng)日照條件下,LFY在擬南芥幼苗的整個(gè)營(yíng)養(yǎng)生長(zhǎng)階段的葉原基中均有表達(dá)[18];經(jīng)成花誘導(dǎo)后,LFY基因活化,LFYmRNA起初積累于頂端花序分生組織的周邊區(qū),繼而強(qiáng)表達(dá)于花原基,而后在開(kāi)花期表達(dá)量急劇上升,其表達(dá)水平?jīng)Q定了植物開(kāi)花時(shí)間的早晚及花發(fā)育的成敗[19]。擬南芥轉(zhuǎn)LFY基因植株的側(cè)芽全部轉(zhuǎn)變?yōu)閱我坏幕ǎ议_(kāi)花時(shí)間大大提前[20]。在煙草[21]、山楊[20]、水稻[22]等物種中,LFY同源基因具有相同的功能。Li等[23]從陸地棉中克隆得到GhLFY基因,ChIP試驗(yàn)結(jié)果表明,GhLFY基因受GhSOC1的調(diào)控,GhLFY轉(zhuǎn)基因擬南芥形態(tài)發(fā)生了改變,莖生花替代了側(cè)枝,開(kāi)花提前。
葉片感受光照和溫度信號(hào),通過(guò)CO、SVP等蛋白及組蛋白修飾水平,精確調(diào)控FT基因的表達(dá)。葉片中FT蛋白通過(guò)維管束迅速移動(dòng)到頂端分生組織,激活SOC1基因的表達(dá),在AGL24的協(xié)助下,細(xì)胞質(zhì)中積累的SOC1蛋白進(jìn)入細(xì)胞核,結(jié)合到LFY基因的啟動(dòng)子區(qū)域,激活其轉(zhuǎn)錄[24]。SOC1在葉片和頂端分生組織中均大量表達(dá),其在頂端分生組織中如何激活花芽分化已經(jīng)明確。在葉片中,F(xiàn)T的大量積累會(huì)增加SOC1的轉(zhuǎn)錄[25],且SOC1受GA、春化路徑和內(nèi)源生長(zhǎng)信號(hào)的調(diào)控,但葉片中SOC1的積累并不影響FT的轉(zhuǎn)錄[6]。因此,葉片中SOC1具有整合信號(hào)的功能,但其如何影響花芽分化尚不清楚。
盡管有花植物的開(kāi)花整合子在進(jìn)化過(guò)程中相對(duì)保守,但仍存在物種特異性,如菊花葉片中受長(zhǎng)日照誘導(dǎo)的CsAFT移動(dòng)到莖尖后與FT復(fù)合體競(jìng)爭(zhēng),精確調(diào)控花芽分化的起始[26]。此外,對(duì)四倍棉基因組研究表明,A、D亞組上同源基因的轉(zhuǎn)錄和功能可能存在差異[27],因此明確四倍體棉花中開(kāi)花整合子的數(shù)目及功能,對(duì)解析棉花開(kāi)花調(diào)控機(jī)制至關(guān)重要,是早熟棉遺傳改良的理論基礎(chǔ)。本文鑒定了棉花基因組中開(kāi)花整合子,明確了其在二倍體和四倍體棉花基因組中的數(shù)目及進(jìn)化關(guān)系,分析了其在棉苗生長(zhǎng)過(guò)程中的時(shí)空表達(dá)特征,并明確了其功能及調(diào)控關(guān)系,為解析棉花開(kāi)花調(diào)控網(wǎng)絡(luò)奠定了分子基礎(chǔ)。
1 材料與方法
1.1 供試材料
陸地棉栽培種‘TM-1’和哥倫比亞野生型擬南芥由本實(shí)驗(yàn)室保存。
DNA聚合酶購(gòu)自南京諾唯贊生物科技公司,限制性?xún)?nèi)切酶購(gòu)自賽默飛公司,瓊脂糖凝膠回收試劑盒和質(zhì)粒提取試劑盒均購(gòu)自天根生物科技有限公司,植物RNA提取試劑盒購(gòu)自北京艾德萊生物科技公司,反轉(zhuǎn)錄試劑盒、Infusion連接酶購(gòu)自TaKaRa公司,熒光定量試劑盒購(gòu)自US EVERBRIGHT?INC公司,大腸桿菌感受態(tài)DH5α購(gòu)自北京天根生化科技有限公司,農(nóng)桿菌GV3103感受態(tài)為自制,pGreen載體由本實(shí)驗(yàn)室提供。引物合成與測(cè)序均由金唯智生物科技有限公司完成。
1.2 樣品準(zhǔn)備
將擬南芥以每穴3棵×3棵的密度種植于營(yíng)養(yǎng)土中,培養(yǎng)于光照培養(yǎng)室(23 ℃,16 h光照/8 h黑暗),在子葉展平后9 d取樣,用于基因表達(dá)分析。棉花培養(yǎng)于河北農(nóng)業(yè)大學(xué)溫室(28 ℃,12 h光照/8 h黑暗),2片真葉期開(kāi)始取樣,到4片真葉期結(jié)束,每次選取長(zhǎng)勢(shì)均勻一致的棉株葉片及頂端分生組織用于基因的實(shí)時(shí)定量表達(dá)分析及相關(guān)基因克隆。在4片真葉期取根、莖、葉、頂端,用于基因的組織表達(dá)特異性分析,所取材料立即投入液氮中冷凍,然后于-80 ℃冰箱保存?zhèn)溆谩@迷噭┖刑崛NA,使用TaKaRa RR047A試劑盒進(jìn)行反轉(zhuǎn)錄。
1.3 生物信息學(xué)分析
擬南芥氨基酸序列下載自TAIR網(wǎng)站(https://www.arabidopsis.org/),通過(guò)公開(kāi)的測(cè)序數(shù)據(jù)庫(kù)(NCBI)在線進(jìn)行blastP比對(duì)。在CottonFGD網(wǎng)站(https://cottonfgd.org/)進(jìn)行基因注釋分析,獲得與之同源的陸地棉開(kāi)花整合子基因。基于CDD數(shù)據(jù)庫(kù)(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)對(duì)其結(jié)構(gòu)域進(jìn)行預(yù)測(cè)。使用DNAMAN及ClustalX2軟件進(jìn)行多重序列比對(duì),使用MEGA7軟件中的鄰位相接法構(gòu)建進(jìn)化樹(shù)。
1.4 基因克隆和擬南芥的遺傳轉(zhuǎn)化
根據(jù)‘農(nóng)大棉8號(hào)’基因組序列[28]設(shè)計(jì)引物(表1),以‘TM-1’的cDNA為模板進(jìn)行克隆。使用Infusion連接酶將GhFT-2、GhSOC1-5和GhLFY-2連接到植物表達(dá)載體pGreen,構(gòu)建融合表達(dá)載體,轉(zhuǎn)化大腸桿菌DH5α后將陽(yáng)性克隆菌液送至金唯智測(cè)序,并保存測(cè)序正確的單克隆菌液以供后續(xù)試驗(yàn)使用。野生型(Col)擬南芥花蕾期沾取農(nóng)桿菌溶液(5%蔗糖,0.05% Silwet L-77),創(chuàng)建轉(zhuǎn)基因植株,并用Basta進(jìn)行篩選[25]。

表1 本研究中所用引物序列Table 1 Primer sequences in the study
1.5 轉(zhuǎn)基因擬南芥目的基因qRT-PCR
以擬南芥的AtTUB2、棉花的His作為內(nèi)參基因,利用Fast Super EvaGreen qPCR Master Mix進(jìn)行qRT-PCR,所用儀器為Applied Biosystems 7500實(shí)時(shí)定量PCR儀。每個(gè)樣品進(jìn)行3次試驗(yàn)重復(fù),采用2-ΔΔCt分析試驗(yàn)數(shù)據(jù)[29]。利用SPSS 11.0軟件處理數(shù)據(jù),Graphpad Prism 7.00軟件作圖。
2 結(jié)果與分析
2.1 棉花開(kāi)花整合子鑒定及序列分析
從NCBI網(wǎng)站下載毛白楊(Populus tomentos)、葡萄(Vitis vinifera)、擬南芥(Arabidopsis thaliana)和煙草(Nicotiana tabacum)的開(kāi)花整合子同源蛋白序列,cottonFGD在線比對(duì)(blast)棉花中同源序列,綜合全部已公開(kāi)棉花基因組序列及注釋?zhuān)b定棉花中FT、SOC1和LFY同源序列。因MADS家族成員結(jié)構(gòu)域高度保守,在鑒定SOC1同源序列時(shí),同時(shí)參考了棉花MADS家族成員鑒定結(jié)果[30-31]。綜合blast結(jié)果,‘農(nóng)大棉8號(hào)’基因組[28]中開(kāi)花整合子序列具有其他基因組序列的共性,故以‘農(nóng)大棉8號(hào)’基因組作為本文的參考序列。
棉花開(kāi)花整合子包含2個(gè)FT、6個(gè)SOC1、和2個(gè)LFY基因,分別命名為GhFT-1/GhFT-2、GhSOC1-1/GhSOC1-2/GhSOC1-3/GhSOC1-4/GhSOC1-5/GhSOC1-6和GhLFY-1/GhLFY-2。GhFTs包含保守基序[FYL]-x-[LVM]-[LIVF]-x-[TIVM]-[DC]-P-D-x-P-[SNG]-x(10)-H和決定FT功能的關(guān)鍵氨基酸位點(diǎn)Tyr85(Y)和Gln140(Q)(圖1A);GhSOC1s均含有MADS-box和K-box保守域,以及1個(gè)典型的SOC1 motif(圖1B);GhLFYs包含1個(gè)保守的FLOLFY結(jié)構(gòu)域(圖1C)。基因結(jié)構(gòu)分析可知,GhFTs基因包含4個(gè)外顯子和3個(gè)內(nèi)含子,GhSOC1s基因包含7個(gè)外顯子和6個(gè)內(nèi)含子,GhLFYs包含3個(gè)外顯子和2個(gè)內(nèi)含子,均與擬南芥的序列結(jié)構(gòu)一致。GhFTs基因結(jié)構(gòu)最為保守,GhLFY在A、D亞組上的序列也相對(duì)保守,但GhSOC1s在外顯子長(zhǎng)度上差異較大,暗示其功能在進(jìn)化中可能產(chǎn)生差異。

圖1 植物開(kāi)花整合子序列分析Fig.1 Sequence analysis of cotton floral integrator
2.2 棉花開(kāi)花整合子基因進(jìn)化分析
本研究分析了棉屬開(kāi)花整合子基因序列的進(jìn)化關(guān)系,發(fā)現(xiàn)陸地棉和海島棉中的FT和LFY基因各有2個(gè)成員,亞洲棉和雷蒙德氏棉中各有1個(gè)成員(圖2A、C),表明棉屬進(jìn)化過(guò)程中FT和LFY比較保守。不同于FT和LFY,在亞洲棉和雷蒙德式棉中分別含有3和2個(gè)SOC1序列,海島棉A、D亞組中共5個(gè)SOC1序列,且與二倍體棉的同源序列相似度高;值得注意的是,陸地棉較海島棉多了1個(gè)SOC1序列,除保留了二倍體棉中的5個(gè)SOC1序列外,來(lái)自A基因組的GhSOC1-3序列在D亞組增加了1個(gè)相似度高的序列GhSOC1-4,可能由片段復(fù)制而來(lái),該現(xiàn)象證明海島棉和陸地棉進(jìn)化事件的獨(dú)立性。此外,四倍體棉中SOC1-1/2/3/4序列與其對(duì)應(yīng)的二倍體棉中序列均能聚在1個(gè)進(jìn)化分支上,但GhSOC1-5/6序列較其二倍體中同源序列相似度略低,說(shuō)明GhSOC1-5/6序列在進(jìn)化過(guò)程中產(chǎn)生了變異(圖2B)。
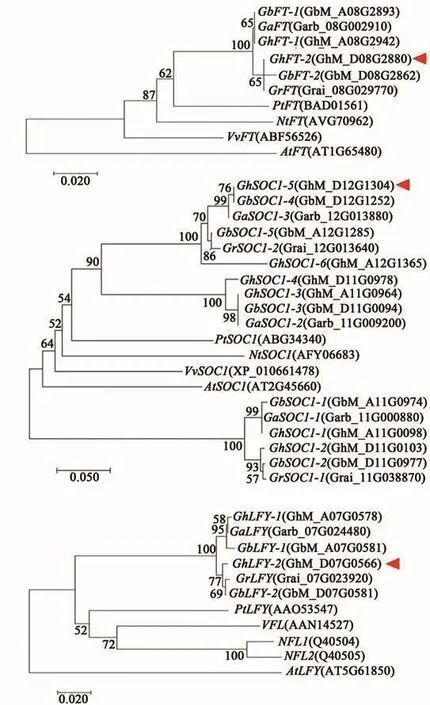
圖2 植物開(kāi)花整合子基因的進(jìn)化樹(shù)Fig.2 Phylogenetic analysis of plant floral integrator genes
綜上,陸地棉在進(jìn)化過(guò)程中,開(kāi)花整合子FT和LFY相對(duì)保守,而SOC1序列在陸地棉棉形成過(guò)程中數(shù)量增加,且基因編碼序列和非編碼序列均產(chǎn)生了變異,這可能與陸地棉對(duì)環(huán)境的適應(yīng)性增強(qiáng)有關(guān)。
2.3 棉花開(kāi)花整合子基因表達(dá)特征分析
植物葉片是感受環(huán)境并產(chǎn)生生物信號(hào)的主要部位,而花芽分化的起始部位在頂端分生組織,開(kāi)花調(diào)控基因的時(shí)空表達(dá)特征與其功能緊密相關(guān)。在擬南芥中,盡管FT、SOC1和LFY均促進(jìn)植物開(kāi)花,但它們的表達(dá)部位完全不重合。棉花3個(gè)開(kāi)花整合子基因均在頂端分生組織中優(yōu)勢(shì)表達(dá),且GhLFY-2在葉片中也有較高的表達(dá)(圖3A),三者的表達(dá)情況與擬南芥不盡相同,預(yù)示著棉花開(kāi)花調(diào)控網(wǎng)絡(luò)的物種特異性。隨著棉苗的不斷生長(zhǎng),在第4片真葉展平時(shí)(4thtrue leaf stage, 4TLS),3個(gè)開(kāi)花整合子基因在葉片和頂端分生組織中的表達(dá)均大幅升高,且在葉片中的升高更為劇烈,表明棉花TM-1在4TLS進(jìn)入生殖生長(zhǎng),3個(gè)開(kāi)花整合子基因在促進(jìn)植物開(kāi)花的功能上是保守的(圖3B和C)。
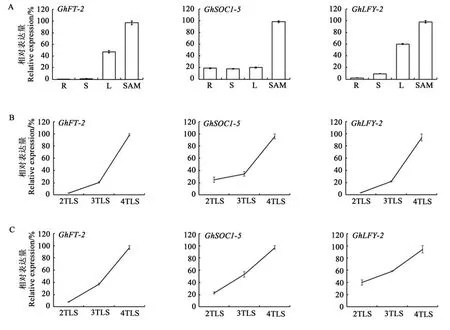
圖3 開(kāi)花整合子基因在陸地棉中表達(dá)Fig.3 Expression of floral integrators genes in Gossypium hirsutum
2.4 棉花開(kāi)花整合子基因?qū)χ参镩_(kāi)花的影響
為進(jìn)一步明確棉花開(kāi)花整合子基因的功能,分別克隆了GhFT-2、GhSOC1-5和GhLFY-2。GhFT-2與GhFT1的相似度達(dá)到了99%,預(yù)示其功能的保守性。SOC1有6個(gè)成員,GhSOC1-5編碼序列長(zhǎng)度為666 bp,與‘農(nóng)大棉8號(hào)’基因組一致,而比其他陸地棉參考基因組在3’端少15 bp,說(shuō)明‘農(nóng)大棉8號(hào)’基因組在該位點(diǎn)的組裝相對(duì)較優(yōu)。GhLFY-2與GhLFY大小一致,存在9個(gè)氨基酸差異,序列上二者相對(duì)保守。
分別構(gòu)建克隆序列的轉(zhuǎn)基因擬南芥,最后各獲得10個(gè)以上轉(zhuǎn)基因株系,與WT相比,過(guò)表達(dá)3個(gè)基因的擬南芥均呈現(xiàn)不同程度的早花表型(圖4A)。分別選取4個(gè)T3代純合株系進(jìn)行開(kāi)花時(shí)間及基因表達(dá)水平分析(圖4B),發(fā)現(xiàn)隨著基因表達(dá)量的升高,轉(zhuǎn)基因植株的開(kāi)花時(shí)間越早,即3個(gè)開(kāi)花整合子促進(jìn)開(kāi)花的功能均具有劑量效應(yīng)。其中,GhFT-2基因?qū)χ参镩_(kāi)花的促進(jìn)作用最強(qiáng),株系#1和#2抽薹最早(當(dāng)5~6片輪作葉形成時(shí)),開(kāi)花時(shí)間提早1倍(圖4C)。此外,在GhLFY-2轉(zhuǎn)基因植株的蓮座葉葉腋里伸出3~4朵單生花(圖4D),表明GhLFY影響花分生組織分化。

圖4 棉花開(kāi)花整合子促進(jìn)開(kāi)花Fig.4 Cotton flowering integrators promotes flowering
2.5 開(kāi)花整合子相互作用關(guān)系
選取生長(zhǎng)14 d的轉(zhuǎn)基因擬南芥,檢測(cè)其開(kāi)花相關(guān)基因的表達(dá)量,以探究棉花開(kāi)花整合子間的相互作用。從圖5可以看出,在GhFT-2轉(zhuǎn)基因的株系中開(kāi)花整合子AtSOC1和AtLFY的表達(dá)量都不同程度的上調(diào)(圖5A)。在GhSOC1-5轉(zhuǎn)基因的株系中,AtLFY的表達(dá)量顯著上調(diào),而AtFT基因表達(dá)量變化不大(圖5B)。在GhLFY-2轉(zhuǎn)基因的株系中,AtFT和AtSOC1均無(wú)明顯變化(圖5C)。由以上結(jié)果可推測(cè),棉花LFY基因位于FT和SOC1基因的下游,而FT可直接影響SOC1的表達(dá)(圖6)。

圖5 擬南芥開(kāi)花整合子基因轉(zhuǎn)錄水平Fig.5 Expression of floral integrator genes in transgenic Arabidopsis plants
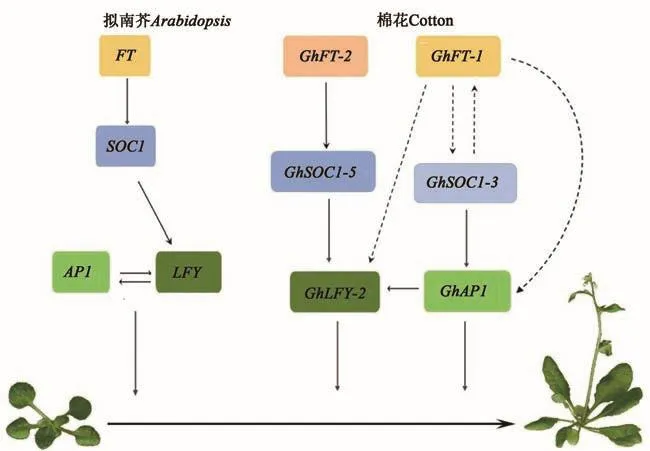
圖6 棉花開(kāi)花整合子調(diào)控機(jī)制模式Fig.6 Model of cotton flowering regulatory mechanism
3 討論
調(diào)控植物開(kāi)花的眾多基因相互作用,形成復(fù)雜的調(diào)控網(wǎng)絡(luò)。其中,SOC1、FT和LFY整合來(lái)自環(huán)境和內(nèi)源的各種信號(hào),最終激活花芽分化決定子的表達(dá),開(kāi)啟生殖生長(zhǎng)。陸地棉和海島棉是異源四倍體,由二倍體棉基因組融合而來(lái)。在馴化過(guò)程中,陸地棉開(kāi)花時(shí)間提前,使得栽培區(qū)域得以北移。本文鑒定了陸地棉、海島棉、亞洲棉和雷蒙德氏棉中的開(kāi)花整合子,其中FT和LFY基因在二倍體和四倍體棉基因組中相對(duì)保守,四倍體棉花中各有2個(gè)成員,二倍體棉花中各有1個(gè)成員。而在擬南芥中只有1個(gè)SOC1基因,但棉花中SOC1基因的數(shù)量加倍,海島棉保留了全部二倍體棉中的5個(gè)SOC1序列,而陸地棉D(zhuǎn)亞組增加了1個(gè)SOC1序列,共6個(gè)成員。且GhSOC1s編碼序列和基因組序列差異明顯,這可能與陸地棉對(duì)環(huán)境的適應(yīng)性有關(guān)。如藜麥隨著種植緯度的降低,其對(duì)長(zhǎng)日照的需求發(fā)生了變化,F(xiàn)T基因序列的多樣性與藜麥對(duì)光周期的適應(yīng)相關(guān)[32]。
以往的研究顯示,除了在根中很難檢測(cè)到GhFT1(Gh_D08G2407,GhFT-1)基因的表達(dá)外,在葉片、莖稈、花、胚珠等組織中都有表達(dá),但在葉片中的表達(dá)量最高[9],其在營(yíng)養(yǎng)組織中的表達(dá)模式與擬南芥AtFT一樣[8]。葉脈FT轉(zhuǎn)錄為mRNA并合成FT蛋白,F(xiàn)T蛋白再由韌皮部經(jīng)過(guò)長(zhǎng)距離運(yùn)輸?shù)巾敹朔稚M織,促進(jìn)下游SOC1基因的活躍轉(zhuǎn)錄。本研究超表達(dá)GhFT-2使擬南芥開(kāi)花時(shí)間減少1倍,與轉(zhuǎn)GhFT1基因的擬南芥超表達(dá)植株對(duì)比,GhFT-2對(duì)開(kāi)花的促進(jìn)作用較GhFT1強(qiáng),說(shuō)明陸地棉中A、D亞組中的FT基因功能均被保留。樊紅娟等[33]以3798品系(陸地棉零式果枝)和‘華中94’(正常果枝)為材料進(jìn)行組織特異性表達(dá)分析,發(fā)現(xiàn)正常果枝中GhFT1在各組織中表達(dá)水平差異并不大,但是在零式果枝中,其莖尖的表達(dá)量要明顯高于其他組織,其次在花芽中的表達(dá)量也相對(duì)較高,表明GhFT1可能參與花器官的發(fā)育。Mcgarry等[8]對(duì)擬南芥及棉花的FT基因進(jìn)行了超表達(dá)和VIGS(virus-induced gene silencing)沉默研究,發(fā)現(xiàn)GhFT1基因影響花分生組織的決定。本研究在棉花頂端分生組織中檢測(cè)到高水平的FT轉(zhuǎn)錄本,也預(yù)示GhFT調(diào)控開(kāi)花時(shí)間的機(jī)制不同于其他物種,GhFT對(duì)SOC1的調(diào)控不必依賴(lài)于葉片中FT蛋白的積累及向上運(yùn)輸,棉花頂端分生組織細(xì)胞可自己合成所需FT蛋白,促進(jìn)開(kāi)花,這可以使棉花對(duì)環(huán)境變化的響應(yīng)更迅速。
不同植物L(fēng)FY同源基因的時(shí)空表達(dá)模式不同。金魚(yú)草FLO基因只在開(kāi)花期表達(dá),四輪花器官中僅雄蕊中不表達(dá)[34]。在擬南芥、煙草、豌豆、葡萄等植物中,LFY同源基因在葉原基中及其開(kāi)花階段均有表達(dá)[35-38]。水稻RFL在幼穗中表達(dá),而在成熟的小花和葉片中不表達(dá),明顯有別于雙子葉植物[22]。這種時(shí)空上的表達(dá)差異反映了LFY同源基因的表達(dá)與植物本身花結(jié)構(gòu)的進(jìn)化有關(guān),同時(shí)表明LFY同源基因在植物的發(fā)育過(guò)程中具有重要的作用。本研究在棉花葉片中檢測(cè)到GhLFY的高水平轉(zhuǎn)錄,這不同于其他植物L(fēng)FY的空間表達(dá)特征。轉(zhuǎn)GhLFY-2植株開(kāi)花早,莖基有單生花,花器官正常,與GhLFY-1(Gh_A07G0404,GhLFY)的功能一致[23],說(shuō)明陸地棉A、D亞組GhLFYs促進(jìn)開(kāi)花、影響花序形成的生物學(xué)功能是保守的。葉片中GhLFY的轉(zhuǎn)錄可能參與了棉花成花誘導(dǎo),響應(yīng)環(huán)境或內(nèi)源信號(hào)。
Guo等[9]發(fā)現(xiàn),GhFT1轉(zhuǎn)基因植株中的開(kāi)花整合子SOC1和LFY以及AP1的表達(dá)量均顯著的提高,而本研究中FT-2僅促進(jìn)了LFY的轉(zhuǎn)錄,說(shuō)明陸地棉2個(gè)FT基因促進(jìn)開(kāi)花的功能是保守的,但調(diào)控機(jī)制不同。在擬南芥中,SOC1在頂端分生組織中直接激活LFY的轉(zhuǎn)錄,進(jìn)而激活A(yù)P1的轉(zhuǎn)錄,開(kāi)啟花芽分化。超表達(dá)GhSOC1-5顯著促進(jìn)了LFY的轉(zhuǎn)錄,表明棉花中FT-SOC1-LFY-AP1調(diào)控路徑的保守性。與本研究結(jié)果不同,超表達(dá)GhSOC1-3(Gh_A11G0755,GhSOC1)極大促進(jìn)了FT和AP1的轉(zhuǎn)錄,且GhSOC1-3可結(jié)合在AP1的基因序列上[13]。陸地棉中存在獨(dú)立于FT-SOC1-LFY-AP1的另1條開(kāi)花時(shí)間調(diào)控路徑,GhSOC1-3可直接調(diào)控AP1,且在GhSOC1-3和GhFT間存在正反饋調(diào)節(jié),即GhSOC1-3的積累會(huì)進(jìn)一步促進(jìn)GhFT的積累。綜上,陸地棉GhSOC1s參與不同的開(kāi)花調(diào)控路徑,這與其序列的多樣性相吻合,可能與陸地棉對(duì)環(huán)境的適應(yīng)有關(guān)。
本研究鑒定并對(duì)比了四倍體棉和二倍體棉基因組中開(kāi)花整合子的序列,發(fā)現(xiàn)FT和LFY在棉屬中的進(jìn)化很保守,而SOC1序列在陸地棉中的增多及序列的變異,導(dǎo)致陸地棉中既存在保守的FT-SOC1-LFY-AP1調(diào)控路徑,又特異性進(jìn)化出SOC1-AP1調(diào)控路徑。陸地棉開(kāi)花整合子調(diào)控路徑的豐富,可能與其對(duì)環(huán)境適應(yīng)性的提高相關(guān)。
猜你喜歡
少兒科學(xué)周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學(xué)閱讀指南·低年級(jí)版(2021年3期)2021-03-19 06:12:40
小天使·二年級(jí)語(yǔ)數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級(jí)語(yǔ)數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學(xué)苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫(huà)刊)(2017年11期)2018-01-09 19:15:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
