關嶺牛FABP1和FABP2基因克隆及其組織表達分析
2023-07-22 04:35:13石鵬飛許家利孫金魁許厚強
南方農業學報 2023年2期
石鵬飛 許家利 孫金魁 許厚強
摘要:【目的】明確肝臟型脂肪酸結合蛋白(FABP1)和腸型脂肪酸結合蛋白(FABP2)基因的分子特征及其在不同年齡段和不同品種牛各組織中的表達情況,為后續研究FABP1和FABP2對牛脂肪酸的調控作用機制提供理論依據。【方法】采用RT-PCR擴增關嶺牛FABP1和FABP2基因編碼區(CDS)序列,通過ProtParam、ProtScale、SOPMA、SWISS-MODEL、TMHMM-2.0、SignalP-5.0和NetPhos-3.1等在線軟件進行生物信息學分析,并以實時熒光定量PCR檢測FABP1和FABP2基因在3日齡關嶺牛(犢牛)、24月齡關嶺牛(成年牛)及24月齡雜交牛(關嶺牛×利木贊牛)各組織中的表達情況。【結果】FABP1、FABP2基因CDS序列全長分別為384和399 bp,對應編碼127和132個氨基酸殘基,其編碼蛋白二、三級結構均以無規則卷曲和延伸鏈為主,無跨膜結構域和信號肽剪切位點,且屬于親水性穩定蛋白。FABP1、FABP2氨基酸序列分別包含19和18個磷酸化位點,其中蘇氨酸在2個氨基酸序列中均廣泛分布。FABP1、FABP2氨基酸序列同源比對及系統發育進化分析結果均顯示,關嶺牛與馬、綿羊和山羊的親緣關系較近,而與雞和鴨的親緣關系較遠,與其他物種的相似性都在75.0%以上。FABP1、FABP2基因在不同年齡段和不同品種牛的各組織中均有表達,其中,FABP1基因以在肝臟和小腸中的相對表達量較高,且犢牛和雜交牛多個組織中的FABP1基因相對表達量極顯著高于成年關嶺牛(P<0.01,下同);FABP2基因則主要在小腸中高表達,且成年關嶺牛小腸中的FABP2基因相對表達量極顯著高于犢牛、顯著高于雜交牛(P<0.05)。【結論】FABP1和FABP2基因在關嶺牛中保守性較高,且在不同年齡階段及不同組織中均有表達,尤其在肝臟和小腸中的表達水平較高,故推測FABP1和FABP2基因協同調控關嶺牛脂肪酸的合成代謝。
關鍵詞: 關嶺牛;FABP1基因;FABP2基因;表達特征;脂肪酸
中圖分類號:S823.81? ? ? ? ? ? ? ? ? ? ? ? ?文獻標志碼:A 文章編號:2095-1191(2023)02-0598-11
Abstract:【Objective】The molecular characteristics of liver-type fatty acid binding protein (FABP1) and intestinal-type fatty acid binding protein (FABP2) genes and their expression in various tissues of cattle of different ages and breeds were clarified to provide a theoretical basis for subsequent studies on the mechanisms of the regulatory effects of FABP1 and FABP2 on bovine fatty acids. 【Method】The sequences of the coding regions (CDS) of FABP1 and FABP2 genes in Guanling cattle were amplified by RT-PCR, and bioinformatics analysis was performed by online softwares such as ProtParam, ProtScale, SOPMA, SWISS-MODEL, TMHMM-2.0, SignalP-5.0 and NetPhos-3.1, and the expression of FABP1 and FABP2 genes in various tissues of 3-day-old Guanling cattle (calves), 24-month-old Guanling cattle (adults) and 24-month-old crossbred cattle (Guanling cattle × Limousin cattle) was examined by real-time fluorescence quantitative PCR. 【Result】The full-length CDS sequences of FABP1 and FABP2 genes were 384 and 399 bp, respectively, encoding 127 and 132 amino acid residues. The secondary and tertiary structures of their encoded proteins were dominated by random coil and extended chain without transmembrane structural domains and signal peptide shear sites, and they were hydrophilic stable proteins. The amino acid sequences of FABP1 and FABP2 contained 19 and 18 phosphorylation sites respectively, of which threonine was widely distributed in both amino acid sequences. Homology comparison and phylogenetic evolutionary analysis of the amino acid sequences of FABP1 and FABP2 showed that Guanling cattle were closely related to horses, sheep and goats, while they were more distantly related to chickens and ducks, and the similarities with other species were also above 75.0%. FABP1 and FABP2 genes were expressed in all tissues of cattle of different ages and breeds, among which, the relative expression of FABP1 gene was higher in liver and small intestine, and the relative expression of FABP1 gene in several tissues of calves and crossbred cattle was extremely significantly higher than that of adult Guanling cattle (P<0.01, the same below). The FABP2 gene was highly expressed mainly in the small intestine, and the relative expression of FABP2 gene in the small intestine of adult Guanling cattle was extremely significantly higher than that of calves and significantly higher than that of crossbred cattle (P<0.05). 【Conclusion】FABP1 and FABP2 genes are highly conserved in Guanling cattle and are expressed in different age stages and tissues, especially in the liver and small intestine, so it is hypothesized that FABP1 and FABP2 genes synergistically regulate fatty acid anabolism in Guanling cattle.
Key words: Guanling cattle; FABP1 gene; FABP2 gene; expression characteristics; fatty acid
Foundation items: National Natural Science Foundation of China (31571279); Ministry of Education Program to Promote Regional Research Cooperation and High-level Talent Training with U.S. University (〔2015〕2062)
0 引言
【研究意義】肝臟型脂肪酸結合蛋白(L-FABP或FABP1)和腸型脂肪酸結合蛋白(I-FABP或FABP2)同屬于脂肪酸結合蛋白(Fatty acid binding protein,FABP)家族,該家族成員20%~70%的氨基酸序列表現出一致性,是一類與脂肪酸有高度親和性的小分子蛋白(Chmurzyńska,2006;Storch and Thumser,2010)。FABPs能與長鏈脂肪酸結合,調節脂肪酸合成與分解的方式,進而調控細胞內的脂類代謝(Ono and Odani,2010)。因此,探究牛FABP1和FABP2基因的分子生物學特征及其組織表達特性,可為揭示FABP1和FABP2對脂肪酸的調控機制提供理論依據。【前人研究進展】長鏈脂肪酸作為代謝能源和生物膜的組成成分,在一些信號轉導途徑中發揮著重要作用(Schaap et al.,2002)。目前,已發現10多種類型的FABPs,其中FABP1是最早發現的FABPs(Chan et al.,1985)。FABP1調控細胞內脂質代謝和極低密度脂蛋白分泌,對脂肪酸的攝入及肝細胞中脂肪酸和膽固醇的穩態起重要作用(Newberry et al.,2003,2009)。與其他FABPs不同的是,FABP1不僅能與長鏈脂肪酸結合,還能與多種疏水配體結合,包括長鏈脂酰CoA、血紅素、維生素K及膽固醇等(Ono and Odani,2010)。研究表明,敲除FABP1基因小鼠對脂肪酸的利用能力下降,但與長鏈脂酰CoA結合的相關蛋白表達量上升(Martin et al.,2003);而過表達FABP1基因能顯著提高肝細胞對脂肪酸的攝取(Wu et al.,2016)、脂肪生成(Mukai et al.,2017)和極低密度脂蛋白分泌(Atshaves et al.,2010)。此外,在下調FABP1基因表達的小鼠中極低密度脂蛋白分泌減少,且能有效抑制小鼠脂肪肝的形成(Spann et al.,2006)。正常狀態下,FABP1與脂肪酸的結合效率高于FABP2,但以飲食來源的脂肪酸為運輸目標時FABP2與脂肪酸的結合效率高于FABP1,使其更有利于對食物中脂類物質的吸收和運輸(Alpers et al.,2000),也說明FABP1和FABP2在脂肪酸的合成代謝過程中發揮著不同作用。FABP2能將細胞中的長鏈脂肪酸運送至內質網,合成三酰甘油,最后形成乳糜微粒并分泌到周圍的淋巴管中(Zimmerman et al.,2001)。Lackey等(2019)研究表明,敲除FABP2基因小鼠表現出腸道絨毛長度縮短、密度降低及腸道通透性增加,因此FABP2基因被認為是腸道完整性的重要標志基因。FABP1基因是調節家禽脂質代謝的重要候選基因,與過氧化物酶體增殖物激活受體α(Peroxisome proliferator activated receptor α,PPARα)協同調控脂肪酸的合成與分解,在肝臟和小腸的脂質代謝中發揮重要作用(He et al.,2013)。FABP1基因多態性與家禽肌內和腹部脂肪的沉積顯著相關(He et al.,2012);FABP2基因多態性除了與雞的腹脂重和腹脂率顯著相關(王啟貴,2004)外,還顯著影響北京鴨的體尺和屠宰性狀(廖秀冬等,2012)。綜上所述,FABP1和FABP2基因可能是家禽脂肪性狀的重要候選基因。【本研究切入點】目前,有關FABP1、FABP2基因的研究主要集中在人類和小鼠上,而針對牛FABP1和FABP2基因的生物功能及其表達特征尚未明確。【擬解決的關鍵問題】通過生物信息學分析關嶺牛FABP1和FABP2基因的分子結構及其功能,運用實時熒光定量PCR檢測這2個基因在不同年齡段和不同牛品種各組織中的表達差異,以期為后續研究FABP1和FABP2對牛脂肪酸的調控作用機制提供理論依據。
1 材料與方法
1. 1 試驗材料
試驗動物來自貴州省安順市關嶺縣關嶺牛產業園,隨機選取相同飼養條件下的3日齡關嶺牛(犢牛)、24月齡關嶺牛(成年牛)及24月齡雜交牛(關嶺牛×利木贊牛)各3頭,采集心臟、肝臟、脾臟、肺臟、腎臟、背最長肌、脂肪及小腸等組織樣品,-80 ℃保存備用。TRIzol購自美國Gibco公司;DL2000 DNA Marker、2×Taq PCR StarMix、逆轉錄試劑盒(Gen Star)購自西寶生物科技(上海)股份有限公司;熒光定量試劑Power UP SYBR GREEN Master Mix購自美國Thermo Fisher公司;膠回收試劑盒購自OMEGA公司;pMD19-T載體購自TaKaRa公司。主要儀器設備有電泳儀(DYY-2C,北京六一儀器廠)、超微量紫外分光光度計(NanoDrop 2000,美國Thermo Fisher公司)、普通PCR儀(C1000 Touch TM)、熒光PCR儀(Biosens SC710)及凝膠成像系統(Universal Hood Ⅱ,Bio-Rad)。
1. 2 引物設計與合成
根據NCBI已發布的牛FABP1基因序列(NM_175817.3)和FABP2基因序列(NM_001025332.1),使用Primer Premier 5.0設計編碼區(CDS)擴增引物及熒光定量PCR特異性引物,綜合選擇得分最高的引物序列,委托北京擎科生物科技股份有限公司合成,引物序列信息見表1。
1. 3 組織總RNA提取及cDNA合成
采用TRIzol法提取各組織總RNA,以超微量分光光度計測定總RNA濃度及OD值;使用逆轉錄試劑盒(Gen Star)將符合條件的總RNA反轉錄合成cDNA第一鏈,-20 ℃保存備用。
1. 4 FABP1和FABP2基因克隆及測序分析
通過特異性引物擴增關嶺牛FABP1和FABP2基因CDS區。PCR反應體系10.0 μL:cDNA模板0.4 μL,上、下游引物(10 μmol/L)各0.4 μL,2×Taq PCR StarMix 5.0 μL,ddH2O 3.8 μL。擴增程序:94 ℃預變性2 min;94 ℃ 30 s,58 ℃ 30 s,72 ℃ 60 s/kb,進行35個循環;72 ℃延伸10 min。以1.0%瓊脂糖凝膠電泳檢測PCR擴增產物,使用膠回收試劑盒進行膠回收,純化后的擴增產物與pMD19-T載體16 ℃連接30 min,然后轉化DH5α感受態細胞,加入液體培養基培養1.5 h,再涂布至固體培養基上,37 ℃過夜培養,經氨芐青霉素篩選后挑選單個菌落進行擴大培養,PCR鑒定呈陽性的菌液送至北京擎科生物科技股份有限公司測序。
1. 5 FABP1和FABP2基因生物信息學分析
通過ProtParam和ProtScale分析FABP1、FABP2蛋白理化性質及親/疏水性;采用SOPMA和SWISS-MODEL分別預測FABP1、FABP2蛋白二級結構及三級結構;利用TMHMM-2.0、SignalP-5.0和NetPhos-3.1預測FABP1、FABP2蛋白跨膜結構域、信號肽及磷酸化位點;使用MegAlign和MEGA 7.0分析FABP1、FABP2氨基酸序列相似性并構建系統發育進化樹。
1. 6 實時熒光定量PCR檢測FABP1和FABP2基因組織表達特征
以GAPDH為內參基因,通過實時熒光定量PCR檢測犢牛、成年關嶺牛及雜交牛各組織中的FABP1和FABP2基因表達情況。實時熒光定量PCR反應體系10.0 μL:Power UP SYBR GREEN Master Mix 5.0 μL,上、下游引物(10 μmol/L)各0.4 μL,cDNA模板0.5 μL,滅菌水3.7 μL。擴增程序:95 ℃預變性2 min;95 ℃ 15 s,60 ℃ 15 s,72 ℃ 60 s,進行39個循環。熔解曲線為儀器默認設置,每個樣品設3個重復,獲得完整光滑的熔解曲線,采用2-ΔΔCt法換算目的基因相對表達量。
2 結果與分析
2. 1 關嶺牛FABP1和FABP2基因CDS序列擴增結果
以關嶺牛cDNA為模板,擴增關嶺牛FABP1和FABP2基因CDS序列,經1.0%瓊脂糖凝膠電泳檢測,結果顯示擴增片段大小與預期結果相符(圖1),初步判斷成功擴增獲得關嶺牛FABP1和FABP2基因目的CDS序列。
2. 2 關嶺牛FABP1和FABP2基因克隆及測序結果
菌液PCR鑒定結果顯示成功獲得關嶺牛FABP1和FABP2基因的目的片段(圖2),測序結果與NCBI已發布的牛FABP1和FABP2基因進行比對分析,結果(圖3)發現,關嶺牛FABP1基因CDS序列第273位核苷酸存在堿基突變(T→G),為同義突變,氨基酸序列未發生改變;關嶺牛FABP2基因CDS序列與NCBI已發布的牛FABP2基因核苷酸序列完全一致,不存在堿基突變位點,表明pMD19-T-FABP1和pMD19-T-FABP2克隆載體構建成功。
2. 3 關嶺牛FABP1和FABP2基因生物信息學分析結果
2. 3. 1 關嶺牛FABP1和FABP2蛋白理化性質分析結果 ProtParam和ProtScale預測結果顯示,關嶺牛FABP1基因CDS序列長384 bp,共編碼127個氨基酸殘基,其蛋白分子式為C629H1009N165O195S7,原子數為2005,分子量為14227.34 Da,含量最高的氨基酸為賴氨酸(占12.6%),含量最低的為組氨酸(占0.8%)、半胱氨酸(占0.8%)和脯氨酸(占0.8%),理論等電點(pI)為7.78,蛋白不穩定指數為9.99,屬于穩定蛋白。FABP1蛋白氨基酸序列的第55位甘氨酸疏水性最強(1.522)、第10位谷氨酰胺親水性最強(-2.133),親水性總體平均值為-0.402,即FABP1蛋白為親水性蛋白(圖4-A)。FABP2基因CDS序列長399 bp,共編碼132個氨基酸殘基,其蛋白分子式為C674H1065N179O202S4,原子數為2124,分子量為15036.19 Da,含量最高的氨基酸為谷氨酸(占10.6%)和賴氨酸(占10.6%),含量最低的為組氨酸(占0.8%),pI為6.62,蛋白不穩定指數20.27,屬于穩定蛋白。FABP2蛋白氨基酸序列的第65位亮氨酸疏水性最強(2.000)、第14位天冬酰胺親水性最強(-2.711),親水性總體平均值為-0.414,也屬于親水性蛋白(圖4-B)。
2. 3. 2 關嶺牛FABP1和FABP2蛋白二、三級結構預測結果 通過SOMPA預測得知,FABP1蛋白二級結構由延伸鏈、無規則卷曲、α-螺旋和β-轉角4種結構構成(圖5-A),其中延伸鏈占比最高,為39.37%;FABP2蛋白同樣由延伸鏈、無規則卷曲、α-螺旋和β-轉角4種結構組成(圖5-B),也是以延伸鏈占比最高(36.36%)。SWISS-MODEL預測得到的關嶺牛FABP1、FABP2蛋白三級結構與對應的二級結構預測結果相符(圖6)。
2. 3. 3 關嶺牛FABP1和FABP2蛋白跨膜結構域、信號肽及磷酸化位點預測結果 通過TMHMM-2.0和SignalP-5.0分別預測關嶺牛FABP1、FABP2蛋白的跨膜結構域及信號肽,結果顯示二者均無跨膜結構域(圖7)和信號肽剪切位點(圖8)。NetPhos-3.1預測結果顯示,關嶺牛FABP1氨基酸序列共有19個磷酸化位點(圖9-A),包括6個絲氨酸磷酸化位點、11個蘇氨酸磷酸化位點和2個酪氨酸磷酸化位點;關嶺牛FABP2氨基酸序列共有18個磷酸化位點(圖9-B),包括5個絲氨酸磷酸化位點、9個蘇氨酸磷酸化位點和4個酪氨酸磷酸化位點;其中蘇氨酸磷酸化位點在2個氨基酸序列中均廣泛分布。
2. 4 不同物種FABP1和FABP2氨基酸序列同源比對分析結果
對多個物種的FABP1和FABP2氨基酸序列進行同源比對分析,結果表明,關嶺牛FABP1氨基酸序列與人類、小鼠、大鼠、馬、豬、綿羊、山羊、雞和鴨的FABP1氨基酸序列相似性分別為81.1%、76.4%、79.5%、96.1%、83.5%、97.6%、95.3%、67.7%和67.7%(圖10-A);關嶺牛FABP2氨基酸序列與人類、小鼠、大鼠、馬、豬、綿羊、山羊、雞和鴨的FABP2氨基酸序列相似性分別為84.1%、78.8%、82.6%、98.5%、87.9%、99.2%、99.2%、73.5%和73.5%(圖10-B)。基于FABP1和FABP2氨基酸序列相似性,分別利用MEGA 7.0構建系統發育進化樹(圖11),結果均顯示關嶺牛與馬、綿羊和山羊的親緣關系較近,而與雞和鴨的親緣關系較遠。
2. 5 FABP1和FABP2基因在犢牛、成年關嶺牛和雜交牛不同組織中的表達情況
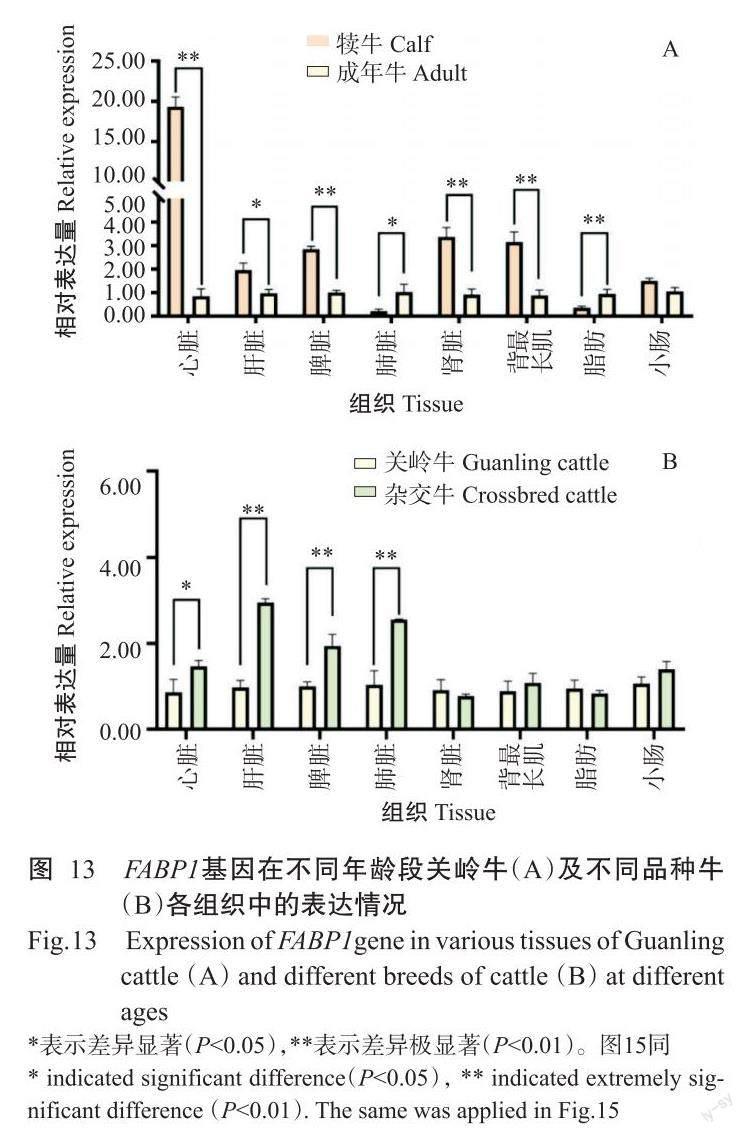
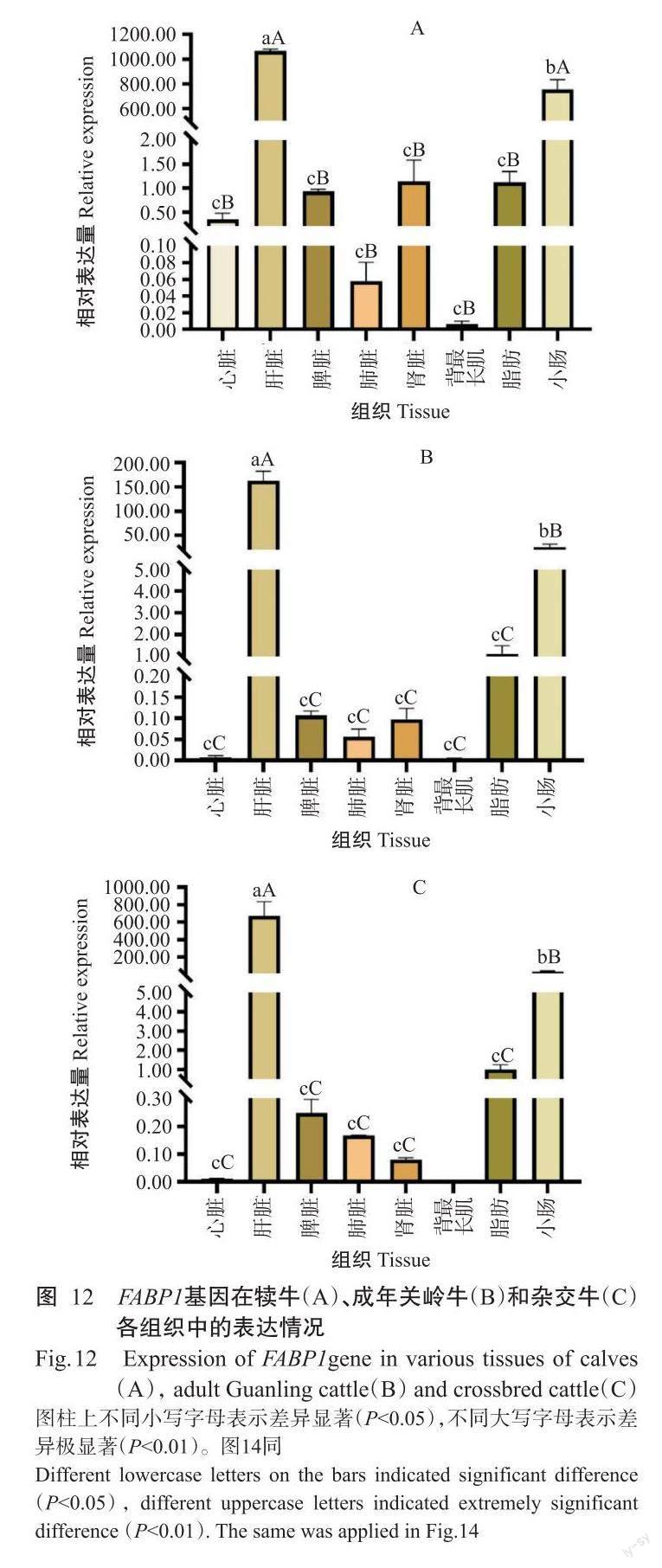
2. 5. 1 FABP1基因在犢牛、成年關嶺牛和雜交牛不同組織中的表達情況 實時熒光定量PCR檢測結果(圖12)顯示,FABP1基因在犢牛、成年關嶺牛和雜交牛的不同組織中均有表達,且以肝臟中的相對表達量最高。FABP1基因在犢牛中的相對表達量排序為肝臟>小腸>腎臟>脂肪>脾臟>心臟>肺臟>背最長肌(圖12-A),其中肝臟和小腸中的相對表達量極顯著高于其他組織(P<0.01,下同);FABP1基因在成年關嶺牛中的相對表達量排序為肝臟>小腸>脂肪>脾臟>腎臟>肺臟>心臟>背最長肌(圖12-B),在肝臟和小腸中的相對表達量極顯著高于其他組織;FABP1基因在雜交牛中的相對表達量排序為肝臟>小腸>脂肪>脾臟>肺臟>腎臟>心臟>背最長肌(圖12-C),在肝臟和小腸中的相對表達量也極顯著高于其他組織。此外,FABP1基因在犢牛心臟、脾臟、腎臟和背最長肌中的相對表達量極顯著高于成年關嶺牛,而在雜交牛肝臟、脾臟和肺臟中的相對表達量極顯著高于關嶺牛(圖13)。
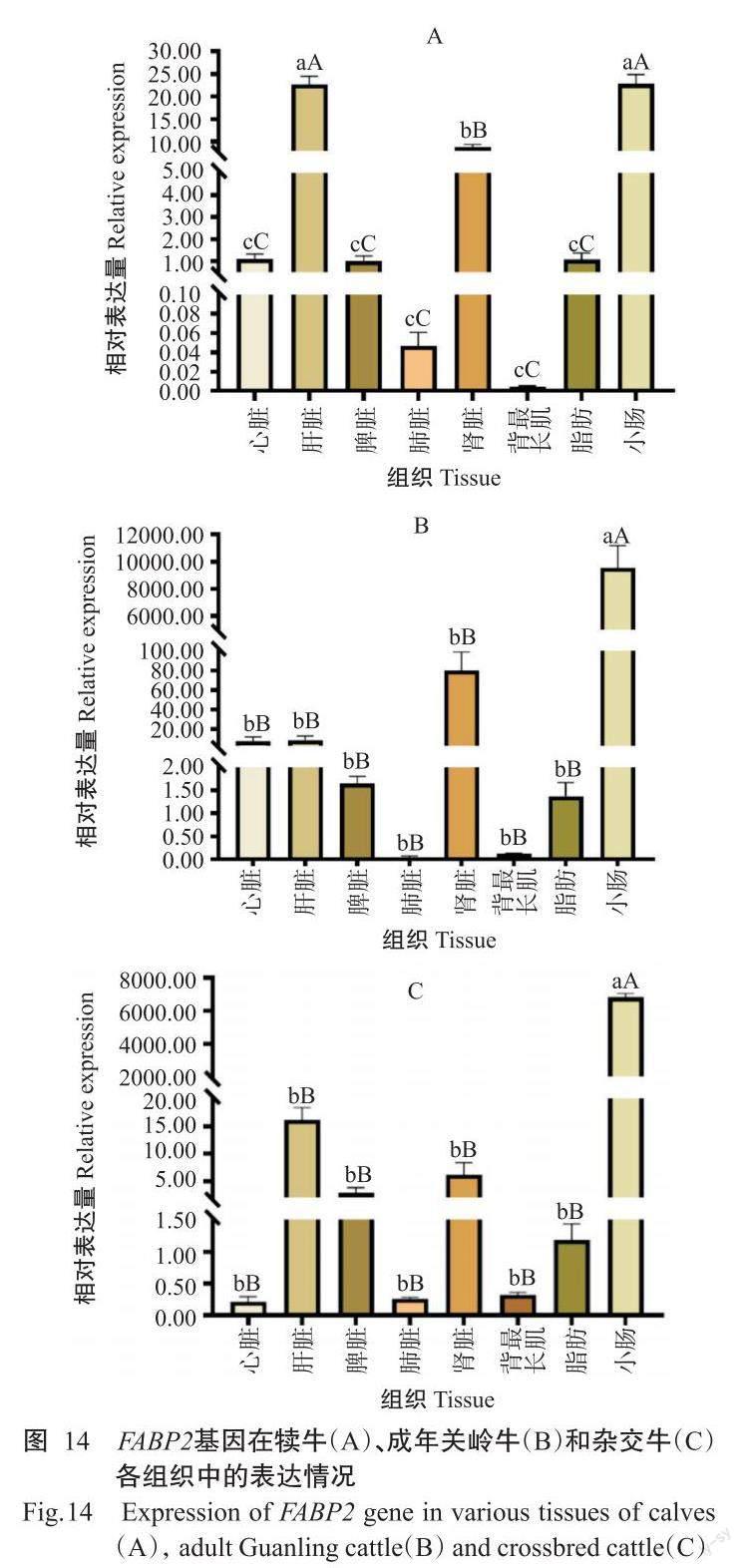
2. 5. 2 FABP2基因在犢牛、成年關嶺牛和雜交牛不同組織中的表達情況 實時熒光定量PCR檢測結果(圖14)顯示,FABP2基因在犢牛、成年關嶺牛和雜交牛的不同組織中均有表達,且以小腸中的相對表達量最高。FABP2基因在犢牛中的相對表達量排序為小腸>肝臟>腎臟>心臟>脂肪>脾臟>肺臟>背最長肌(圖14-A),在小腸和肝臟中的相對表達量極顯著高于其他組織;FABP2基因在成年關嶺牛中的相對表達量排序為小腸>腎臟>肝臟>心臟>脾臟>脂肪>背最長肌>肺臟(圖14-B),在雜交牛中的相對表達量排序為小腸>肝臟>腎臟>脾臟>脂肪>背最長肌>肺臟>心臟(圖14-C),FABP2基因在成年關嶺牛和雜交牛小腸中的相對表達量均極顯著高于其他組織。此外,FABP2基因在犢牛肝臟、肺臟和脂肪中的相對表達量顯著高于成年關嶺牛,但在小腸和背最長肌中表現為成年關嶺牛極顯著高于犢牛;FABP2基因在關嶺牛心臟、脾臟、腎臟和脂肪中的相對表達量極顯著高于雜交牛(圖15)。
3 討論
FABP1是FABP家族的重要成員之一,不僅可調控脂肪酸代謝,還影響細胞的生長。在細胞有絲分裂各時期,FABP1基因表達水平顯著上調,且在切除部分肝臟組織后的再生過程中FABP1基因表達也呈顯著上調趨勢(Wang et al.,2004)。此外,FABP1對細胞具有保護作用,正常狀態下可結合一些潛在的有毒脂肪酸、血紅素及其他分子,解除結合即造成嚴重的細胞毒性(Wang et al.,2015)。在人類、獼猴及其他哺乳動物中,FABP2基因特異性表達于營養物質的主要吸收場所——小腸,與FABP2對食物中長鏈脂肪酸的吸收、轉運及脂類物質代謝密切相關(Gajda and Storch,2015)。Baier等(1995)還研究證實,人類FABP2基因多態性與高血脂、糖尿病和肥胖癥等疾病顯著相關。目前,有關FABP1和FABP2基因的研究主要涉及人類的疾病及其免疫,在家畜上尤其是針對牛的研究鮮見報道。
本研究成功克隆出關嶺牛FABP1和FABP2基因CDS序列,與NCBI已公布的相應序列對比分析發現,關嶺牛FABP1基因核苷酸序列存在突變位點,但屬于同義突變,不改變氨基酸序列,關嶺牛FABP2基因CDS序列與NCBI已發布的牛FABP2基因核苷酸序列完全一致,表明FABP1和FABP2基因在關嶺牛上遺傳較穩定;FABP1和FABP2蛋白二、三級結構主要由延伸鏈和無規則卷曲構成,無規則卷曲是蛋白肽鏈中與配體和受體結合的重要部分(趙采芹等,2022),可能是FABP1和FABP2蛋白能有效結合并運輸脂肪酸的原因之一。關嶺牛FABP1、FABP2蛋白均無跨膜結構域和信號肽剪切位點,故推測二者不屬于分泌蛋白。基于FABP1和FABP2氨基酸序列相似性構建的系統發育進化樹均顯示,關嶺牛與馬、綿羊和山羊的親緣關系較近,而與雞和鴨的親緣關系較遠,與其他物種的相似性都在75.0%以上。可見,關嶺牛FABP1和FABP2基因保守性較高。
實時熒光定量PCR檢測結果表明,FABP1、FABP2基因在關嶺牛和雜交牛各組織中均有不同程度的表達,FABP1基因以在肝臟和小腸中的相對表達量較高,FABP2基因則主要在小腸中高表達,與姜延志和李學偉(2006)、Wang等(2019)的研究結果一致。肝臟和小腸是脂肪酸合成、運輸的重要部位,調控機體的脂質代謝而影響肌內脂肪含量沉積,與肉品質密切相關(Jensen et al.,1990;Hunter,2001)。已有研究表明, FABP1基因啟動子區含有過氧化物酶體增殖物受體元件,轉錄因子PPARα與之結合后調控FABP1基因表達(Venkatachalam et al.,2017),在肝臟和小腸中PPARα也呈豐富表達(Nakamura et al.,2014)。FABP1基因在犢牛多個組織(心臟、脾臟、腎臟和背最長肌)中的相對表達量極顯著高于成年關嶺牛,暗示著FABP1基因在關嶺牛的早期生長發育過程中發揮重要作用。FABP1基因在雜交牛肝臟、脾臟和肺臟中的相對表達量極顯著高于成年關嶺牛,可能是FABP1基因對雜交牛的影響大于關嶺牛。與成年關嶺牛相比,FABP2基因在犢牛肝臟、肺臟和脂肪中的相對表達量極顯著高于成年關嶺牛,與FABP1基因在關嶺牛中的表達情況相似;在成年關嶺牛心臟、脾臟、腎臟和脂肪中的FABP2基因相對表達量極顯著高于雜交牛,故推測FABP2基因對關嶺牛的影響大于雜交牛。該結論為進一步揭示FABP1和FABP2基因對脂肪酸的調控機制提供了理論依據。
4 結論
FABP1和FABP2基因在關嶺牛中保守性較高,且在不同年齡階段及不同組織中均有表達,尤其在肝臟和小腸中的表達水平較高,故推測FABP1和FABP2基因協同調控關嶺牛脂肪酸的合成代謝。
參考文獻:
姜延志,李學偉. 2006. 豬I-FABP基因的分子克隆與組織特異性表達分析(英文)[J]. 遺傳學報,33(2):125-132. [Jiang Y Z,Li X W. 2006. Molecular cloning and tissue-specific expression of intestinal-type fatty acid binding protein in porcine[J]. Acta Genetica Sinica,33(2):125-132.] doi:10.1016/S0379-4172(06)60031-0.
廖秀冬,任立明,王光瑛,李昂,黃銀花. 2012. 北京鴨FABP2基因多態性與體尺和屠體性狀的相關性研究[J]. 中國家禽,34(17):23-26. [Liao X D,Ren L M,Wang G Y,Li A,Huang Y H. 2012. Polymorphism of FABP2 gene and its association with body size and carcass traits in Peking duck[J].China Poultry,34(17):23-26.] doi:10. 16372/j.issn.1004-6364.2012.17.006.
王啟貴. 2004. 雞FABP基因克隆、表達特性及功能研究[D]. 哈爾濱:東北農業大學. [Wang Q G. 2004. Cloning,expression and function of chicken FABP genes[D]. Harbin:Northeast Agricultural University.] doi:10.7666/d.y631339.
趙采芹,王燕碧,張福平,唐宏,周磊,韓一帆,段志強. 2022. 雞TRAF6和TIFA基因克隆及其組織表達特征分析[J]. 南方農業學報,53(2):546-556. [Zhao C Q,Wang Y B,Zhang F P,Tang H,Zhou L,Han Y F,Duan Z Q. 2022. Cloning and analysis tissue expression of chicken TRAF6 and TIFA genes[J]. Journal of Southern Agriculture,53(2):546-556.] doi:10.3969/j.issn.2095-1191.2022.02.028.
Alpers D H,Bass N M,Engle M J,DeSchryver-Kecskemeti K. 2000. Intestinal fatty acid binding protein may favor differential apical fatty acid binding in the intestine[J]. Biochimica et Biophysica Acta,1483(3):352-362. doi:10.1016/s1388-1981(99)00200-0.
Atshaves B P,Martin G G,Hostetler H A,McIntosh A L,Kier A B,Schroeder F. 2010. Liver fatty acid-binding protein and obesity[J]. The Journal of Nutritional Biochemistry,21(11):1015-1032. doi:10.1016/j.jnutbio.2010.01.005.
Baier L J,Sacchettini J C,Knowler W C,Eads J,Paolisso G,Tataranni P A,Mochizuki H,Bennett P H,Bogardus C,Prochazka M. 1995. An amino acid substitution in the human intestinal fatty acid binding protein is associated with increased fatty acid binding, increased fat oxidation,and insulin resistance[J]. The Journal of Clinical Investigation,95(3):1281-1287. doi:10.1172/JCI117778.
Chan L,Wei C F,Li W H,Yang C Y,Ratner P,Pownall H,Gotto Jr A M,Smith L C. 1985. Human liver fatty acid binding protein cDNA and amino acid sequence. Functional and evolutionary implications[J]. The Journal of Biological Chemistry,260(5):2629-2632.
Chmurzyńska A. 2006. The multigene family of fatty acid-binding proteins (FABPs):Function,structure and polymorphism[J]. Journal of Applied Genetics,47(1):39-48. doi:10.1007/BF03194597.
Gajda A M,Storch J. 2015. Enterocyte fatty acid-binding proteins (FABPs):Different functions of liver and intestinal FABPs in the intestine[J]. Prostaglandins Leukotrienes & Essential Fatty Acids,93:9-16. doi:10.1016/j.plefa. 2014.10.001.
He J,Chen J C,Lu L Z,Tian Y,Tao Z G,Wang D Q,Li J J,Li G Q,Shen J D,Fu Y,Niu D. 2012. A novel SNP of liver-type fatty acid-binding protein gene in duck and its associations with the intramuscular fat[J]. Molecular Bio-logy Reports,39(2):1073-1077. doi:10.1007/s11033-011-0833-z.
He J,Tian Y,Li J J,Shen J D,Tao Z G,Fu Y,Niu D,Lu L Z. 2013. Expression pattern of L-FABP gene in different tissues and its regulation of fat metabolism-related genes in duck[J]. Molecular Biology Reports,40(1):189-195. doi:10.1007/s11033-012-2048-3.
Hunter J E. 2001. Studies on effects of dietary fatty acids as related to their position on triglycerides[J]. Lipids,36(7):655-668. doi:10.1007/s11745-001-0770-0.
Jensen G L,Mascioli E A,Seidner D L,Istfan N W,Domnitch A M,Selleck K,Babayan V K,Blackburn G L,Bistrian B R. 1990. Parenteral infusion of long-and medium-chain triglycerides and reticuloendothelial system function in man[J]. Journal of Parenteral and Enteral Nutrition,14(5):467-471. doi:10.1177/0148607190014005467.
Lackey A I,Chen T N,Zhou Y X,Bottasso Arias N M,Gajda A M,Joseph L B,Storch J. 2019. Altered small intestinal morphology and whole-body energy homeostasis in the intestinal fatty acid binding protein (IFABP) knockout mouse[J]. The FASEB Journal,33(S1):583.5. doi:10. 1096/fasebj.2019.33.1_supplement.583.5.
Martin G G,Danneberg H,Kumar L S,Atshaves B P,Erol E,Bader M,Schroeder F,Binas B. 2003. Decreased liver fatty acid binding capacity and altered liver lipid distribution in mice lacking the liver fatty acid-binding protein gene[J]. Journal of Biological Chemistry,278(24):21429-21438. doi:10.1074/jbc.M300287200.
Mukai T,Egawa M,Takeuchi T,Yamashita H,Kusudo T. 2017. Silencing of FABP1 ameliorates hepatic steatosis,inflammation,and oxidative stress in mice with nonalcoholic fatty liver disease[J]. FEBS OpenBio,7(7):1009-1016. doi:10.1002/2211-5463.12240.
Nakamura M T,Yudell B E,Loor J J. 2014. Regulation of energy metabolism by long-chain fatty acids[J]. Progress in Lipid Research,53:124-144. doi:10.1016/j.plipres.2013. 12.001.
Newberry E P,Kennedy S M,Xie Y,Luo J Y,Davidson N O. 2009. Diet-induced alterations in intestinal and extrahepatic lipid metabolism in liver fatty acid binding protein knockout mice[J]. Molecular and Cellular Biochemi-stry,326:79-86. doi:10.1007/s11010-008-0002-4.
Newberry E P,Xie Y,Kennedy S M,Han X L,Buhman K K,Luo J Y,Gross R W,Davidson N O. 2003. Decreased hepatic triglyceride accumulation and altered fatty acid uptake in mice with deletion of the liver fatty acid-binding protein gene[J]. Journal of Biological Chemistry,278(51):51664-51672. doi:10.1074/jbc.M309377200.
Ono T,Odani S. 2010. Initial studies of the cytoplasmic FABP superfamily[J]. Proceedings of the Japan Academy. Series B:Physical and Biological Sciences,86(3):220-228. doi:10.2183/pjab.86.220.
Schaap F G,van der Vusse G J,Glatz J F C. 2002. Evolution of the family of intracellular lipid binding proteins in vertebrates[J]. Molecular & Cellular Biochemistry,239(1-2):69-77. doi:10.1023/a:1020519011939.
Spann N J,Kang S,Li A C,Chen A Z,Newberry E P,Davidson N O,Hui S T Y,Davis R A. 2006. Coordinate transcriptional repression of liver fatty acid-binding protein and microsomal triglyceride transfer protein blocks hepa-tic very low density lipoprotein secretion without hepatosteatosis[J]. Journal of Biological Chemistry,281(44):33066-33077. doi:10.1074/jbc.M607148200.
Storch J,Thumser A E. 2010. Tissue-specific functions in the fatty acid-binding protein family[J]. Journal of Biological Chemistry,285(43):32679-32683. doi:10.1074/jbc.R110.135210.
Venkatachalam A B,Parmar M B,Wright J M. 2017. Evolution of the duplicated intracellular lipid-binding protein genes of teleost fishes[J]. Molecular Genetics and Geno-mics,292(4):699-727. doi:10.1007/s00438-017-1313-5.
Wang G Q,Bonkovsky H L,de Lemos A,Burczynski F J. 2015. Recent insights into the biological functions of liver fatty acid binding protein 1[J]. Journal of Lipid Research,56(12):2238-2247. doi:10.1194/jlr.R056705.
Wang G Q,Chen Q M,Minuk G Y,Gong Y W,Burczynski F J. 2004. Enhanced expression of cytosolic fatty acid bin-ding protein and fatty acid uptake during liver regeneration in rats[J]. Molecular and Cellular Biochemistry,262:41-49. doi:10.1023/b:mcbi.0000038214.52184.82.
Wang Y J,Tang K Q,Zhang W,Guo W L,Wang Y N,Zan L S,Yang W C. 2019. Fatty acid-binding protein 1 increa-ses steer fat deposition by facilitating the synthesis and secretion of triacylglycerol in liver[J]. PLoS One,14(4):e0214144. doi:10.1371/journal.pone.0214144.
Wu Y L,Peng X E,Zhu Y B,Yan X L,Chen W N,Lin X. 2016. Hepatitis B virus X protein induces hepatic steatosis by enhancing the expression of liver fatty acid bin-ding protein[J].Journal of Virology,90(4):1729-1740. doi:10.1128/JVI.02604-15.
Zimmerman A W,van Moerkerk H T,Veerkamp J H. 2001. Ligand specificity and conformational stability of human fatty acid-bindingproteins[J]. The International Journal of Biochemistry and Cell Biology,33(9):865-876. doi:10.1016/s1357-2725(01)00070-x.
(責任編輯 陳德元)
