水稻Osp39基因的組織表達(dá)和亞細(xì)胞定位分析
2023-07-08 01:36:12騰海艷
華北農(nóng)學(xué)報(bào) 2023年3期
騰海艷
(宜春學(xué)院 化學(xué)與生物工程學(xué)院,江西 宜春 336000)
P39蛋白又稱SP2(Suppressor of ppi1 locus 2)蛋白[1]或OEP80(Outer envelope protein 80),屬于OMP85(Outer membrane protein of 85 ku)蛋白家族[2],該家族成員定位于葉綠體、線粒體膜或細(xì)菌細(xì)胞膜,參與蛋白質(zhì)跨膜轉(zhuǎn)運(yùn)或膜上定位[2-3]。大部分OMP85蛋白由C端β-桶狀跨膜結(jié)構(gòu)域和N端多肽轉(zhuǎn)運(yùn)結(jié)合(Polypeptide transport associated,POTRA)結(jié)構(gòu)域組成,β-桶狀跨膜結(jié)構(gòu)域是蛋白質(zhì)通道,POTRA結(jié)構(gòu)域的作用類似分子伴侶,能夠結(jié)合被轉(zhuǎn)運(yùn)的蛋白質(zhì)并將其傳遞到內(nèi)膜[4],但是P39蛋白及其同源蛋白P36缺少POTRA結(jié)構(gòu)域[3]。在擬南芥中,P39蛋白定位于葉綠體外膜[3],Hsueh等[5]發(fā)現(xiàn),P39蛋白影響成熟擬南芥的代謝穩(wěn)態(tài),p39基因突變后,擬南芥成熟植株的光合活性和類囊體結(jié)構(gòu)均發(fā)生改變,多條代謝途徑出現(xiàn)異常。根據(jù)Ling等[1]的報(bào)道,擬南芥通過SP1蛋白、P39蛋白和CDC48(Cell division cycle 48)蛋白協(xié)作,促進(jìn)葉綠體外膜轉(zhuǎn)運(yùn)蛋白的泛肽化和降解,從而控制外源蛋白質(zhì)向葉綠體的輸入,此過程稱為葉綠體相關(guān)蛋白降解(Chloroplast-associated protein degradation,CHLORAD)機(jī)制,該機(jī)制不僅參與葉綠體蛋白的輸入調(diào)控和氧化脅迫響應(yīng)[1],在番茄中,還參與番茄有色體的發(fā)育,從而影響果實(shí)的成熟過程[6]。
蛋白質(zhì)的輸入調(diào)控影響著質(zhì)體的功能和植物的光合效率,在農(nóng)作物中,葉綠體和有色體的蛋白質(zhì)輸入調(diào)控還將影響到作物的產(chǎn)量、抗逆性及農(nóng)產(chǎn)品品質(zhì)[6-8]。在前期工作中,已經(jīng)對(duì)CHLORAD機(jī)制中的sp1基因在水稻中的同源基因OsSP1(登錄號(hào):Os07g0647800)進(jìn)行了研究,并發(fā)現(xiàn)該基因的CRISPR/Cas9基因編輯水稻植株具有嚴(yán)重的生長(zhǎng)抑制表型,顯示了葉綠體蛋白輸入調(diào)控途徑對(duì)水稻生長(zhǎng)發(fā)育的重要影響[9-10]。為繼續(xù)探究水稻的CHLORAD機(jī)制,本研究選擇p39基因在水稻中的同源基因Osp39作為研究對(duì)象,通過生物信息學(xué)手段以及轉(zhuǎn)錄分析、原生質(zhì)體瞬時(shí)表達(dá)等方法,確定該基因及其編碼蛋白的結(jié)構(gòu)和性質(zhì)特征、組織表達(dá)情況及亞細(xì)胞定位情況,為深入研究Osp39基因的功能,進(jìn)而為闡明水稻葉綠體蛋白質(zhì)輸入調(diào)控機(jī)制提供基礎(chǔ)。
1 材料和方法
1.1 試驗(yàn)材料及生長(zhǎng)條件
本研究使用的水稻(OryzasativaL.)為粳稻品種中花11,亞細(xì)胞定位、組織表達(dá)使用的土培和水培試驗(yàn)材料均種植于宜春學(xué)院苗圃,4—7月份自然溫度和光照條件下生長(zhǎng),水培材料每3 d更換1次木村B營(yíng)養(yǎng)液,常規(guī)管理。
1.2 主要試劑
RNA提取試劑盒和DNA回收試劑盒購(gòu)自Magen公司,反轉(zhuǎn)錄試劑和DNA聚合酶購(gòu)自諾唯贊公司,2×SYBR mix、EcoR Ⅰ和HindⅢ限制性內(nèi)切酶、T4DNA連接酶均購(gòu)自TaKaRa公司,原生質(zhì)體制備使用的纖維素酶和離析酶購(gòu)自Yakult公司,其他木村B營(yíng)養(yǎng)液配制、凝膠電泳、原生質(zhì)體制備、轉(zhuǎn)化等試劑均為國(guó)產(chǎn)分析純。
1.3 Osp39基因及其蛋白產(chǎn)物的生物信息學(xué)分析
基于擬南芥p39基因的mRNA序列及AtP39蛋白的氨基酸序列,在NCBI(https://www.ncbi.nlm.nih.gov/)網(wǎng)站通過Blast工具搜索水稻中的同源基因Osp39的mRNA序列和完整基因序列,以及水稻、玉米、大豆等共12 種植物的P39蛋白質(zhì)序列。PLACE數(shù)據(jù)庫(kù)和PlantCARE數(shù)據(jù)庫(kù)進(jìn)行啟動(dòng)子順式作用元件分析,DNAMAN軟件進(jìn)行蛋白質(zhì)氨基酸序列同源性分析,MEGA軟件構(gòu)建系統(tǒng)進(jìn)化樹,Expasy-ProtParam數(shù)據(jù)庫(kù)進(jìn)行蛋白質(zhì)理化性質(zhì)分析,SignalP數(shù)據(jù)庫(kù)進(jìn)行蛋白質(zhì)信號(hào)肽預(yù)測(cè),TMHMM數(shù)據(jù)庫(kù)進(jìn)行蛋白質(zhì)跨膜區(qū)位置預(yù)測(cè),SOPMA數(shù)據(jù)庫(kù)進(jìn)行二級(jí)結(jié)構(gòu)元件分析,SWISS-MODEL數(shù)據(jù)庫(kù)進(jìn)行蛋白質(zhì)空間結(jié)構(gòu)預(yù)測(cè),STRING數(shù)據(jù)庫(kù)進(jìn)行蛋白質(zhì)互作分析,PredictProtein數(shù)據(jù)庫(kù)進(jìn)行亞細(xì)胞定位預(yù)測(cè)。
1.4 RNA提取和定量PCR分析
苗期水稻樣品取自木村B營(yíng)養(yǎng)液培養(yǎng)14 d的水稻苗,分別取根(葉鞘下約5 cm長(zhǎng))、葉鞘和由上至下第2 片葉片,揚(yáng)花期水稻取自土培的揚(yáng)花期水稻植株,分別取旗葉中段、莖和花(包括雌蕊、雄蕊和穎殼),每種樣品均平行取樣3 份,混合,快速放入液氮中,經(jīng)試劑盒提取總RNA,反轉(zhuǎn)錄獲得cDNA后,以cDNA作為模板,進(jìn)行Osp39基因和內(nèi)參基因OsActin1(登錄號(hào):Os03g07181000)的定量PCR擴(kuò)增。定量PCR反應(yīng)程序:95 ℃,3 min;95 ℃,15 s,60 ℃,30 s,40次循環(huán)。定量PCR反應(yīng)體系:2×SYBR mix 5.0 μL;F、R引物(10 μmol/L)各0.5 μL;cDNA 1.0 μL;雙蒸水補(bǔ)至10.0 μL。PCR結(jié)果用2-ΔΔCt法[11]進(jìn)行計(jì)算,SPSS軟件進(jìn)行顯著性分析。定量PCR引物序列見表1。
1.5 Osp39-gfp瞬時(shí)表達(dá)載體的構(gòu)建
以1.4中獲得的cDNA作為模板,擴(kuò)增不含終止密碼子的Osp39編碼序列,PCR產(chǎn)物經(jīng)凝膠電泳檢測(cè),DNA試劑盒回收后,采用酶切、連接法將酶切產(chǎn)物克隆到pOX載體的gfp序列上游的多克隆位點(diǎn),構(gòu)建OsP39蛋白與GFP蛋白的融合表達(dá)載體并轉(zhuǎn)化大腸桿菌TOP10菌株。PCR擴(kuò)增引物見表1。
1.6 亞細(xì)胞定位分析
一葉期土培水稻幼苗,取約5 cm長(zhǎng)的葉鞘部分制備水稻原生質(zhì)體[12],將OsP39蛋白與GFP蛋白的融合表達(dá)載體質(zhì)粒轉(zhuǎn)入原生質(zhì)體,同時(shí)轉(zhuǎn)化GFP蛋白的載體質(zhì)粒作為對(duì)照,室溫避光孵育過夜后,激光共聚焦顯微鏡(Carl Zeiss,LSM7 DUO)觀察綠色熒光蛋白GFP在細(xì)胞內(nèi)的定位情況。
2 結(jié)果與分析
2.1 Osp39基因的結(jié)構(gòu)
在NCBI數(shù)據(jù)庫(kù)通過Blast功能獲得水稻Osp39基因序列,結(jié)果顯示,Osp39基因位于水稻5號(hào)染色體,基因登錄號(hào)為Os05g0510200,基因編碼區(qū)共有11個(gè)短外顯子,L6 loop區(qū)保守序列的編碼區(qū)位于第9外顯子上,轉(zhuǎn)錄產(chǎn)物無可變剪切方式(圖1),成熟mRNA的編碼區(qū)長(zhǎng)1 086 bp,使用該編碼區(qū)序列進(jìn)行Blast,在水稻基因組中未找到其他功能相關(guān)的同源序列。
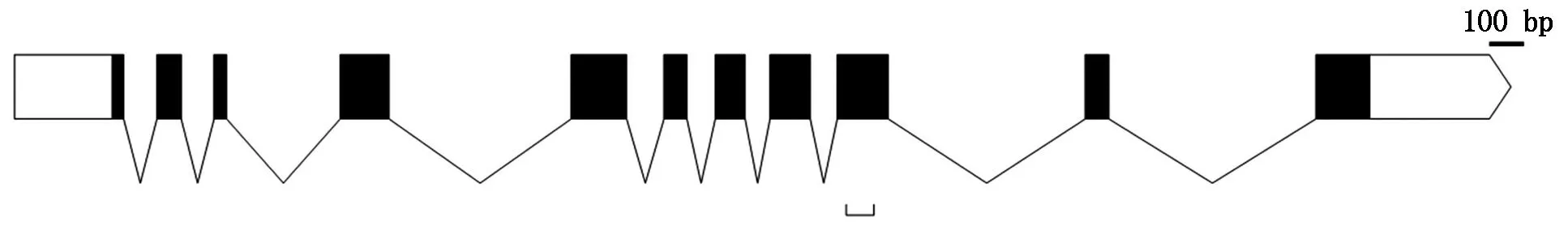
下括號(hào)位置為 L6 loop區(qū)編碼位點(diǎn)。
2.2 Osp39啟動(dòng)子的上游元件
從Osp39基因起始密碼子ATG前面截取長(zhǎng)度為2 200 bp的上游啟動(dòng)子序列,使用PLACE數(shù)據(jù)庫(kù)和PlantCARE數(shù)據(jù)庫(kù)進(jìn)行啟動(dòng)子順式作用元件分析,結(jié)果顯示,Osp39啟動(dòng)子中含有I box、ATCT-motif、CPBCSPOR等多個(gè)光響應(yīng)元件和葉綠體調(diào)控位點(diǎn),此外還有TGACG-motif等多個(gè)激素響應(yīng)元件,多個(gè)MYB轉(zhuǎn)錄因子識(shí)別和結(jié)合位點(diǎn)及ARE、LTR等厭氧和低溫響應(yīng)元件(表2),這些上游元件調(diào)節(jié)不同生長(zhǎng)狀態(tài)和環(huán)境條件下基因的轉(zhuǎn)錄水平,是基因表達(dá)調(diào)控的重要元件。
2.3 P39蛋白的同源進(jìn)化分析
目前,有研究報(bào)道的P39蛋白僅擬南芥AtP39一種[1-3,5-8],本研究選擇了水稻、玉米、大豆等13 種常見的單、雙子葉農(nóng)作物和經(jīng)濟(jì)作物,通過Blast搜索獲得這些植物的P39同源蛋白序列(蛋白質(zhì)登錄號(hào)見圖2)。采用DNAMAN進(jìn)行序列比對(duì),結(jié)果顯示,13 種植物的P39蛋白均與AtP39蛋白具有較高的序列相似性(圖3)。此外,AtP39蛋白含有一個(gè)高度保守的由27個(gè)氨基酸殘基組成的L6 loop區(qū),位于第245—271 氨基酸殘基處,其功能可能是參與蛋白質(zhì)構(gòu)象轉(zhuǎn)變的調(diào)節(jié)[3],參與比對(duì)的12種P39蛋白也均在相同位置存在L6 loop區(qū),其中,水稻OsP39的L6 loop區(qū)與AtP39僅相差一個(gè)氨基酸殘基(圖3)。采用MEGA軟件,選擇最大可能性法[13],對(duì)P39蛋白進(jìn)行序列比對(duì)和進(jìn)化樹構(gòu)建,結(jié)果顯示,水稻OsP39蛋白序列與高粱SbP39的遺傳距離最近,為0.086 8(圖2)。
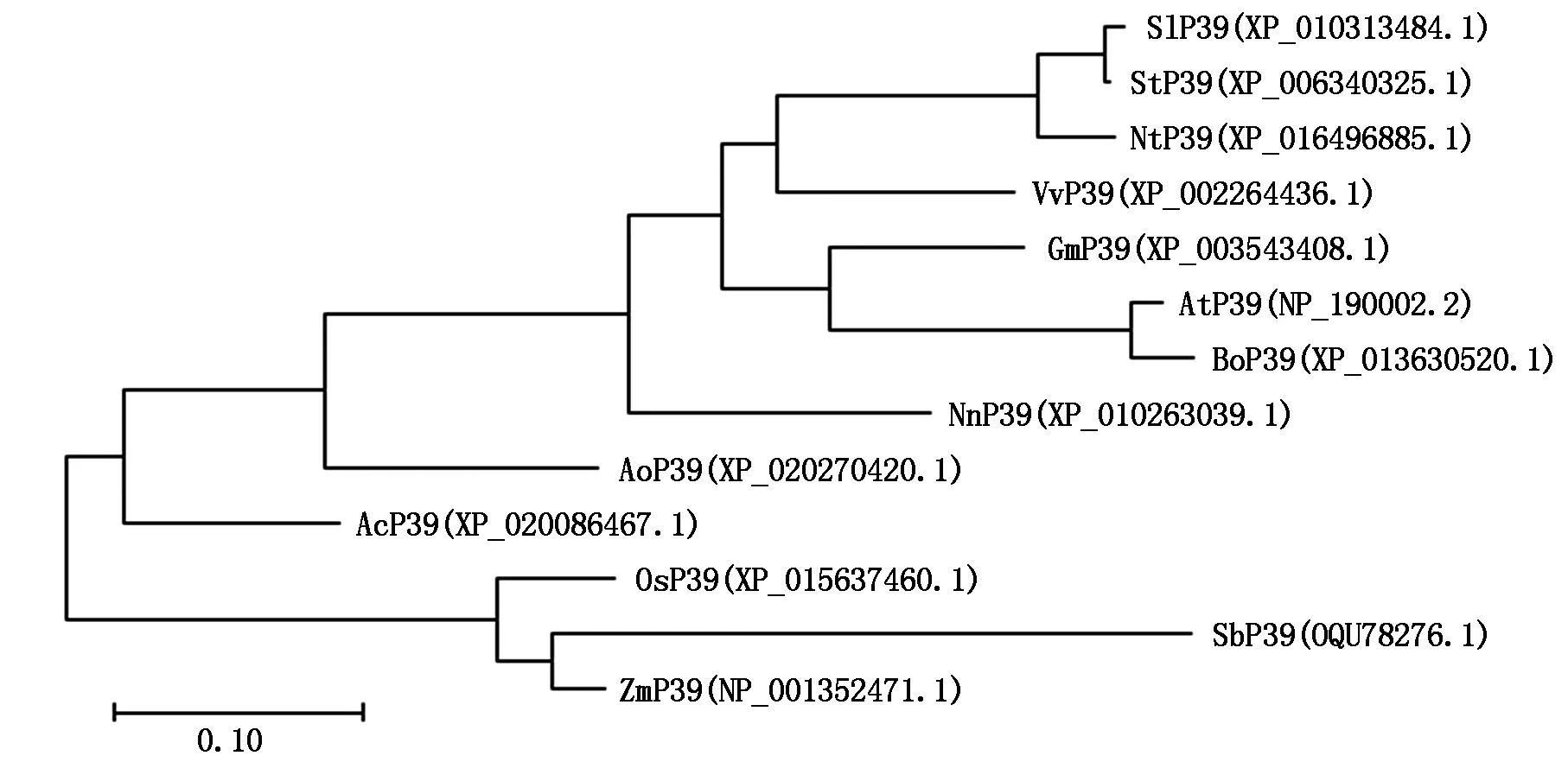
At.擬南芥;Os.水稻;Sb.高粱;Zm.玉米;Ac.菠蘿;Ao.蘆筍;Bo.甘藍(lán);Gm.大豆;Nn.荷花;Nt.煙草;Sl.番茄;St.馬鈴薯;Vv.葡萄。圖3同。At.Arabidopsis thalianan;Os.Oryza sativa;Sb.Sorghum bicolor;Zm.Zea mays;Ac.Ananas comosus;Ao.Asparagus officinalis;Bo.Brassica oleracea var.Oleracea;Gm.Glycine max;Nn.Nelumbo nucifera;Nt.Nicotiana tabacum;Sl.Solanum lycopersicum;St.Solanum tuberosum;Vv.Vitis vinifera.The same as Fig.3.
2.4 OsP39蛋白的理化性質(zhì)
利用Expasy-ProtParam數(shù)據(jù)庫(kù)進(jìn)行蛋白質(zhì)性質(zhì)分析,結(jié)果顯示,OsP39蛋白含361 個(gè)氨基酸殘基,包含了全部20 種蛋白質(zhì)氨基酸,其中甘氨酸含量最高,為44 個(gè),占總數(shù)12.2%,其次為亮氨酸和絲氨酸,分別為33,32 個(gè),色氨酸數(shù)量最少,為2 個(gè),生理?xiàng)l件下帶負(fù)電荷氨基酸(谷氨酸+天冬氨酸)總數(shù)29 個(gè),帶正電荷氨基酸(賴氨酸+精氨酸)總數(shù)33 個(gè)。蛋白質(zhì)分子量38.7 ku,理論等電點(diǎn)pH值8.64,不穩(wěn)定指數(shù)32.6,小于40,穩(wěn)定性較好[14],脂肪指數(shù)83.71,熱穩(wěn)定性高,屬于耐熱型蛋白[15],平均疏水性-0.103,偏向親水。
2.5 OsP39蛋白信號(hào)肽和跨膜螺旋區(qū)預(yù)測(cè)
通過SignalP進(jìn)行信號(hào)肽分析,在OsP39蛋白中未識(shí)別到氨基端信號(hào)肽序列(圖4),利用TMHMM數(shù)據(jù)庫(kù)分析蛋白質(zhì)的跨膜螺旋區(qū),未預(yù)測(cè)到跨膜螺旋結(jié)構(gòu)(圖5)。
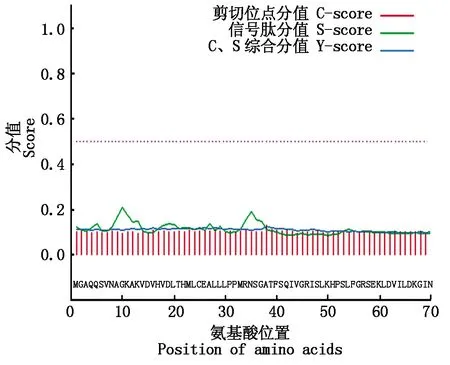
圖4 OsP39蛋白的信號(hào)肽分析Fig.4 Analysis of signal peptide of OsP39 protein
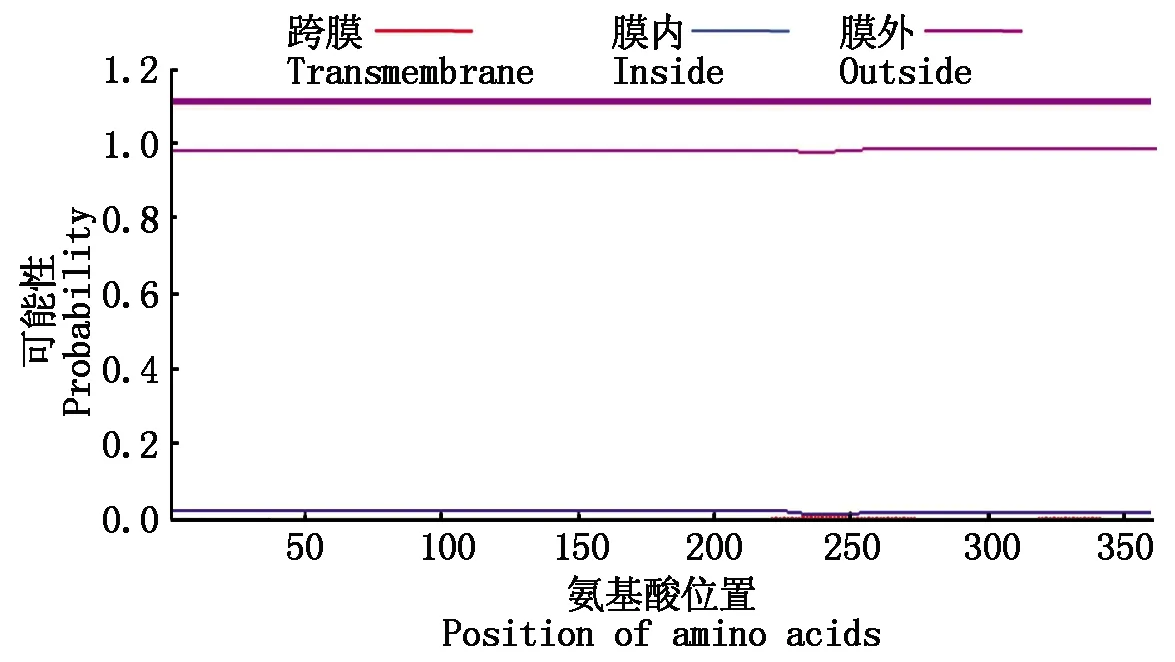
圖5 OsP39蛋白的跨膜螺旋區(qū)分析Fig.5 Analysis of transmembrane helical region of OsP39 protein
2.6 OsP39蛋白的空間結(jié)構(gòu)
二級(jí)結(jié)構(gòu)預(yù)測(cè)顯示,OsP39蛋白的二級(jí)結(jié)構(gòu)元件以無規(guī)卷曲和伸展肽鏈(β折疊片)為主,其中無規(guī)卷曲比例最高,占50.97%,其次為伸展肽鏈,占30.57%,α螺旋和β轉(zhuǎn)角分別占12.74%,5.54%,不含310螺旋、π螺旋等元件(圖6)。
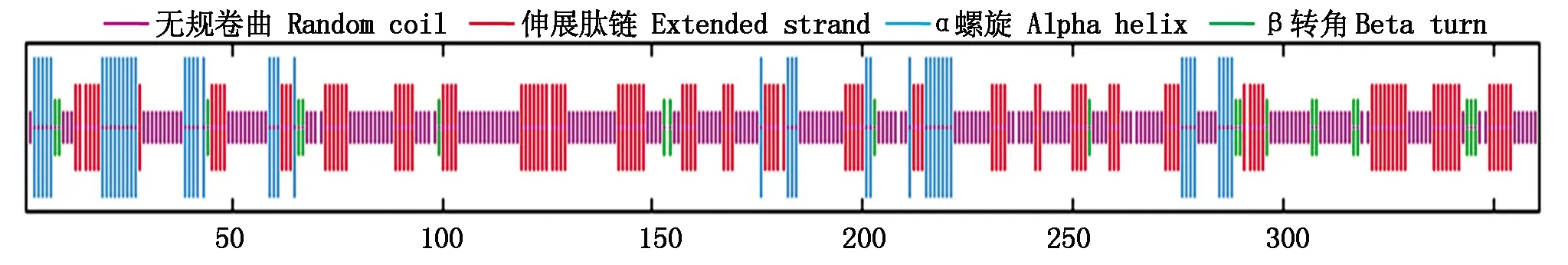
圖6 OsP39蛋白的二級(jí)結(jié)構(gòu)元件Fig.6 Secondary structure elements of OsP39 protein
通過SWISS-MODEL數(shù)據(jù)庫(kù)進(jìn)行三級(jí)結(jié)構(gòu)預(yù)測(cè)顯示,OsP39的空間結(jié)構(gòu)與擬南芥AtP39蛋白幾乎相同,都為含多個(gè)β折疊片的β-桶狀膜蛋白(圖7),桶狀結(jié)構(gòu)主體部分插入膜內(nèi),L6 loop區(qū)形成松散的環(huán)狀結(jié)構(gòu),位于通道入口處(圖7)。

黑色箭頭指示L6 loop區(qū)。Black arrows indicate the L6 loop region.
2.7 OsP39蛋白的蛋白質(zhì)互作和亞細(xì)胞定位預(yù)測(cè)
通過STRING數(shù)據(jù)庫(kù)進(jìn)行蛋白質(zhì)互作分析顯示,目前有證據(jù)支持的與OsP39蛋白(轉(zhuǎn)錄產(chǎn)物ID:OS05T0510200-01)存在互作關(guān)系的水稻基因有10 個(gè)(圖8),其中可信度最高的是谷胱甘肽硫轉(zhuǎn)移酶(OS06T0168000-01),其次為OsDVB1蛋白(OS03T0840900-01),該蛋白與植物發(fā)育有關(guān),以及OS06T0111600-01(含有依賴AMP的合成酶/連接酶結(jié)構(gòu)域的蛋白質(zhì)),其他可能與OsP39存在互作關(guān)系的蛋白質(zhì)分別為OsJ_04947(ABC轉(zhuǎn)運(yùn)蛋白亞基)、OS12T0422971-00(NADH脫氫酶亞基9類似蛋白)、OsJ_20876(果糖/塔格糖二磷酸醛縮酶)、OsJ_10671(類ISP42蛋白)、OsJ_05254(未知功能蛋白)、OS02T0496900-01(線粒體輸入受體TOM9-2亞基)、OS01T0276200-00(線粒體輸入受體亞基TOM40類似蛋白)。上述可能的互作蛋白在水稻中多數(shù)未被深入研究,其中的類ISP42蛋白、OS02T0496900-01等蛋白與線粒體蛋白質(zhì)輸入有關(guān)。通過PredictProtein數(shù)據(jù)庫(kù)進(jìn)行亞細(xì)胞定位預(yù)測(cè)則顯示,OsP39蛋白定位于葉綠體膜(圖9)。
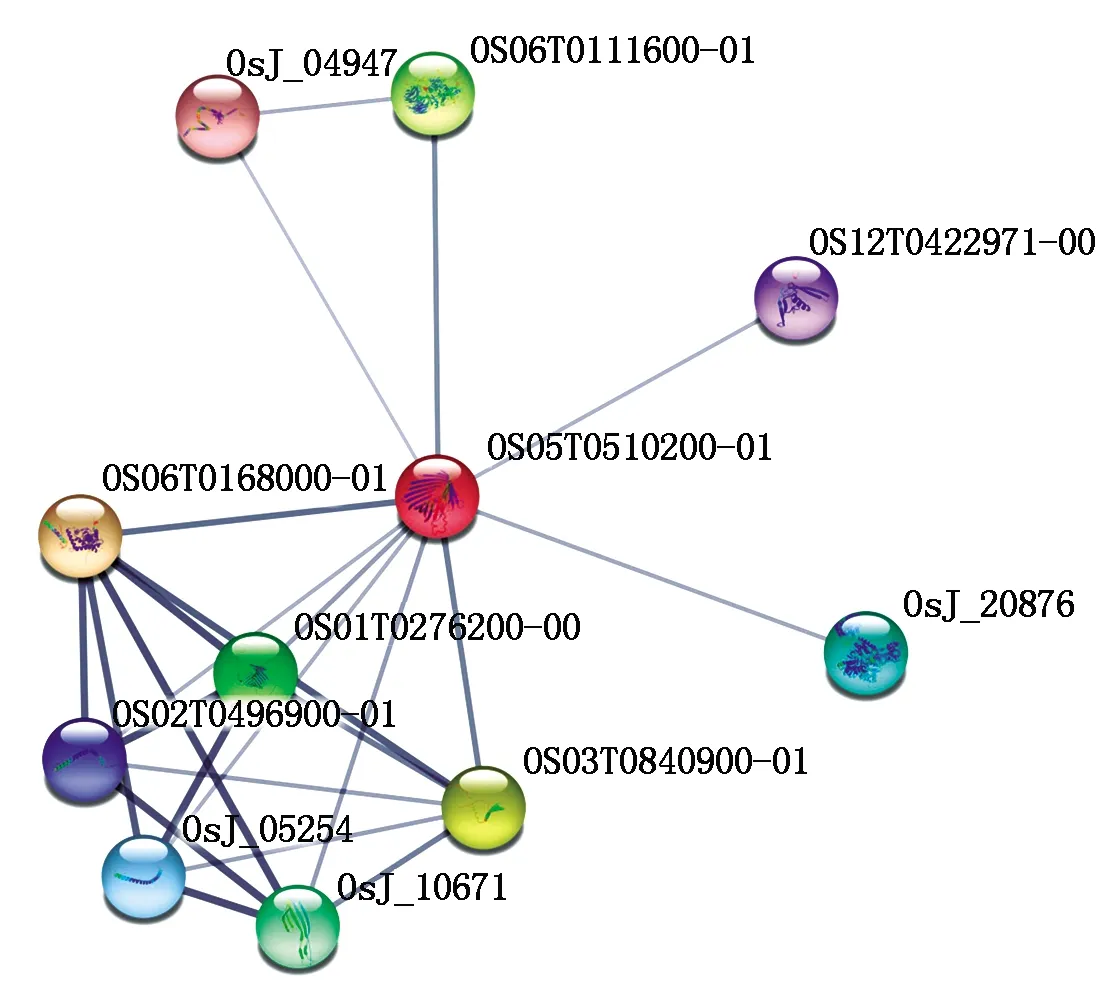
直線的粗細(xì)程度表示可信程度。The thickness of the straight line indicates the degree of confidence.
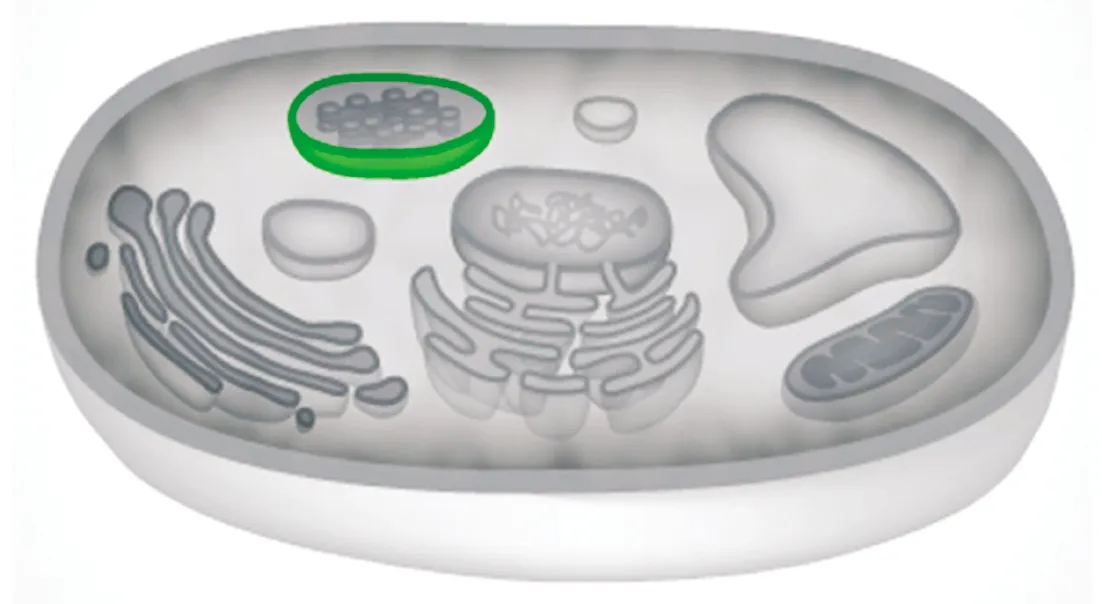
圖9 OsP39蛋白的亞細(xì)胞定位預(yù)測(cè)Fig.9 Prediction of subcellular localization of OsP39 protein
2.8 OsP39蛋白的亞細(xì)胞定位
盡管通過SignalP未預(yù)測(cè)到信號(hào)肽序列,但空間結(jié)構(gòu)預(yù)測(cè)結(jié)果顯示,OsP39蛋白為膜蛋白,亞細(xì)胞定位預(yù)測(cè)也顯示,該蛋白為葉綠體膜定位蛋白。為確定OsP39蛋白在水稻細(xì)胞內(nèi)的定位情況,以綠色熒光蛋白GFP作為報(bào)告蛋白,使用酶切、連接法構(gòu)建了Osp39編碼序列與gfp序列融合的瞬時(shí)表達(dá)載體。用帶有Osp39-gfp融合表達(dá)框的質(zhì)粒轉(zhuǎn)化原生質(zhì)體后,發(fā)現(xiàn)OsP39-GFP融合蛋白主要出現(xiàn)在葉綠體區(qū)域,且GFP熒光與葉綠素自發(fā)熒光疊加后并未混合為黃光,因此,可以確定OsP39-GFP融合蛋白沒有出現(xiàn)在葉綠體基質(zhì),而是定位在葉綠體膜(圖10),表明OsP39是與水稻葉綠體膜功能相關(guān)的蛋白質(zhì)。另外,少量OsP39-GFP融合蛋白積累在細(xì)胞質(zhì),推測(cè)為蛋白質(zhì)表達(dá)過量導(dǎo)致。作為對(duì)照的gfp質(zhì)粒轉(zhuǎn)化原生質(zhì)體后,GFP蛋白則全部定位于細(xì)胞質(zhì)(圖10)。
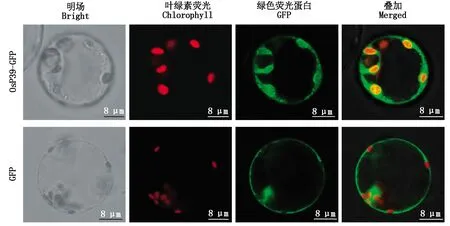
圖10 OsP39-GFP融合蛋白和GFP在水稻原生質(zhì)體中的定位Fig.10 Subcellular localization of OsP39-GFP fusion protein and GFP in rice protoplasts
2.9 Osp39基因的組織表達(dá)
為明確Osp39基因在水稻中的組織表達(dá)情況,采用定量PCR進(jìn)行了基因在不同時(shí)期和不同組織中的轉(zhuǎn)錄水平分析。結(jié)果顯示,在苗期,Osp39在水稻葉片中表達(dá)水平最高,其次為葉鞘,而在根中最低。在開花期,Osp39在水稻旗葉中表達(dá)水平最高,其次為莖,在花(包括雄蕊、雌蕊和穎殼)中表達(dá)量最低。在不同時(shí)期和不同組織中,Osp39基因的表達(dá)水平不同,但在葉和葉鞘等綠色組織中的表達(dá)水平極顯著高于根和花等組織(圖11)。
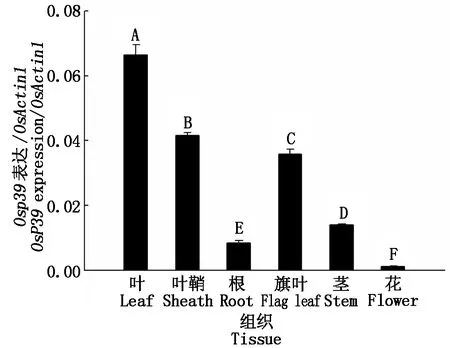
葉、葉鞘和根取自14 d水稻苗;旗葉、莖和花取自揚(yáng)花期水稻。不同的大寫字母表示差異的極顯著性(P<0.01)。Leaves,leaf sheaths and roots were taken from 14-day rice seedlings;flag leaves,stems and flowers were taken from flowering rice.Different capital letters indicate extremely significant differences(P<0.01).
3 結(jié)論與討論
葉綠體中含有約3 000種蛋白質(zhì),其中絕大多數(shù)由核基因組編碼,在細(xì)胞質(zhì)中翻譯后,輸入到葉綠體[16]。擬南芥通過CHLORAD機(jī)制調(diào)節(jié)葉綠體等質(zhì)體蛋白質(zhì)的輸入,AtP39蛋白是該機(jī)制中的一個(gè)重要作用因子,參與泛肽化蛋白由葉綠體外膜轉(zhuǎn)運(yùn)回細(xì)胞質(zhì)的過程[1]。本研究采用生物信息學(xué)手段及轉(zhuǎn)錄分析、瞬時(shí)表達(dá)等方法,確定了水稻Osp39基因的序列特征、組織表達(dá)情況以及OsP39蛋白的性質(zhì)、結(jié)構(gòu)和亞細(xì)胞定位情況,確定OsP39與擬南芥AtP39蛋白具有高度相似性。
由于目前尚未在β-桶狀膜蛋白中發(fā)現(xiàn)規(guī)律性的信號(hào)肽序列[17-18],因此,本研究利用SignalP數(shù)據(jù)庫(kù)對(duì)OsP39蛋白進(jìn)行信號(hào)肽分析時(shí),未能預(yù)測(cè)到信號(hào)肽序列。Gross等[19]研究表明,β-桶狀膜蛋白的定位信息存在于蛋白質(zhì)N端的6個(gè)β-折疊片中,定位過程還涉及葉綠體外膜TOC蛋白等蛋白質(zhì)的參與,其詳細(xì)定位機(jī)制尚不清楚。但此類蛋白確實(shí)可以進(jìn)行正確的膜定位,擬南芥AtP39蛋白的葉綠體膜定位[3],以及本研究中OsP39蛋白的亞細(xì)胞定位結(jié)果,均反映了這一事實(shí),也證實(shí)了OsP39是一種在葉綠體膜上發(fā)揮功能的蛋白質(zhì),與Osp39基因的啟動(dòng)子上游元件分析結(jié)果和組織表達(dá)結(jié)果相符合。
Osp39基因啟動(dòng)子中含有多個(gè)光響應(yīng)、葉綠體調(diào)節(jié)和激素響應(yīng)順式作用元件,組織表達(dá)分析也顯示,Osp39基因的表達(dá)具有明顯的組織特異性,在苗期和開花期,其在葉中均具有最高表達(dá)水平,而在非綠色組織根和花中的表達(dá)水平最低,這種表達(dá)模式與P39蛋白參與葉綠體調(diào)節(jié)的功能相對(duì)應(yīng)。在擬南芥中已經(jīng)確定AtP39蛋白與SP1蛋白和CDC48蛋白均具有互作關(guān)系[1],但由于水稻OsP39蛋白和OsSP1蛋白的研究報(bào)道較少,缺乏文獻(xiàn)等支持,因此,在蛋白質(zhì)互作預(yù)測(cè)中未能預(yù)測(cè)到OsP39與水稻OsSP1蛋白和OsCDC48蛋白的互作關(guān)系。本研究的組織表達(dá)結(jié)果顯示,Osp39基因在葉片、葉鞘和根中的表達(dá)水平和變化趨勢(shì)均與水稻OsSP1基因接近[9],并且OsSP1蛋白也定位于葉綠體[9],反映了二者在功能上的關(guān)聯(lián)性。
本研究在前期工作中發(fā)現(xiàn),水稻的OsSP1基因編輯植株具有嚴(yán)重的生長(zhǎng)發(fā)育障礙,且結(jié)實(shí)率極低[10],Huang等[20-21]的研究顯示,Oscdc48基因突變后水稻也出現(xiàn)早衰、葉片黃化、褐斑、結(jié)實(shí)率下降、早亡等表型,表明參與CHLORAD機(jī)制的基因?qū)λ镜纳L(zhǎng)發(fā)育具有重要調(diào)控作用,Osp39基因在水稻生長(zhǎng)發(fā)育、脅迫響應(yīng)過程中的作用有待于進(jìn)一步研究,本研究分析結(jié)果為Osp39基因功能的深入研究提供了基礎(chǔ),也為水稻葉綠體蛋白的輸入調(diào)控研究提供了新的線索。
猜你喜歡
青少年科技博覽(中學(xué)版)(2022年6期)2022-12-27 19:44:27
軍事文摘(2021年22期)2021-11-26 00:43:51
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
電子制作(2018年18期)2018-11-14 01:48:24
財(cái)經(jīng)(2017年2期)2017-03-10 14:35:35
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
財(cái)經(jīng)(2016年15期)2016-06-03 07:38:02
財(cái)經(jīng)(2016年3期)2016-03-07 07:44:46
