基于轉錄組分析的紫花苜蓿組成型抗薊馬性研究
2023-06-30 12:00:49爽爽代蕊陳崎劉慧潔姜曉紅特木爾布和譚瑤張志強
江蘇農業科學 2023年5期
爽爽 代蕊 陳崎 劉慧潔 姜曉紅 特木爾布和 譚瑤 張志強

摘要:為了探究紫花苜蓿組成型抗薊馬轉錄調控機制,以自主培育的抗薊馬苜蓿品種草原4號為研究對象,以感薊馬苜蓿品種草原2號為對照,采用RNA-seq技術進行轉錄組測序分析。試驗共獲得82.34 Gb clean data,組裝得到77 837個unigenes,139 949個轉錄本,平均序列長度為830.17 bp;共有47 573個unigenes被注釋到六大數據庫中。與草原2號相比,草原4號共檢測到差異基因2 388個,其中上調基因1 361個,下調基因1 027個。KEGG對相關差異基因的功能注釋表明,上調基因主要注釋到能量代謝、碳水化合物代謝等代謝過程以及遺傳信息處理相關途徑,下調基因主要注釋到遺傳信息處理、細胞過程和環境適應相關途徑。qRT-PCR驗證結果發現所選的8個基因在2個品種中的相對表達量同RNA-Seq測的結果一致,說明本試驗中轉錄組的結果可靠。結果可為揭示苜蓿組成型抗薊馬機理奠定基礎,進而為抗薊馬新品種培育及薊馬防治提供理論支撐。
關鍵詞:苜蓿;抗薊馬;轉錄組;RNA-seq
中圖分類號:S476;S433.89文獻標志碼:A
文章編號:1002-1302(2023)05-0044-07
紫花苜蓿(Medicago sativa L.)被稱為“牧草之王”,是一種高產、優質且固氮能力強的優良牧草,是中國及世界栽培最廣泛的牧草。但在紫花苜蓿的栽培生產過程中,蟲害是影響紫花苜蓿產量和品質的重要因素。我國已報道的苜蓿害蟲有8目48科297種,而薊馬(Thripidae)已成為苜蓿生產的主要害蟲之一[1-2]。薊馬是纓翅目昆蟲的統稱,是一種危險性入侵害蟲,寄主范圍較廣,其特有的銼吸式口器既可以通過挫破植物表皮,消耗韌皮部組織的汁液造成直接的攝食損害,也可以通過傳播番茄斑萎病毒在內的多種病毒造成間接的損害,從而導致作物、蔬菜及牧草的產量和質量下降[3-4]。據報道,我國在苜蓿生產中,薊馬每年造成10%~30%的草產量損失[5]。目前,薊馬的防治管理主要依靠化學農藥,但過量使用化學農藥會導致薊馬抗藥性、生態系統破壞和食品安全等問題[6-7]。因此,從長遠和經濟環保的角度出發,通過培育抗薊馬苜蓿新品種減少薊馬危害具有重要的經濟價值和生態效益[8-9]。從不同角度對植物抗薊馬機理的揭示是抗薊馬新品種培育及薊馬防治的理論支撐。
在與昆蟲的長期進化過程中,植物已經進化出包括組成型防御和誘導型防御在內的復雜的防御機制來抵御草食性害蟲的侵害。組成型防御是植物所固有的,不需要昆蟲取食誘導就能產生的防御策略,一般包括物理防御和化學防御。通常,植物表面毛狀體、蠟質角質層、硅和細胞壁等物理結構是阻止昆蟲取食的第一道防線[10-12],而包括毒素或代謝物在內的生化防御則是第二道防線[13-14]。近年來,從表型特征、生化防御、代謝調控、多組學及分子機制等不同層面有關不同植物抗薊馬機制的研究也成為熱點[4,8-9,15-16]。
在廣泛收集抗源的基礎上,筆者所在課題組采用輪回選擇法,成功選育出草原4號紫花苜蓿——抗薊馬苜蓿新品種[10]。以感薊馬苜蓿品種草原2號為對照,筆者所在課題組已經從表型特征、生化防御、激素等角度開展苜蓿抗薊馬機理的探索[16-20],但缺乏從分子層面對草原4號組成型抗薊馬機制的分析。因此,本研究以草原4號和草原2號苜蓿為材料,利用轉錄組測序技術,探索草原4號組成型抗薊馬的相關分子調控途徑,為苜蓿組成型抗蟲機制的深入挖掘及通過生物技術手段進行苜蓿新品種培育提供理論支撐。
1 材料與方法
1.1 供試材料及生長條件
以草原4號苜蓿(Medicago sativa L.‘Caoyuan No.4)和草原2號苜蓿(Medicago varia Martin.‘Caoyuan No.2)為試驗材料,2個品種皆為課題組培育的國家審定品種。依據蟲情指數低于0.5表示品種為抗蟲品種的評價標準[21],前期的抗薊馬評價證明,草原4號苜蓿的蟲情指數為0.334,為抗薊馬品種;草原2號苜蓿的蟲情指數為0.901,為感薊馬品種[19]。于2019年在內蒙古農業大學草原與資源環境學院人工氣候室內種植草原2號和草原4號苜蓿各30盆,每盆3株,其生長條件為晝夜平均溫度為(30±5) ℃和(20±5) ℃,相對濕度為(65±5)%和(70±5)%,保證所有材料生長環境一致,培育3周后,取頂部3~4張葉子于液氮中速凍,-80 ℃ 保存備用,每個品種3個重復。
1.2 試驗方法
1.2.1 RNA 測序
將低溫保存的葉片材料送上海美吉生物醫藥科技有限公司進行RNA-Seq測序分析,所用測序平臺為Illumina Hiseq 4000(版本2×150 bp)。測序原始序列數據保存在NCBI存檔(http://www.ncbi.nlm.nih.gov/Traces/sra,登錄號 PRJNA622603)。
1.2.2 RNA-Seq數據分析
使用Trinity (https://github.com/trinityrnaseq/trinityrnaseq)在線工具對所有樣本的clean data進行De novo組裝,獲得unigenes,并對其進行優化評估;將所有基因和轉錄本分別與 NR、Swiss-Prot、Pfam、COG、GO 和 KEGG 等六大數據庫進行比對從而獲取注釋信息,并統計各數據庫注釋情況;通過與NR(NCBI非冗余蛋白庫)綜合數據庫比對,查看本物種轉錄本序列與相近物種的相似情況;通過主成分分析(PCA)深入挖掘樣品之間的關系和變異大小。使用DESeq2軟件對測序數據進行差異基因的篩選,篩選參數為adjust P-value<0.05 & |log2FC| ≥1;并對差異基因進行KEGG功能注釋與富集分析[16,22]。
1.2.3 qRT-PCR 驗證
從不同處理的葉子中提取總RNA,使用帶有gDNA Eraser的 PrimeScript RT試劑進行cDNA反轉錄。使用ABI7500FAST 實時熒光定量PCR儀進行qRT-PCR相關基因的表達量分析。反應使用3個生物學重復和3個技術重復進行,并使用2-ΔΔCT 法進行相對表達量計算[23]。qRT-PCR結果同RNA-seq測定的相應基因的TPM值進行比較分析,驗證測序結果。試驗中使用的基因和引物序列如表1所示。
2 結果與分析
2.1 轉錄組測序、組裝和注釋概述
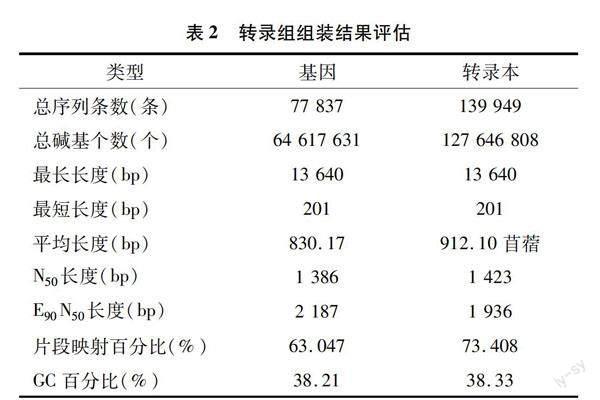
通過使用Illumina HiSeq 2500平臺,對抗薊馬苜蓿品種草原4號(R_CK)和感薊馬苜蓿品種草原2號(S_CK)的轉錄組進行分析。每個處理3個獨立的生物學重復,共完成6個樣品的轉錄組分析,獲得82.34 Gb clean data。使用Trinity對所有樣本clean data進行De novo組裝,組裝得到的unigene個數為77 837條,轉錄本個數為139 949條,平均序列長度為830.17 bp,N50平均長度為1 386 bp,E90N50 平均長度為2 187 bp(表2)。
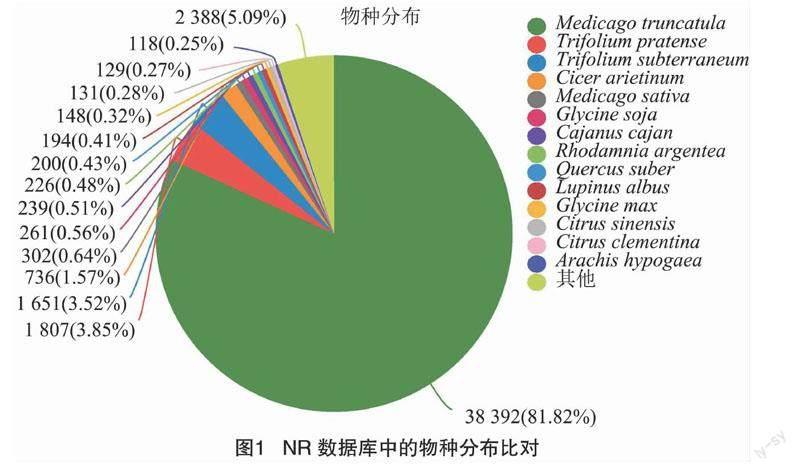
通過所有unigenes和轉錄本分別與NR、Swiss-Prot、Pfam、COG、GO 和 KEGG 等數據庫進行比對,結果發現在75 756個組裝的unigenes中,發現共有46 789個unigenes被注釋到六大數據庫中,分別為NR (46 152)、Swiss-Prot(28 837)、Pfam (28 506)、eeNOG(33 999)、GO(39 565) 和 KEGG(17 270),且其中11 511個unigenes在6個數據庫共同注釋(圖1-a)。通過與NR庫的比對,發現 38 392 個(81.82%)注釋的unigenes與蒺藜苜蓿的序列匹配(圖1-b)。
2.2 差異基因分析
樣品的主成分分析(PCA)表明,S_CK和R_CK之間存在明顯差異,且2個品種的3個樣品分別聚在一起,說明樣本間差異主要來源于品種差異,且測試樣品重復性較好,可以依據品種進行差異基因分析(圖2-a)。如圖2-b所示,與草原2號相比,草原4號共檢測到差異基因2 388個,其中上調基因1 361個,下調基因1 027個。
KEGG注釋分析發現,與草原2號相比,草原4號中有174個上調基因和212個下調基因分別注釋到代謝、遺傳信息處理、環境信息處理、細胞過程、生物體系統等五大類KEGG代謝通路的18個不同的代謝途徑中(圖3-a、圖3-b)。其中,上調基因主要注釋到能量代謝、碳水化合物代謝等代謝過程以及折疊、分類和降解、復制和修復、轉錄和翻譯等遺傳信息處理相關途徑(圖3-a);下調基因主要注釋到折疊、分類和降解、復制和修復等遺傳信息處理相關途徑以及屬于細胞過程的轉運和催化途徑和環境適應相關途徑(圖3-b)。
Go富集分析表明,草原4號和草原2號差異基因主要富集在端粒組織、端粒維持、解剖結構穩態等生物過程中, 說明2個品種本身的遺傳特點可能同抗薊馬性直接相關(圖4)。
2.3 差異基因的qRT-PCR基因驗證
為了確認通過RNA-Seq鑒定的不同基因表達的重現性和準確性,隨機選取了8個基因進行qRT-PCR驗證。如圖5所示,qRT-PCR結果表明草原4號中TRINITY_DN10608_c0_g2、TRINITY_DN12291_c0_g1和TRINITY_DN17228_c0_g1等3個基因的表達量都極顯著高于草原2號,而TRINITY_DN11714_c0_g1、TRINITY_DN11779_c0_g2、TRINITY_DN10336_c0_g1、TRINITY_DN10194_c1_g1和TRINITY_DN1145_c1_g1等5個基因的表達量都顯著低于草原2號,8個基因的qRT-PCR表達量和轉錄組測序TPM值(RNA-Seq)的表達趨勢一致,說明RNA-Seq測得的基因相對表達量結果可靠。
3 討論
隨著國家“振興奶業苜蓿發展行動”計劃的實施及我國農業產業結構的調整,我國苜蓿種植面積逐年增加,僅內蒙古目前苜蓿人工草地保有面積超過1 000萬畝(1 hm2=15畝)。然而,蟲害已經成為影響苜蓿產量和品質的重要因素。無論是草產業發展還是生態建設均需要大量抗病蟲的優良苜蓿品種。植物抗病蟲新品種選育及病蟲害防治受多方面因素影響,通過深入了解植物的抗蟲機制,特別是多角度闡釋苜蓿抗薊馬的機理,對苜蓿抗蟲品種選育及薊馬防治具有重要的理論意義[24]。隨著生物技術的發展,對于植物抗薊馬的分子機制也多有報道。研究者通過QTL圖譜在胡椒的第6染色體上定位到了抗薊馬位點并開展了抗薊馬基因功能的研究[25-26],相似的研究在辣椒[27]和番茄[28]中都有開展。此外,Tu等利用轉錄組測序技術,以甘農 1 號和 WL323 苜蓿品種為研究對象,發現苜蓿誘導性抗薊馬與類黃酮生物合成、β-丙氨酸代謝及水楊酸代謝途徑的基因密切相關[4]。
生物體的表型由 DNA、RNA、蛋白質及代謝物等多個層面調控決定,mRNA研究可以獲得樣本之間的差異基因、復雜調控網絡。轉錄組是連接基因組遺傳信息與生物功能的必然紐帶,是解析生物分子調控機理的重要手段[29]。在前期研究的基礎上,本研究以自主培育的抗薊馬苜蓿品種草原4號為研究對象,采用轉錄測序技術從轉錄水平探索苜蓿組成型抗薊馬的調控機制。通過對測序結果的整體評估,表明測序結果質量較好。PCA結果表明草原4號和草原2號品種間差異較大,且各處理間重復性較好,為差異基因的篩選鑒定提供保證。該結果與Tu等報道的結果[14]一致,說明抗薊馬品種和感薊馬品種在基因型上存在明顯的差異。對差異基因的功能注釋發現,抗薊馬品種草原4號上調基因主要注釋到遺傳信息處理相關途徑、細胞過程和環境適應相關途徑, 表明草原4號抗薊馬特性和其本身的遺傳信息直接相關。研究發現,植物初級代謝如碳水化合物、氮代謝相關產物在植物對昆蟲的組成和誘導型防御中都具有重要作用[30]。本研究發現與感蟲品種草原2號相比,草原4號下調基因主要注釋到能量代謝、碳水化合物代謝等初生代謝過程以及遺傳信息處理相關途徑。因此,筆者所在課題組推測,在苜蓿組成型抗薊馬防御反應中,初生代謝途徑可能扮演著更重要的角色。與此不同的是大量的報道發現,在植物的組成或誘導型抗薊馬過程中,生物堿、黃酮類、萜類等次生代謝產物更為重要[4,6,16,31]。本研究結果為深入闡述苜蓿組成型抗薊馬機理、薊馬防治及利用生物技術手段進行抗薊馬植物品種選育提供理論支撐。
4 結論
轉錄組測序結果表明,草原4號紫花苜蓿組成型抗蟲與其本身遺傳信息處理相關途徑、細胞過程和環境適應相關途徑密切相關,同時能量代謝、碳水化合物代謝等初生代謝途徑在苜蓿組成型抗薊馬防御反應中具有重要作用。研究結果為揭示苜蓿組成型抗薊馬機理奠定了基礎,進而為抗薊馬新品種培育及薊馬防治提供理論支撐。
參考文獻:
[1]張 奔,周敏強,王 娟,等. 我國苜蓿害蟲種類及研究現狀[J]. 草業科學,2016,33(4):785-812.
[2]Wu S Y,Tang L D,Zhang X R,et al. A decade of a thrips invasion in China:lessons learned[J]. Ecotoxicology,2018,27(7):1032-1038.
[3]Steenbergen M,Broekgaarden C,Pieterse C M J,et al. Bioassays to evaluate the resistance of whole plants to the herbivorous insect Thrips[J]. Methods in Molecular Biology (Clifton,N.J.),2020,2085:93-108.
[4]Tu X B,Liu Z K,Zhang Z H.Comparative transcriptomic analysis of resistant and susceptible alfalfa cultivars (Medicago sativa L.) after thrips infestation[J]. BMC Genomics,2018,19(1):116.
[5]楊青川. 苜蓿生產與管理指南[M]. 北京:中國林業出版社,2003:167-171.
[6]Steenbergen M,Abd-el-Haliem A,Bleeker P,et al. Thrips advisor:exploiting thrips-induced defences to combat pests on crops[J]. Journal of Experimental Botany,2018,69(8):1837-1848.
[7]Gao Y L,Lei Z R,Reitz S R.Western flower thrips resistance to insecticides:detection,mechanisms and management strategies[J]. Pest Management Science,2012,68(8):1111-1121.
[8]van Haperen P,Voorrips R E,Lucatti A F,et al. The effect of a thrips resistance QTL in different Capsicum backgrounds[J]. Euphytica,2020,216(12):187.
[9]Wu F,Shi S L,Li Y Z,et al. Physiological and biochemical response of different resistant alfalfa cultivars against thrips damage[J]. Physiology and Molecular Biology of Plants,2021,27(3):649-663.
[10]Wahyuni Dinar S C,Hae C Y,Leiss K A,et al. Morphological and chemical factors related to western flower thrips resistance in the ornamental Gladiolus[J]. Plants,2021,10(7):1384.
[11]Gibson R W. Glandular hairs providing resistance to aphids in certain wild potato species[J]. Annals of Applied Biology,1971,68(2):113-119.
[12]Macel M,Visschers I G S,Peters J L,et al. High concentrations of very long chain leaf wax alkanes of thrips susceptible pepper accessions (Capsicum spp.)[J]. Journal of Chemical Ecology,2020,46(11):1082-1089.
[13]Tayal M,Somavat P,Rodriguez I,et al. Polyphenol-rich purple corn pericarp extract adversely impacts herbivore growth and development[J]. Insects,2020,11(2):98.
[14]Singh S,Kariyat R R. Exposure to polyphenol-rich purple corn pericarp extract restricts fall armyworm (Spodoptera frugiperda) growth[J]. Plant Signaling & Behavior,2020,15(9):1784545.
[15]Mouden S,Leiss K A. Host plant resistance to thrips (Thysanoptera:Thripidae)-current state of art and future research avenues[J]. Current Opinion in Insect Science,2021,45:28-34.
[16]Zhang Z Q,Chen Q,Tan Y,et al. Combined transcriptome and metabolome analysis of alfalfa response to thrips infection[J]. Genes,2021,12(12):1967.
[17]特木爾布和,肖燕子. 苜蓿新品種草原4號的選育[J]. 草業科學,2017,34(4):855-860.
[18]劉玉良. 抗薊馬苜蓿新品系抗性機理研究[D]. 呼和浩特:內蒙古農業大學,2009.
[19]武杰瑞,劉秉毅,特木爾布和,等. 薊馬取食對苜蓿氨基酸組成及含量的影響[J]. 草地學報,2022,30(1):62-68.
[20]薩如拉,張志強,偉勒斯,等. 兩個苜蓿品種營養器官解剖結構特征比較[J]. 西北植物學報,2020,40(11):1881-1887.
[21]吳永敷,薇 玲,趙秀華,等. 苜蓿對薊馬的抗源篩選試驗[J]. 中國草地,1990,12(5):61-63,65.
[22]Young M D,Wakefield M J,Smyth G K,et al. Gene ontology analysis for RNA-seq:accounting for selection bias[J]. Genome Biology,2010,11(2):R14.
[23]Zhang Z Q,Wang Y F,Chang L Q,et al. MsZEP,a novel zeaxanthin epoxidase gene from alfalfa (Medicago sativa),confers drought and salt tolerance in transgenic tobacco[J]. Plant Cell Reports,2016,35(2):439-453.[HJ2.05mm]
[24]劉啟航,曹進軍,付曉偉,等. 波譜光照效應對西花薊馬趨光響應變化的影響[J]. 江蘇農業科學,2021,49(20):124-132.
[25]Maharijaya A,Vosman B,Steenhuis-Broers G,et al. QTL mapping of thrips resistance in pepper[J]. Theoretical and Applied Genetics,2015,128(10):1945-1956.
[26]Maharijaya A,Vosman B,Pelgrom K,et al. Genetic variation in phytochemicals in leaves of pepper (Capsicum) in relation to thrips resistance[J]. Arthropod-Plant Interactions,2019,13(1):1-9.
[27]van Haperen P,Voorrips R E,van Loon J J A,et al. The effect of plant development on thrips resistance in Capsicum[J]. Arthropod-Plant Interactions,2019,13(1):11-18.
[28]Bac-molenaar J A,Mol S,Verlaan M G,et al. Trichome independent resistance against western flower thrips in tomato[J]. Plant and Cell Physiology,2019,60(5):1011-1024.
[29]Zhou S,Lou Y,Tzin V,et al. Alteration of plant primary metabolism in response to insect herbivory[J]. Plant Physiology,2015,169(3):1488-1498.
[30]Wainberg M,Sinnott-Armstrong N,Mancuso N,et al. Opportunities and challenges for transcriptome-wide association studies[J]. Nature Genetics,2019,51(4):592-599.
[31]Macel M,Visschers I G S,Peters J L,et al. Metabolomics of rhrips resistance in pepper (Capsicum spp.) reveals monomer and dimer acyclic diterpene glycosides as potential chemical defenses.[J]. Journal of Chemical Ecology,2019,45(5/6):490-501.
收稿日期:2022-04-20
基金項目:國家自然科學基金(編號:32160333、32060388);內蒙古自然科學基金(編號:2021MS03011)。
作者簡介:爽 爽(1999—),女,內蒙古通遼人,碩士研究生,主要從事草學方面的研究。E-mail:bss@emails.imau.edu.cn。
通信作者:張志強,博士,副教授,主要從事牧草抗逆及品種選育方面的研究。E-mail:zhangzq19890102@126.com。
