青花菜BoWHY2基因克隆及其高溫脅迫下的表達特征
2023-06-14 00:30:20裴徐梨荊贊革施俊宏胡文婷
貴州農業科學 2023年6期
關鍵詞:植物
裴徐梨,荊贊革,2,施俊宏 ,胡文婷,唐 征,焦 鵬
(1.昆明學院 農學與生命科學學院,云南 昆明 650214;2.溫州科技職業學院 農業與生物技術學院,浙江 溫州 325006)
0 引言
【研究意義】Whirly(WHY)基因家族是植物特有的一類轉錄因子,其家族成員可參與植物多種生命進程的調控[1]。首個被鑒定的Whirly家族成員為馬鈴薯StWHY1,其與誘導應答元件ERE相結合,從而調節抗病基因的表達[2]。青花菜屬于十字花科蕓薹屬植物,營養成分齊全,并含有抗癌物質葡糖異硫氰酸鹽,具有特殊的保健作用。因此,研究青花菜WHY基因有助于探討其在逆境脅迫下的應答機制,對選育適應性廣的青花菜新品種意義重大。【前人研究進展】目前,植物WHY基因的研究相對較少,僅在杧果[3]、水稻[4]、辣椒[5]、黃瓜[6]、大豆[7]、番茄[8]和擬南芥[9]等鑒定出WHY基因。WHY基因家族在非生物脅迫和抗病等方面發揮著重要的生物學功能[10]。三明野生蕉WHY基因在8℃條件下表達量變化明顯,說明該基因可能在抗寒過程中發揮著重要作用[11];在鹽脅迫處理下,天寶蕉WHY基因表現出明顯的抗性反應,表明該基因可能在天寶蕉抗鹽脅迫過程中發揮著重要作用[12]。此外,SiWHY2轉基因型的煙草可通過緩解青枯假單胞菌造成的ROS積累,抵抗病菌的侵染擴散,推測SiWHY2可能通過參與水楊酸依賴的抗病響應途徑增強植株的抗病性;茉莉酸甲酯可誘導超表達Whirly2擬南芥葉片更早出現衰老癥狀,說明在茉莉酸酯處理下,Whirly2對植物葉片的衰老具有調控作用[2]。【研究切入點】青花菜喜冷涼,高溫是其生長發育的重要影響因素。雖WHY基因的分子機制在模式植物中研究較多,但其在青花菜高溫脅迫下的調控機制仍不清楚。【擬解決的關鍵問題】通過克隆青花菜BoWHY2基因,分析其序列特征和分子進化,檢測其高溫脅迫下的表達特征,為進一步研究青花菜BoWHY2在高溫脅迫下的應答機制提供理論依據。
1 材料與方法
1.1 材料
試驗材料為青花菜高代自交系WN12-95B,由昆明學院青花菜遺傳育種課題組選育。常規育苗,五葉期時進行高溫脅迫(38℃)處理3 h和12 h,每個處理重復3次。
1.2 方法
1.2.1 青花菜BoWHY2基因的克隆 提取青花菜葉片總RNA,使用Prime Script 1ststrand cDNA Synthesis Kit試劑盒合成cDNA第1鏈。以青花菜轉錄組數據中的WHY2基因序列為模板,設計特異引物(F:5′-ATGATGAAGCAAGCTCGCACT-3′和R:5′-TTACTCTTTACCCCACTCCAGCTC-3′)進行PCR擴增。PCR擴增程序和體系參照裴徐梨等[13]的方法。
1.2.2 青花菜BoWHY2序列特征分析 利用ProtParam tool、ProtScale和DNAMAN等工具分析BoWHY2的等電點、相對分子量和親疏水性等序列特征。采用GOR4和SWISS-MODEL預測該蛋白的高級結構[13]。
1.2.3 青花菜BoWHY2同源性和系統進化分析 利用BLAST分析青花菜BoWHY2基因的序列同源性。采用MEGA 5.0構建系統進化樹[14]。
1.2.4 青花菜BoWHY2基因的表達特征分析 青花菜BoWHY2基因在高溫脅迫下的轉錄組數據來源于昆明學院青花菜遺傳育種課題組前期的高通量測序結果。基因表達量經過標準化處理,用FPKM值表示。
2 結果與分析
2.1 青花菜BoWHY2基因的克隆
由圖1可見,青花菜BoWHY2基因PCR擴增獲得700 bp左右的條帶,經回收純化后進行TA克隆,測序顯示該序列長度為768 bp,包含上下游引物序列(圖2),將其命名為BoWHY2。
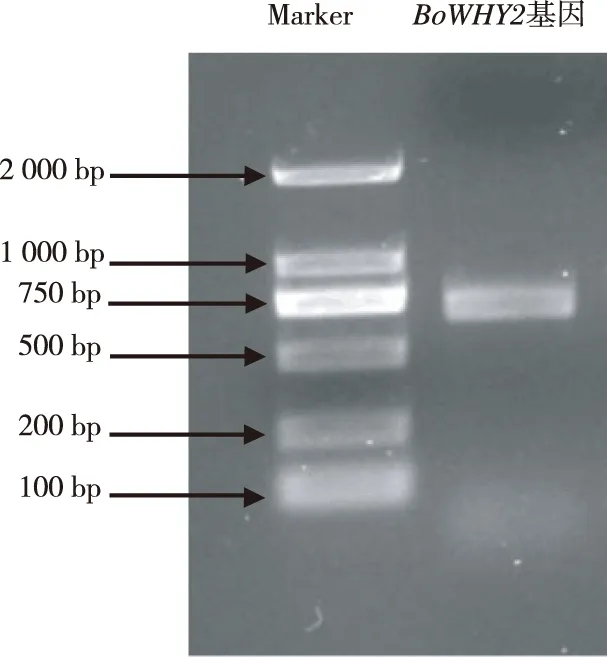
圖1 青花菜BoWHY2基因PCR擴增產物Fig.1 PCR products of broccoli BoWHY2 gene

圖2 青花菜BoWHY2基因及其推導的氨基酸序列Fig.2 Broccoli BoWHY2 gene and its deduced amino acid sequence
2.2 青花菜BoWHY2序列特征
青花菜BoWHY2的蛋白分子量為26.2 kD,理論等電點為9.48,屬于堿性蛋白。該蛋白脂肪系數為74.42。ProtScale和DNAMAN分析結果顯示,其親水性較差,疏水性較好。二級結構主要由α螺旋(21.67%)、延長線(20.42%)和無規則卷曲(57.92%)組成。三維結構預測顯示,該蛋白表面有1個中心空穴并形成帶電的內腔。
2.3 青花菜BoWHY2基因序列同源性和進化樹
將青花菜BoWHY2基因的核苷酸序列進行NCBI-BLAST比對發現,該基因核苷酸序列與甘藍(Brassicaoleracea,XM_013734587.1)、油菜(Brassicanapus,XM_013843540.2)、白菜(Brassicarapa,XM_009107547.3)和蘿卜(Raphanussativus,XM_018598754.1)的核苷酸同源性分別是100%、97.79%、96.69%和92.76%,核苷酸相似性均在90%以上。與鹽芥(Eutremasalsugineum,XM_006390725.2)、亞麻薺(Camelinasativa,XM_010429650.2)、河岸葡萄(Vitisriparia,XM_034818773.1)、薺菜(Capsellarubella),XM_006302697.2)、葡萄(Vitisvinifera,MN395405.1)和醉蝶花(Tarenayahassleriana,XM_010536568.2)的核苷酸同源性分別是82.46%、81.31%、79.63%、79.58%、78.75%和74.61%,核苷酸相似性均在70%以上。
將青花菜BoWHY2核苷酸序列與多個植物構建進化樹(圖3)。該進化樹可分為兩大組,第一大組中屬十字花科的甘藍、白菜、油菜、蘿卜、鹽芥、擬南芥、亞麻芥等聚在同一亞組中,而醉蝶花單獨為一亞組。葡萄科的3個物種聚為第二大組。
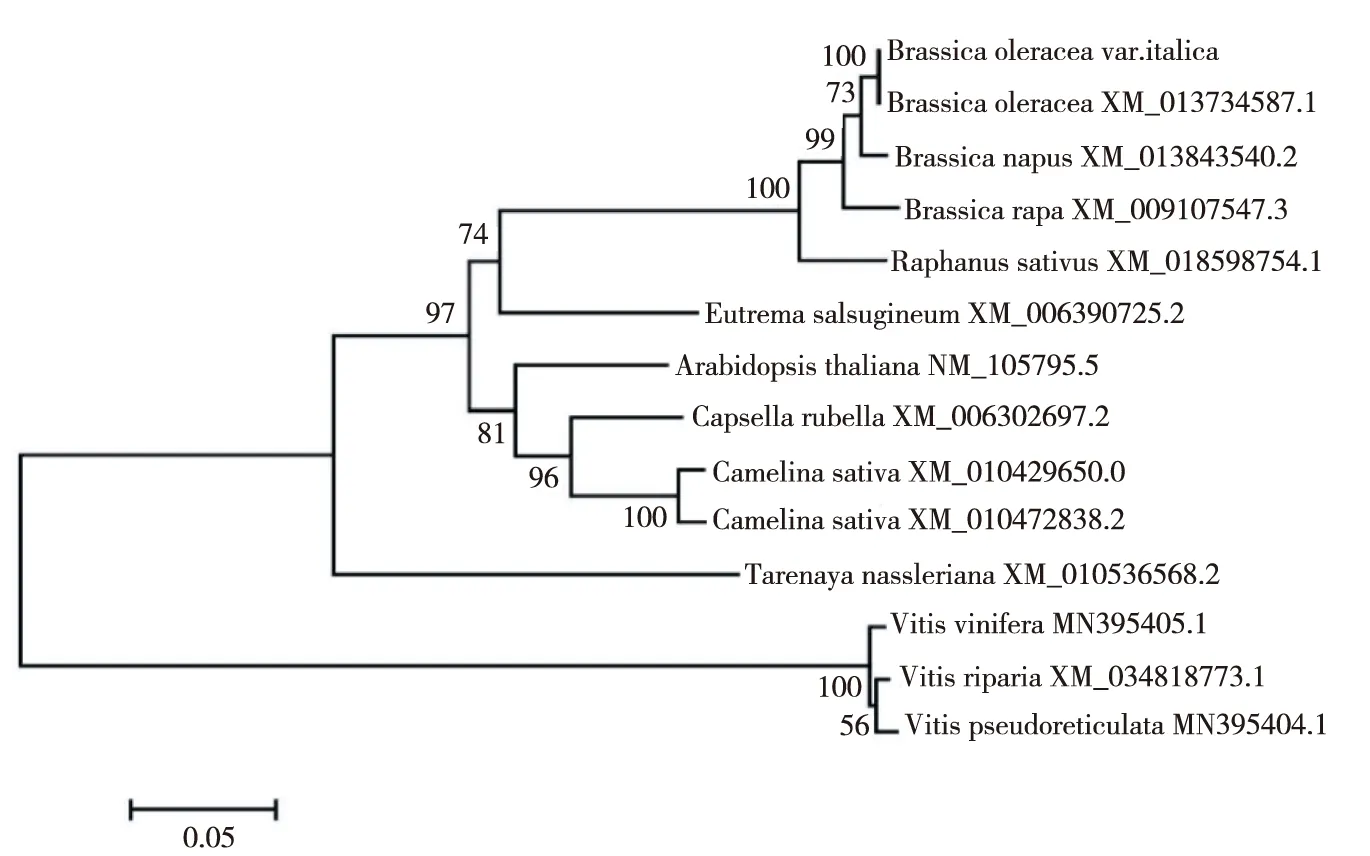
圖3 青花菜BoWHY2系統進化樹Fig.3 The phylogenetic tree of broccoli BoWHY2 gene
2.4 青花菜BoWHY2基因在高溫脅迫下的表達特征
由圖4可見,高溫脅迫處理12 h內,青花菜BoWHY2基因的表達量先下降后上升。高溫脅迫下處理3 h時,青花菜BoWHY2基因的表達量逐漸下降至2.15;高溫脅迫下處理12 h時,BoWHY2基因的表達量持續上升至4.76,是對照組的1.3倍。
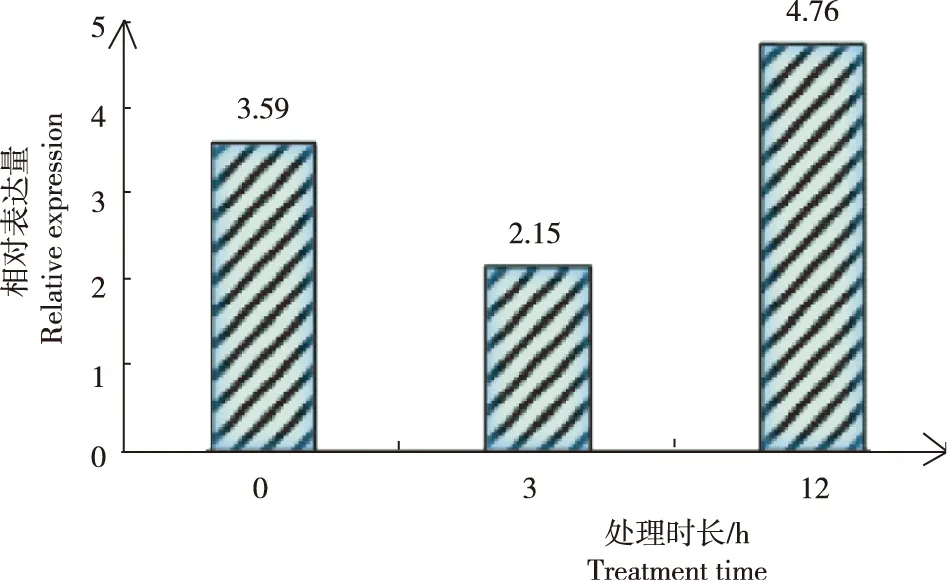
圖4 青花菜BoWHY 2基因高溫脅迫下的表達特征Fig.4 Expression characteristics of broccoli BoWHY2 gene under high temperature stress
3 討論
利用PCR技術克隆得到的青花菜BoWHY2基因其序列特征分析結果與前人研究相似[15],表明植物WHY家族的保守型較強,具有類似的生物學功能。通過與其他植物的WHY家族基因進行序列比對發現,青花菜與十字花科植物的親緣關系較近,和甘藍、油菜、白菜的同源性均在90%以上。因此,推測Whirly基因的保守性較強,在不同植物中可能有著相似的生物學功能。
非生物脅迫是影響植物生長發育的重要因素。Whirly基因參與了三明野生蕉的抗低溫反應,在抗寒過程中發揮著重要的作用。天寶蕉Whirly可被低溫處理誘導,推測其參與了抗低溫反應,在抗寒相關基因的轉錄過程中發揮作用[12];王潔如[16]研究表明,Whirly基因表達量隨低溫處理時間的總體變化趨勢與天寶蕉、三明野生蕉的研究結果相似,說明該基因對低溫脅迫能夠作出積極反應。多個GmWHY的表達量在鹽脅迫處理24 h和48 h的葉片中明顯上調,表明GmWHY基因可能參與了鹽脅迫的調控[7]。研究結果表明,青花菜BoWHY2在高溫處理3 h后,基因表達量上調;高溫處理12 h后,基因表達量增加,高于對照。說明BoWHY2基因對高溫脅迫能夠作出正向反應,推測該基因在青花菜抗高溫脅迫上有一定的作用。
4 結論
從青花菜克隆的BoWHY2基因長度為768 bp,編碼240個氨基酸,該蛋白分子量為26.2 kD,屬于堿性蛋白,疏水性較好,理論等電點為9.48。青花菜BoWHY2基因與甘藍和油菜的進化關系較近,保守性較強,該基因在高溫脅迫下其表達量持續升高,對高溫脅迫能夠作出正向反應。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
