遠志葉綠體基因組序列特征與系統發育分析
2023-06-08 12:21:52王星璐李慧娟李彥青秦雪梅張福生
中草藥 2023年11期
王星璐,李慧娟,趙 偉,李彥青,秦雪梅,張福生*
1. 山西大學 中醫藥現代研究中心,山西 太原 030006
2. 山西中醫藥大學基礎醫學院,山西 晉中 030619
在植物中核基因組的復雜性使得低拷貝基因的篩選比較困難,目前僅局限在基因組相對較小的模式物種或具有重要經濟生態價值的物種中,整體應用范圍較為有限。植物線粒體基因組具有在植物類群中變異很大、存在基因組之間水平轉移的外源基因和進化速率較慢等特性,使其在系統發育研究中的應用受到了限制[1]。葉綠體基因組作為第2 大基因組,屬于相對獨立于核基因組之外的基因組[2],其基因組完整且相對獨立。大多數被子植物的葉綠體含有大量遺傳信息,同時具有相對分子質量小、結構簡單,進化速率中等、突變率較低、遺傳穩定、成本低及開發難度低、廣泛分布著微衛星序列和線粒體 DNA 等特點[3-4],在一定程度上彌補了線粒體和核基因組的部分缺點與不足。
葉綠體起源于古細菌入侵植物細胞[5],是一種擁有自身遺傳物質的多功能細胞器[6],為質體中的一種類型。葉綠體基因組的結構大部分為共價閉合的雙鏈狀環形分子,也有少部分是線性分子或者多聚體。對于高等植物而言,葉綠體存在于細胞質的基質中,其形狀呈現為綠色橢圓球型或者是圓球形[7]。典型的環式雙鏈葉綠體基因組結構,包括4 個部分,即大單拷貝區(large single copy,LSC),小單拷貝區(small single copy,SSC)和2 個反向重復區(inverted repeats,IRs);其中,2 個反向重復區把基因組分隔為大單拷貝區和小單拷貝區[8]。LSC 區長81~90 kb,SSC 區介于18~20 kb;2 個IR 區序列基本一致,大小介于20~30 kb,是葉綠體基因組進化過程中延展或縮小的區域[9]。大多數陸生植物的葉綠體基因組中有110~130 個基因[10]。近年來,由于葉綠體基因組大小、結構和基因種類一般較為保守,葉綠體全基因組的物種鑒定及系統進化研究成為植物系統分類學的一個新趨勢,為研究藥用植物系統進化及進行物種鑒定提供可靠工具[11]。目前,葉綠體基因組技術已經廣泛應用于藥用植物的研究中[12-14]。
遠志植物為雙子葉植物蕓香目遠志科一種一年生或多年生草本、灌木或小喬木,約500 種,廣布于全世界,我國有42 種8 變種,廣布于全國各地,而以西南和華南地區最盛[15]。遠志屬遠志PolygalatenuifoliaWilld.最早記載于《華氏中藏經》[16],藥用價值廣泛,歷史悠久,具有較大的藥用研究和開發利用價值。近年來由于過度采挖遠志藥材,導致野生藥材資源嚴重不足,遠志已被收入《國家重點保護野生藥材物種名錄》,保護級別為Ⅲ級。目前關于遠志的研究主要集中于其化學成分與藥理作用等方面。研究發現,遠志具有改善學習記憶、抗氧化、抗抑郁、抗衰老、鎮靜催眠、益智和抗腫瘤以及祛痰鎮咳、影響藥物代謝以及抗炎等[17-24]作用。分子生物方面研究主要集中在遠志隨機擴增多態性DNA 分析[25]、遺傳多樣性[26]、遠志譜系地理學[27-28]、序列鑒定[29-30]等方面;對葉綠體基因組的研究報道較少,雖有對西南遠志P.crotalarioidesBuch.-Ham. ex DC.[31]和遠志P.tenuifoliaWilld.[32]葉綠體基因組進行測序、拼裝和注釋的報道,但是報道的遠志[32]僅限于描述了序列全長和系統發育位置等一些基本信息,未對其展開詳細的分析描述。
目前,遠志屬物種的葉綠體基因組研究十分缺乏,其相關的親緣關系值得深入研究,NCBI 數據庫中有遠志P.tenuifoliaWilld.(NC_050829.1)[32]完整葉綠體基因組數據,但是僅記載了葉綠體基因組總長(165 423 bp)等基礎信息,也未進行深入挖掘研究,相關信息依舊不清楚。本研究利用生物信息學相關軟件,分析其葉綠體基因組的構成,并對其進行基因組裝和注釋,分析序列特征,密碼子偏好性以及系統發育,闡明遠志葉綠體基因組結構特征以及其物種之間的親緣關系。遠志葉綠體基因組的研究是對遠志藥材種質資源遺傳多樣性的進一步了解,有利于遠志藥材優良品種的選育和種質資源的評價及其有效保護與合理利用。
1 材料與方法
1.1 樣品收集
從美國國立生物技術信息中心(National Center of Biotechnology Information,NCBI)數據庫(https://www.ncbi.nlm.nih.gov/)檢索遠志科遠志屬物種的葉綠體全基因組序列信息。檢索到遠志科遠志屬8 個物種以及齒果草屬1 個葉綠體全基因組序列信息,遠志科親緣關系較近的蕓香目苦木科鴉膽子Bruceajavanica(L.) Merr.和蕓香科九里香MurrayaexoticaL. Mant.完整葉綠體全基因組序列。下載檢索到的物種的葉綠體基因組名稱、基因組登錄號(表1)。
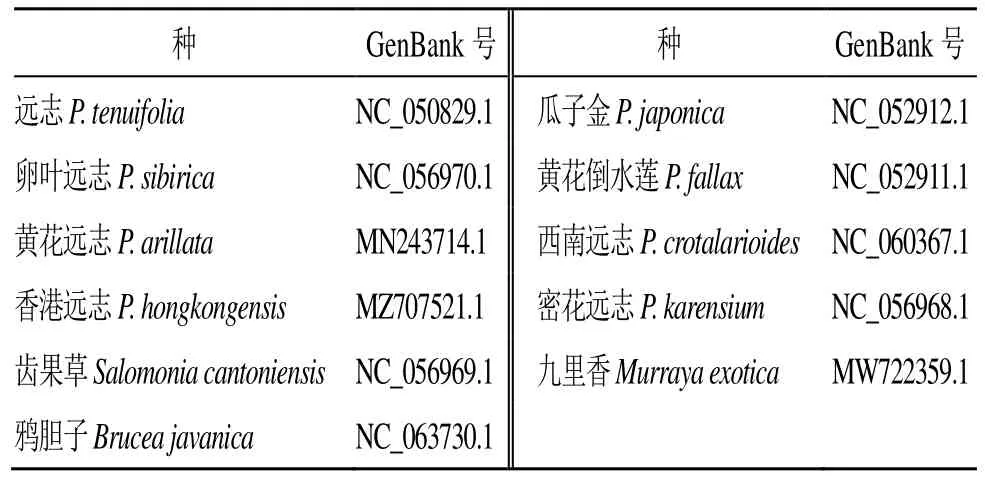
表1 遠志科、苦木科和蕓香科葉綠體GenBank 登錄號Table 1 GenBank accession number of chloroplast in Polygalaceae, Simaroubaceae and Rutaceae
1.2 葉綠體基因組注釋、圖譜繪制以及基本特征分析
遠志屬遠志的葉綠體全基因組序列(登錄號為NC_050829.1 ) 通 過 GeSeq :( https://chlorobox.mpimp-golm.mpg.de/geseq.htm)[33]和Plastid Genome Annotator(PGA)[34]軟件進行基因注釋,將結果對比矯正,去除錯誤及冗余注釋。通過 Chloroplot(https://irscope.shinyapps.io/Chloroplot/)[35]在線繪制工具繪制。通過GeSeq 初步注釋和Chloroplot 畫圖相結合對葉綠體基因組的總長度及各個區域(LSC、SSC、IR)的長度、基因組成(蛋白編碼基因、tRNA基因、rRNA 基因)、堿基組成、GC(AT)含量進行統計和比較分析,解析遠志葉綠體基因組序列的基本特征。
1.3 葉綠體全基因組重復序列檢測
利用Perl 語言通過MISA 軟件(http://pgrc.ipk-gatersleben.de/misa/misa.html)[36]完成簡單重復序列(simple sequence repeats,SSRs)檢測,參數設置為單核苷酸重復單元不少于10 個,二核苷酸重復單元不少于5 個,三核苷酸和四核苷酸重復單元不少于4 個,五核苷酸和六核苷酸重復單元不少于3 個,且2 個SSRs 之間的最小距離為100 bp[37]。
葉綠體全基因組中的散在重復序列(dispersed repeats)利用REPute 軟件(https://bibiserv.cebitec.uni-bielefeld.de/reputer)[38]進行檢測,正向重復(forward repeats,F)、反向重復(reverse repeats,R)、互補重復(complement repeats,C)、回文重復(palindromic repeats,P)。參數設置最小重復長度(minimal repeat size)設置為30,hamming 距離(hamming distance)為 3,最大計算重復次數(maximum computed P repeats)5000[39]。串聯重復序列(tandem repeats)利用Tandem repeats finder軟件(https://tandem.bu.edu/trf/trf.html)進行檢測。參數設置選擇默認值[40]。
1.4 密碼子使用分析
采用CodonW(http://codonw.sourceforge.net)軟件[41]分析密碼子使用情況。對遠志的葉綠體基因組同義密碼子相對使用頻次( relative synonymous codon usage,RSCU)進行分析和統計。當RSCU>1 時,表明該密碼子使用頻率較高;RSCU=1 時,說明該密碼子無偏好性;RSCU<1時,表明該密碼子使用頻率較低[42]。
1.5 基因組比較分析
采用Geneious[43]軟件統計遠志屬8 個物種的葉綠體基因組序列的4 個邊界(SSC、LSC、IRa和IRb 區域)長度和基因數目類型、GC 含量等信息,并用EXCEL 計算各自的GC 值。使用IRscope(https://irscope.shinyapps.io/irapp/)[44]可視化工具,比較遠志屬葉綠體基因組4 個區域邊界的差異。并采用mVISTA 對其進行全基因組比對分析。
1.6 基于葉綠體基因組序列的系統進化分析
為了確定遠志的系統發育位置,下載NCBI 數據庫中收錄的遠志科上述所有9 種植物的葉綠體全基因組序列。與本研究遠志葉綠體全基因組序列共同構建序列矩陣,同時選擇遠志屬的近緣類鴉膽子和九里香作為外類群。利用MAFFT 7 軟件(https:// mafft.cbrc.jp/alignment/software/)[45]完成序列比對。利用IQtree2.0.5(http://www.iqtree.org/)[46]以最大似然法(maximum likelihood,ML)進行系統發育關系重建分析,核苷酸替換模型選擇GTR+G,其他參數選擇默認設置,進行1000 次自展重復分析,計算系統發育樹各節點的靴帶支持率(bootstrap values,BS)。并利用MEGA X 軟件,使用鄰接法(neighbor-joining,NJ)構建系統進化樹,并進行系統發育樹的比較。
2 結果與分析
2.1 遠志葉綠體基因組的基本特征
遠志的葉綠體基因組與典型的陸地植物葉綠體基因組基本特征一樣,均為典型的四分體結構,遠志屬遠志葉綠體基因組呈雙鏈環狀四分體結構(圖1),大小為165 423 bp,包括大單拷貝區、小單拷貝區以及2 個反向重復區,各區長度分別為83 699、8044、36 840 bp。遠志葉綠體基因組的總GC 量為36.7%,IR 區的GC 量為39.7%,高于LSC 區(34.7%)及SSC 區(29.4%)。整體而言,IR 區的保守性高于LSC 區和SSC 區,這可能是IR 區的GC 量高于LSC 區和SSC 區而造成的。計算各分區的堿基組成(表2)。
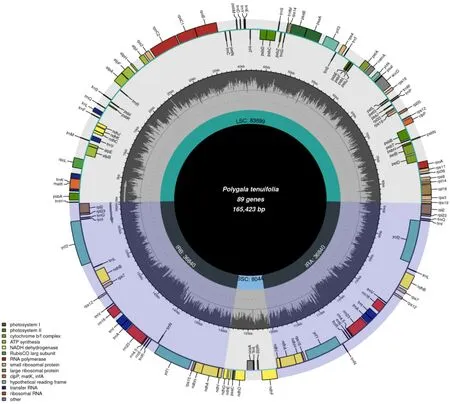
圖1 遠志葉綠體基因組圖譜Fig. 1 Chloroplast genome map of P. tenuifolia
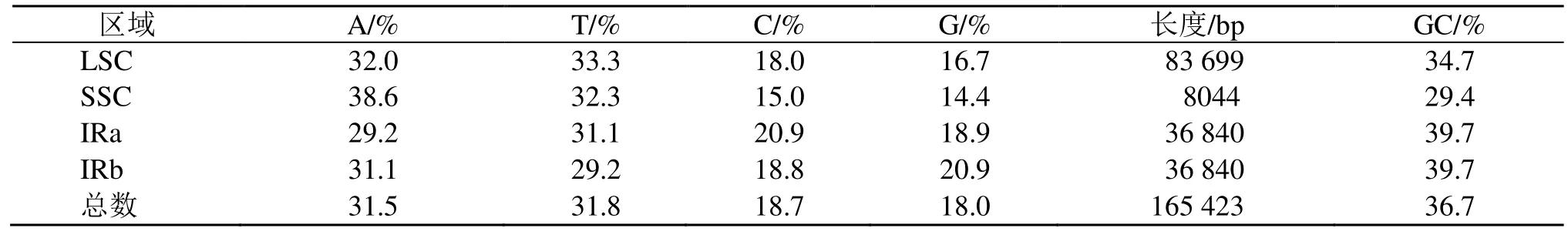
表2 遠志葉綠體基因組堿基組成Table 2 Base composition of chloroplast genome of P. tenuifolia
2.2 遠志葉綠體基因組的基因注釋及歸類分析
遠志葉綠體基因組共注釋得到135 個基因,包括8 個rRNA 基因、38 個tRNA 基因和89 個蛋白編碼基因。根據其功能可分為以下4 個大類(表3)。第1 類:48 個與光合作用相關的基因,包括5 個光合系統I基因、15 個光合系統II 基因、6 個細胞色素b/f復合體基因、6 個ATP 合酶基因、1 個二磷酸核糖體羧化酶、15 個NADH 氧化還原酶基因。第2 類:除rRNA 基因和tRNA 基因外,有30 個與自我復制相關的基因,包括10 個核糖體蛋白大亞基基因、16 個核糖體蛋白小亞基基因和4 個RNA 聚合酶亞基基因。第3 類:6 個其他編碼蛋白質的基因。第4 類:6 個功能未知的基因。在這些基因中,有12 個蛋白編碼基因(rpl2、rpl23、ycf2、rps12、rps7、rps15、ycf1、ndhA、ndhB、ndhH、ndhI、ndhN)、7 個tRNA 編碼基因(trnQ、trnI、trnL、trnV、TrnA、trnN、trnR)和4 個rRNA 編碼基因(rrn4.5、rrn5、rrn16、rrn23)位于IR 區。

表3 遠志葉綠體基因組基因功能注釋與分類Table 3 Gene functional annotation and classification of P. tenuifolia chloroplast genome
2.3 重復序列統計分析
葉綠體基因組SSR 分析在遠志葉綠體基因組中搜索到161 個SSR 位點。其中,29 個為復合型SSRs;單核苷酸重復Motif 位點最多為113 個,二核苷酸、三核苷酸、四核苷酸、五核苷酸和六核苷酸重復基序分別有30、3、9、4、2 個(表4)。SSR的類型以A/T 為主,無含有G/C 組成的位點,占總重復類型的70.18%;其次為AT/AT,共有25 個(15.52%),AG/CT(3.11%),其余總數占總重復類型的11.18%。A/T 和AT/AT 占簡單重復序列位點重復單元總數量的85.71%,因此遠志葉綠體基因組富含AT。從分布區段上看,34 個位點位于IR區段,115 個位于LSC 區段,12 個位于SSC 區段,說明遠志葉綠體基因組SSRs 分布的不均勻性。

表4 遠志葉綠體基因組SSRs 位點類型及數量Table 4 Type and number of SSRs of P. tenuifolia chloroplast genome
使用REPuter 軟件在遠志葉綠體基因組中共檢測到223 個散在重復序列,包括正向重復序列(F型)、反向重復(R 型)、回文重復序列(P 型)和互補重復(C 型)4 種類型。其中,正向重復序列(F 型)89 個,反向重復(R 型)13 個,回文重復序列(P 型)109 個和互補重復(C 型)12 個。長度為30~39 bp 的重復序列中包括61 個正向重復,12 個反向重復,72 個回文重復和12 個互補重復。其分布數量相對較多(圖2)。發現90 條長度范圍在2~54 bp 的串聯重復序列。串聯重復序列,4個區域均有分布,其中,LSC 區35 條,重復次數1.9~26.8 次,TRB 區24 條,重復次數由1.9~12.2次,SSC 區1 條,重復次數由2 次,TRA 區30 條,重復次數由1.9~8.1 次。
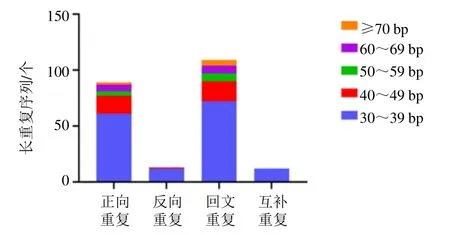
圖2 遠志葉綠體基因組散在重復序列類型與數量Fig. 2 Type and number of long repeats in P. tenuifolia chloroplast genome
2.4 葉綠體基因組密碼子偏好性分析
用CodonW 軟件分析遠志葉綠體基因組蛋白編碼序列密碼子的使用偏好性(圖3),共預測了55 141 個密碼子,其中編碼亮氨酸(Leu)的密碼子數量最多(5629 個,10.21%),編碼色氨酸(Trp)的密碼子數量最少(705 個,1.28%)。在遠志葉綠體基因組中有20 種氨基酸,除甲硫氨酸(Met)和色氨酸(Trp)使用1 個密碼子AUG 和UGG 外,其余氨基酸均有 2~6 個同義密碼子。亮氨酸(Leu)、精氨酸(Arg)、絲氨酸(Ser)使用6 個同義密碼子,使用頻率最高的密碼子分別為UUA、UCU、AGA;丙氨酸(Ala)、甘氨酸(Gly)、脯氨酸(Pro)、蘇氨酸(Thr)、纈氨酸(Val)分別使用4 個同義密碼子,使用頻率最高的密碼子分別為GGA、CCA、GUU、CCU、ACU;異亮氨酸(lle)使用3 個同義密碼子,使用頻率最高的密碼子是AUU;半胱氨酸(Cys)、天冬氨酸(Asp)、谷氨酸(Glu)、苯丙氨酸(Phe)、組氨酸(His)、賴氨酸(Lys)、天冬酰胺(Asn)、谷氨酰胺(Gln)、酪氨酸(Tyr)均使用2 個同義密碼子,但使用頻率并不相同,這9 個氨基酸使用頻率較高的密碼子分別為UUU、AAA、AAU、UAU、UUC、GAA、GAU、AAG、CAA。結果表明,遠志葉綠體基因偏好使用含有A、U 堿基的密碼子,密碼子第3 位也偏好以A 和U 結尾。
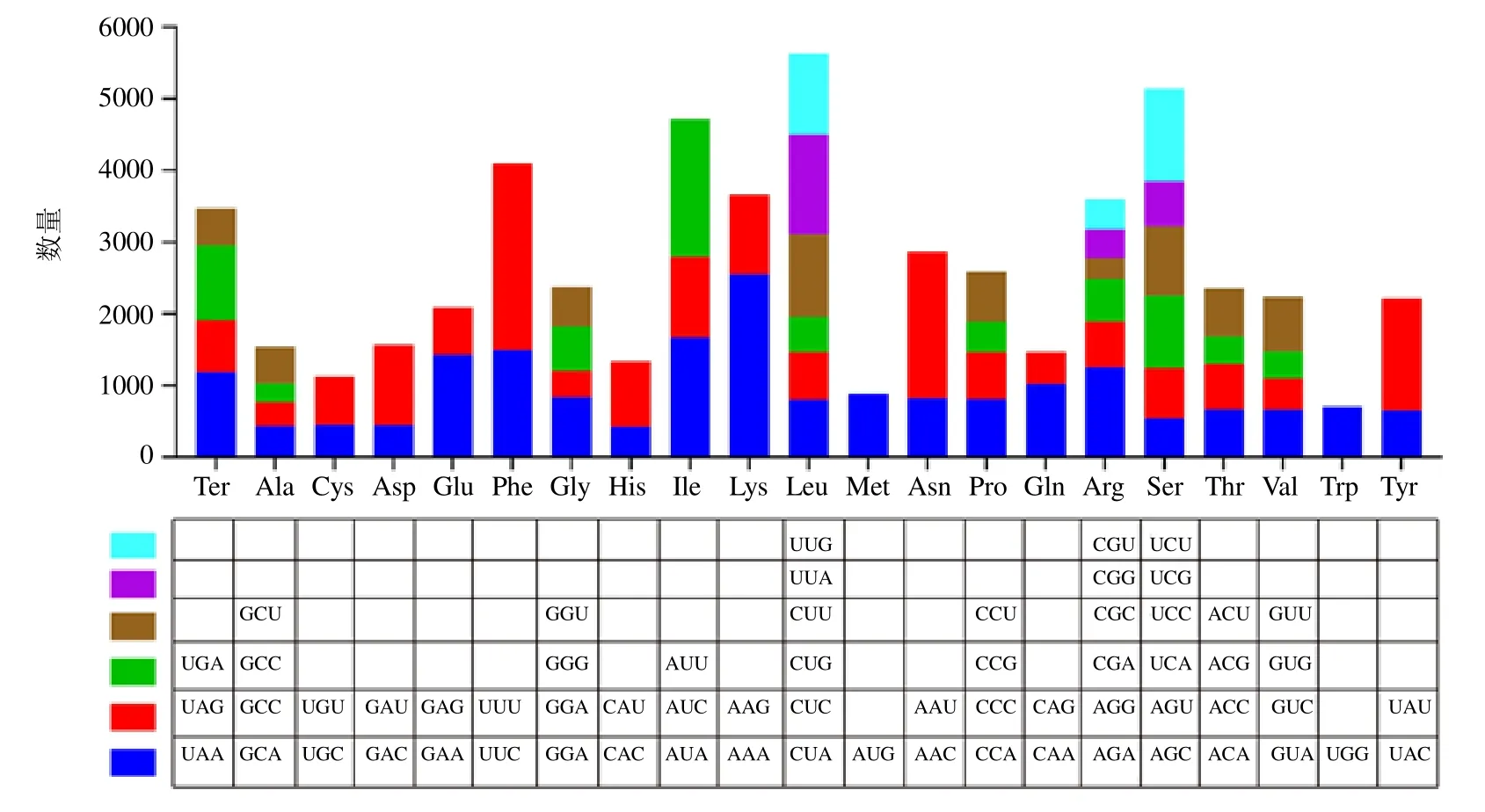
圖3 遠志葉綠體基因組的密碼子使用頻率Fig. 3 Codon usage frequency in chloroplast genome of P. tenuifolia
相對同義密碼子使用度(RSCU)分析表明,在所有被編碼的密碼子中,RSCU>1 的密碼子共有35 個(AGA、UCU、UUA 等),其中27 個密碼子以A/U 堿基結尾,以A 結尾的占51.85%,以U 結尾的占48.15%,故這些密碼子的結尾具有A/U 偏好性。RSCU<1 的密碼子共有29 個(CGC、AAC、GAC 等),以G/C 結尾的共有23 個,且以C 結尾占52.13%和以G 結尾的占47.83%(表5)。
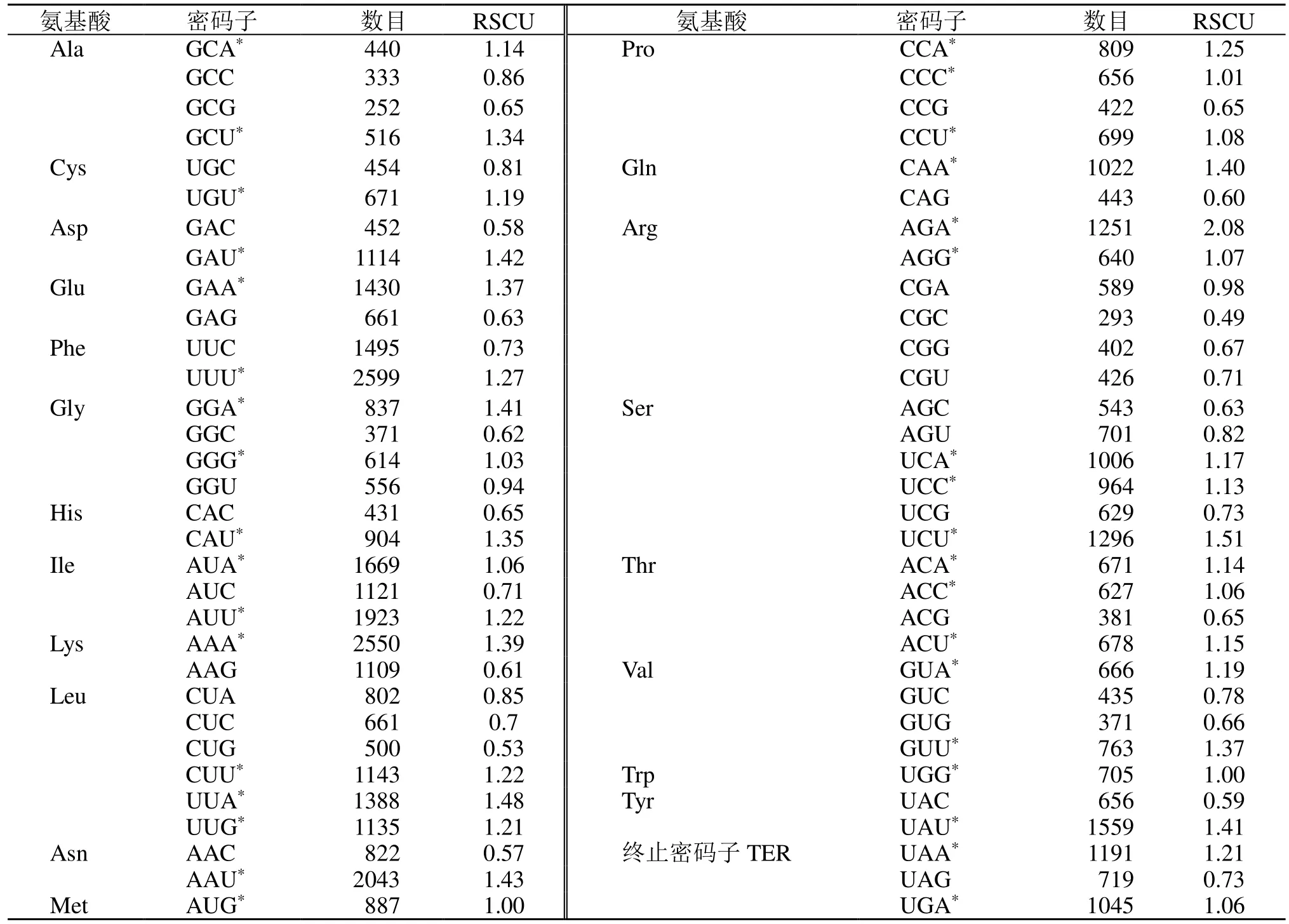
表5 遠志綠體基因組蛋白編碼序列RSCU 分析Table 5 RSCU of protein coding region in chloroplast genome of P. tenuifolia
2.5 基因組特征及IR 邊界比較分析
從NCBI 下載了遠志、瓜子金、卵葉遠志、黃花倒水蓮、黃花遠志、西南遠志、香港遠志和密花遠志8種遠志屬植物的葉綠體基因組,它們的全長分別為165 423、165 439、165 397、164 687、164 947、164 268、165 296、171 893 bp。黃花倒水蓮葉綠體基因組的GC 值最大,為36.9%,其余GC 含量均在36.7%~36.9%(表6)。瓜子金的LSC 最長,為83 722 bp、密花遠志的SSC 最長為8409 bp。

表6 8 種遠志屬植物葉綠體基因組的特征Table 6 Chloroplast genome characteristics of eight Polygala plants
在遠志屬8 種植物的葉綠體基因組邊界區域中,遠志葉綠體基因組共存在4 個邊界,遠志與其他7種植物的LSC/IRb、IRb/SSC、SSC/IRa 和IRa/LSC邊界及基因分布(圖4)。除密花遠志和西南遠志外,JLB(LSC/IRb)邊界兩側分布均有rpsl2和rps19基因,密花遠志邊界位于petB基因中,是其獨有的基因,該基因的保守長度為1390 bp,西南遠志JLB 邊界有rpl23基因存在,距離邊界184 bp。其中遠志中的rps19基因距離JLB(LSC/IRb)邊界長度為1 bp,其位于LSC 區域內,rps12位于IRb 區域內。
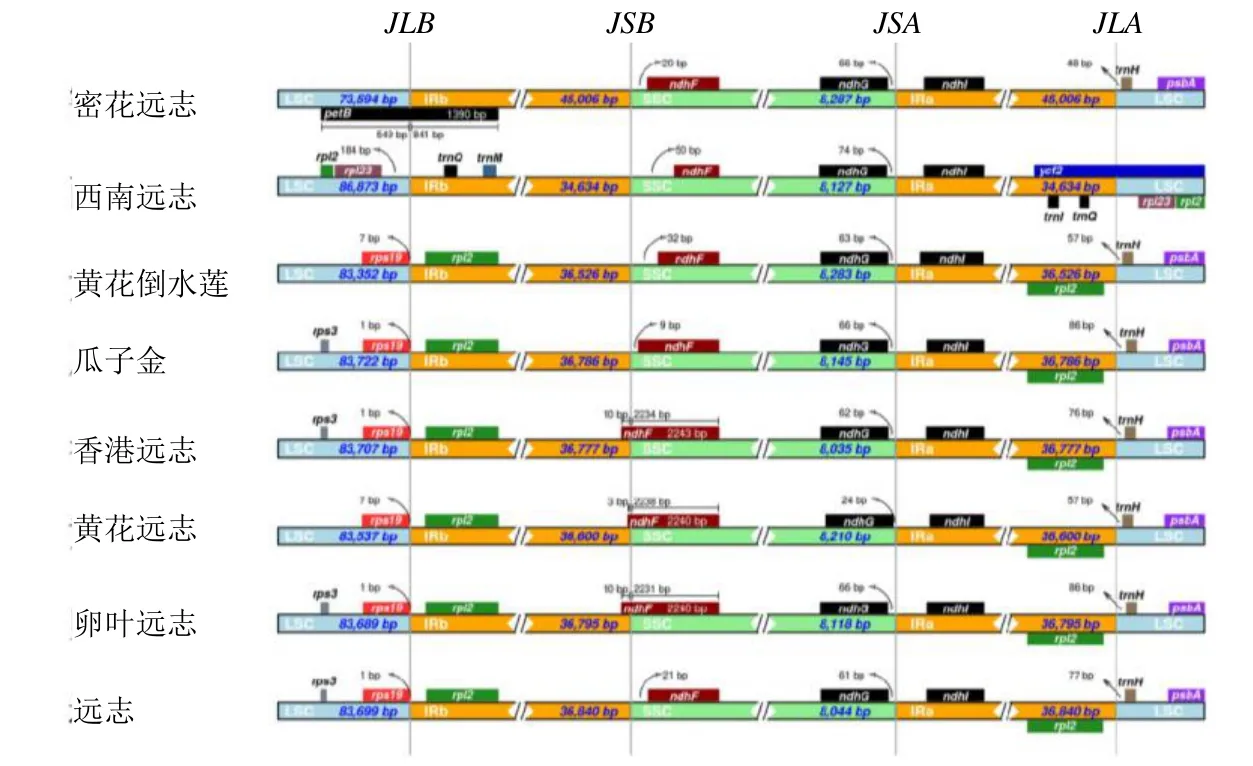
圖4 遠志屬8 個物種葉綠體基因組的IR 與SC 邊界比較Fig. 4 Boubdary comparison of IR and SC region of chloroplast genomes in nine species of Polygala
ndhF基因位于JSB(IRb/SSC)的邊界,8 種植物在該位置與基因存在間隙或重疊存在。其中該基因在遠志中距SSC 區僅21 bp,其中香港遠志和卵葉遠志ndhF擴張最大。香港遠志、黃花遠志和卵葉遠志的JSB(IRb/SSC)邊界均位于ndhF,其余遠志屬物種均在SSC 區域,該基因相對保守長度均為2240 bp。另一個ndhG基因位于JSA(SSC/IR)交界處,均位于SSC 區域中。除了黃花遠志距離邊界24 bp,其余距離邊界60~74 bp。
ndhI基因位于JSB(IRb/SSC)的邊界,均位于IRa 區域中。西南遠志JLA(IRa/LSC)邊界分布有ycf2基因,密花遠志分布有trnH基因。而其他6 種植物在該處均為trnH和rpl2基因。其中,trnH基因均分布在LSC 區域,rpl2基因均分布在IRa 區域。IRA-LSC 邊界均與trnH基因存在著48~86 bp 的間隙。總體來說,遠志屬的西南遠志和密花遠志葉綠體基因組和邊界基因與其余6 個葉綠體基因組相互之間具有明顯的差異。
2.6 基于葉綠體基因組的遠志系統發育分析
利用最大似然法構建系統發育分析結果顯示,11 種植物被分為2 大類,即遠志科和外類群(苦木科和蕓香科),見圖5。包括遠志在內的遠志屬8 個物種和一個齒果草屬以100%的支持率構成一個單系分支與蕓香科和苦木科區分開來。同時,這9 個物種可以進一步劃分為2 個次級單系分支,其中,同來自遠志屬的黃花遠志、黃花倒水蓮、密花遠志和齒果草屬齒果草4 個物種構成一個單系的支持率為100%,齒果草屬和部分遠志屬聚類到一塊值得進一步研究,側面說明葉綠體基因組序列也并不能完全解決遠志屬內部網狀進化等問題,仍需和其他基因組學聯合分析。結合IR 邊界分析可知遠志、瓜子金、卵葉遠志和香港遠志的親緣關系最近。本研究的遠志葉綠體基因組序列可為遠志科后續開展遺傳多樣性研究提供重要信息。

圖5 基于葉綠體全基因組序列構建ML 系統進化樹Fig. 5 Phylogenetic analysis based on chloroplast genome sequences by maximum likelihood (ML) tree
鄰接法的結果與最大似然法構樹除了齒果草和密花遠志位置互換外,其余結果一致,見圖6。遠志科仍分為2 大類同來自遠志屬的遠志和卵葉遠志、瓜子金和香港遠志4 個物種分為一類,黃花倒水蓮、黃花遠志、密花遠志和齒果草屬齒果草聚到一類。仍然可以說明遠志和卵葉遠志、瓜子金和香港遠志關系密切。
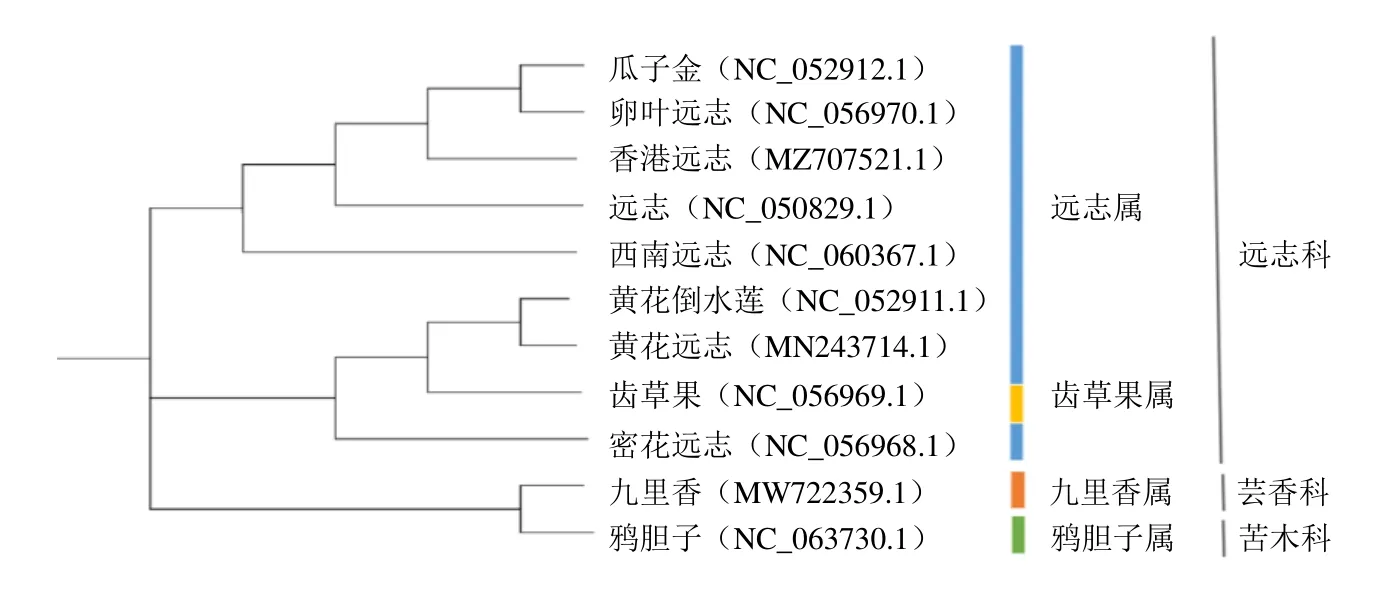
圖6 基于葉綠體全基因組序列構建NJ 系統進化樹Fig. 6 Phylogenetic analysis based on chloroplast genome sequences by neighbor-joining (NJ) tree
3 討論
《中國藥典》2020 年版中將遠志科植物遠志P.tenuifoliaL.的干燥根作為遠志的主流基原藥材[47]進行商品流通,其藥用價值顯著。近年來,面臨遠志的野生資源狀況不容樂觀,植物資源日益貧乏的危險。本實驗基于葉綠體基因組技術進行序列分析、密碼子偏好分析、遠志屬基因組比較分析以及遠志屬植物系統發育關系研究為遠志在野生資源保護、分子育種及遠志屬藥用植物的物種鑒定等領域提供了寶貴的基因資源。
本實驗結果發現,遠志葉綠體基因組長為165 423 bp,GC 含量為36.7%,共編碼135 個基因,這與先前報道的遠志屬其他植物西南遠志[31]的葉綠體基因組結構特征類似,表明遠志的葉綠體基因組結構相對保守。與韓國學者Lee 等[32]發表的文章相比較,本研究將系統發育樹聚焦到遠志科植物上,且增加對其重復序列,密碼子偏好性,遠志屬基因組比較,IR 邊界內容的分析,補充和完善前者文章分析量少的缺點,從而對其葉綠體基因組有了更加全面的了解。
SSR 在植物葉綠體基因組中十分常見,其類型、數目及分布都因植物不同而異,被廣泛應用于植物群體遺傳多樣性和系統發育研究以及分子標記研究等內容[48-49]。本研究通過分析遠志葉綠體基因組,共檢測到161 個SSR 位點,其主要位于LSC 區域;并且單核苷酸A/T 堿基在SSR 位點中出現頻率最高,調研文獻推測原因是在大部分植物中A/T 含量均高于G/C 含量,即A/T 類型的SSR 在植物中最多[37,39,42]。本研究通過對遠志葉綠體基因組中SSR 的數量、組成進行分析,為后續進一步研究分子標記、群體遺傳分析以及作物育種提供參考。
CUB 是基因組中重要的進化特征,RSCU 是作為密碼子偏好性的指標之一[50]。研究表明,自然界存在的20 種氨基酸中,除Met 和Trp 由唯一密碼子編碼外,其他氨基酸均對應2~6 個同義密碼子,由于同義密碼子在機體內的使用頻率存在差異,導致植物密碼子的出現頻率不同,在不同物種翻譯的過程中。存在突變和自然選擇等多種因素,表現出一定的偏好性[51]。本研究中密碼子偏好性分析表明,亮氨酸(Leu)是遠志葉綠體基因組中占比最高的氨基酸,并且RSCU>1 的27 種密碼子均以A/U 結尾,這與其他高等植物相似。
通過對遠志屬基因組特征分析,可知黃花倒水蓮葉綠體基因組的GC 值最大,為36.9%,其余GC 含量均在36.7%~36.9%。瓜子金的LSC 最長為83 722 bp、密花遠志的SSC 最長為8409 bp,通過IR 邊界分析遠志、卵葉遠志、瓜子金、香港遠志、黃花遠志、黃花倒水蓮親緣關系較近,其物種結構更加相似。相對保守的IR 區域的收縮與擴張現象代表著植物的進化,不同植物的葉綠體基因組大小與其密切相關。分析IR-LSC/SSC 區域的邊界信息,對葉綠體基因組結構的差異、物種進化等有進一步了解[52]。通過對8 個物種的IR 邊界研究發現,在密花遠志葉綠體基因組的IR 長度最大,其葉綠體基因組長度8 個物種中也最大,遠志屬植物葉綠體基因組除了西南遠志和密花遠志外,基本邊界變化呈現規律性。
基于遠志科9 個物種的葉綠體基因組數據構建最大似然樹和鄰接法兩種構建系統發育樹,支持率均為100,其進化樹的拓撲結構略有不同,與Ma 等[31]構建的進化樹存在一些差異,分析其系統發育關系可知,遠志、卵葉遠志、瓜子金和香港遠志為姊妹類群,表明四者親緣關系最近,另外遠志科與外類群其他屬能夠很好的區分。原因首先基于葉綠體不同數據集構建的進化樹,本研究相比其數據量更為豐富些。其次葉綠體基因片段可能會丟失某些重要的信息,難以解決物種多、分類較難的大科的系統進化問題。并且基于葉綠體的不同數據集構建的進化樹相比于其他方法構建的進化樹支持率更高,可靠性更強,為遠志的系統分類地位和種間進化關系研究奠定一定的理論基礎。
本研究對遠志葉綠體基因組進行研究,并分析了其葉綠體基因組結構特征,挖掘其葉綠體基因組的重復序列位點、分析其密碼子偏好性和遠志屬葉綠體基因組IR 與SC 邊界分析并利用葉綠體基因組數據構建系統發育樹,有助于后續分子標記、DNA條形碼技術等研究的深入進展,提供了遠志葉綠體基因組信息支持。揭示遠志屬物種之間的系統關系,為藥用遠志的資源篩選、鑒定、保存及遺傳多樣性分析等后續研究提供了分子依據,為產業化應用奠定基礎,進而為保護遠志物種資源提供有力保障。
利益沖突所有作者均聲明不存在利益沖突
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
電子制作(2018年18期)2018-11-14 01:48:24
汽車觀察(2018年10期)2018-11-06 07:05:26
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業技術(2016年15期)2016-12-01 05:31:22
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
