產腸毒素脆弱擬桿菌在結直腸癌中致病機制和治療策略研究進展
2023-06-07 06:39:32王芳芳黃亞輝王孝清盛春泉劉娜海軍軍醫大學上海00433福建中醫藥大學福州350
中南藥學 2023年5期
王芳芳,黃亞輝,王孝清,盛春泉,劉娜*(. 海軍軍醫大學,上海 00433;. 福建中醫藥大學,福州 350)
據統計,全球每年約有90萬人死于結直腸癌(colorectal cancer,CRC),CRC已成為全球第三大常見惡性腫瘤和第二大致命癌癥[1]。雖然科研人員已經進行了廣泛的研究,但CRC的致病機制仍未完全闡明。由于高通量測序等科學技術的進步,腸道微生物群失衡被認為和CRC的發病機制相關[2]。微生物群是指存在于特定環境中的微生物的集合,包括細菌、古細菌、病毒(噬菌體和真核病毒)、真菌、單細胞原生動物和蠕蟲[3]。人體健康雖然與我們自身的基因組密切相關,但是很大程度上依然與生活在我們體內的微生物有關[4]。在人體中,胃腸道是微生物最易富集的部位,大約含有3×1013個細菌,其中大多數是共生菌[5]。腸道微生物通常分為有益微生物和有害微生物:有益微生物在腸道內為人體提供營養物質和預防疾病發生,而有害微生物在腸道內定植會產生毒素和致癌物質[6]。腸道內細菌主要由擬桿菌門、厚壁菌門、放線菌門、變形菌門和瘤胃球菌科等組成,此外還包括少量的梭狀芽孢桿菌[7]。腸道內定植的細菌中,擬桿菌門的產腸毒素脆弱擬桿菌尤其引人關注且被發現參與CRC的發生發展。
1 脆弱擬桿菌(Bacteroides fragilis,B. fragilis)
擬桿菌門為無芽孢、專性厭氧的革蘭氏陰性菌。擬桿菌門種類繁多,超過20多種,這些細菌在人體腸道中通常扮演健康菌群的角色,維持著腸道菌群平衡。脆弱擬桿菌隸屬于擬桿菌門,屬于條件致病菌,分為非產腸毒素脆弱擬桿菌(nontoxigenicBacteroides fragilis,NTBF)和產腸毒素脆弱擬桿菌 (enterotoxigenicBacteroides fragilis,ETBF)[8]。NTBF與ETBF的不同之處在于是否能夠產生脆弱擬桿菌毒素(Bacteroides fragilisenterotoxin,BFT)。BFT是ETBF的毒力因子,共有BFT-1、BFT-2 和BFT-3 三種基因型。BFT是一種20 kDa鋅依賴型金屬蛋白酶,也被稱為Fragilysin[9]。BFT作為一種無活性酶原,由一個N前端結構域、一個柔性連接體和一個C端催化結構域組成[10],通過一種被稱為fragipain(Fpn)的肽酶裂解其柔性連接蛋白中的精氨酸位點而被激活[10-12]。Fpn是一種C11家族半胱氨酸蛋白酶,通過去除BFT前端結構域而直接激活脆弱擬桿菌細胞中的BFT。BFT通過破壞細胞間緊密連接和增加腸黏膜通透性而導致腸道炎癥和組織損傷[9],進一步研究發現,腸道炎癥和組織損傷會促進CRC的形成[13]。
2 ETBF促進CRC發生的作用機制
ETBF可通過以下幾種途徑促進CRC的發生:① 誘導炎癥反應;② 降解E-鈣黏蛋白(E-cadherin)刺激細胞增殖;③ 激活轉錄激活因子3(STAT3)通路;④ 促進調節性T細胞(Treg)和輔助性T細胞17(Th17)累積;⑤ 參與CRC相關的多種信號通路;⑥ 形成生物膜(見圖1)。
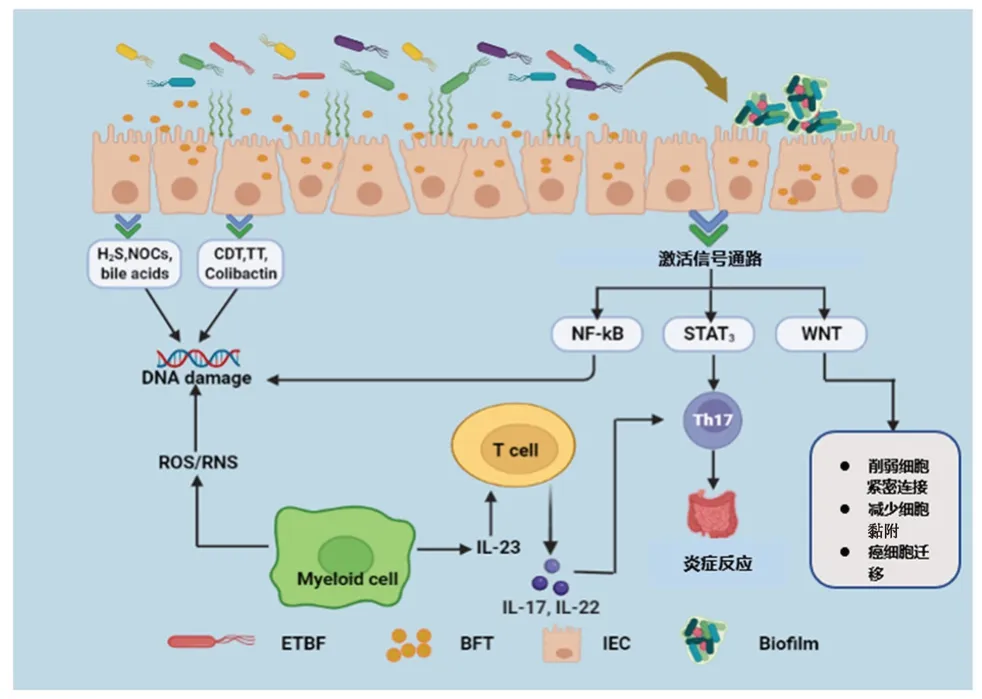
圖1 ETBF與結直腸癌相關作用機制圖Fig 1 ETBF associated mechanism in colorectal cancer
2.1 ETBF誘導炎癥反應
正常情況下,腸道黏膜屏障由一層緊密連接的腸上皮細胞組成并將腸道菌群與免疫細胞隔離開來[14]。在人體和CRC小鼠模型中,腸黏膜屏障具有高滲透性[15]。ETBF產生的BFT,會降解結腸上皮細胞上的E-cadherin,從而破壞結腸黏膜屏障[16]。ETBF還通過分泌顆粒刺激結腸上皮細胞產生外泌體樣納米顆粒,這些顆粒含有高水平的C-C類趨化因子20和前列腺素E2,是Th17細胞募集和IL-17信號增殖轉化所必需的[17]。ETBF還會與宿主免疫系統相互作用,進而發展為慢性炎癥,導致腫瘤發生[18]。導致CRC發展的主要環境風險因素之一是慢性腸道炎癥,慢性炎癥會改變細胞微環境,促使基因突變,抑制細胞凋亡,誘導血管新生和細胞增殖產生癌前病變,最終導致癌癥[19]。在腫瘤發展過程中,腫瘤細胞和ETBF相互作用,產生基因和炎癥免疫因子,這些因子影響腫瘤細胞的存活和增殖[20]。
2.2 ETBF通過降解E-cadherin刺激細胞增殖
除了炎癥反應,BFT還會通過降解E-cadherin改變結腸上皮細胞的結構和功能[21]。E-cadherin是一種120 kDa跨膜糖蛋白,是黏著帶的主要結構蛋白,也被認為是腫瘤抑制因子黏著帶蛋白。正常情況下,E-cadherin的表達與細胞凋亡以及同型細胞之間黏附等細胞功能相關[22]。然而,當E-cadherin與腸上皮細胞中的BFT相互作用時,它以一種不依賴三磷酸腺苷(ATP)的方式迅速降解E-cadherin。這種降解會導致結腸損傷加重,進而導致細胞形態學改變和細胞轉移能力增強。同時,E-cadherin的降解也促進了β-連環蛋白(β-catenin)和依賴T細胞因子的轉錄激活因子的核定位結合[23-25],這種結合促進了基因調控與轉錄。此外,β-catenin在WNT信號通路中通過促進細胞增殖和上皮-間充質轉化,在增強原癌基因表達中發揮重要作用[21,26]。
2.3 ETBF激活STAT3通路
ETBF引發的慢性炎癥導致基因改變直接影響STAT3通路并促進癌變[27]。ETBF能夠在結腸上皮細胞和結腸細胞中通過磷酸化和核易位迅速激活黏膜免疫細胞中的STAT3。而要激活免疫細胞中的STAT3,上皮細胞要響應產生的細胞因子,如白細胞介素(IL)-6、IL-10和IL-23。除了細胞因子,生長因子如血管內皮生長因子(VEGF)和堿性成纖維細胞生長因子(FGF-2)也參與了STAT3的激活。當ETBF與結腸上皮細胞相互作用時,它們將刺激結腸黏膜免疫細胞中STAT3的早期激活。這種STAT3激活持續緩慢上升,直到達到峰值水平,此時意味著ETBF激活了免疫系統[28]。在ETBF誘發的結腸炎中,由于激活了結腸黏膜中的STAT3通路和Th17細胞免疫應答,因此誘發了原癌性炎癥反應并增加了腸黏膜的滲透性[28]。雖然STAT3激活是長期的,可持續數月,但它會極大地增加慢性炎癥引起腫瘤的概率。此外,STAT3激活會促進腫瘤Treg的積累,并阻止產生抗腫瘤免疫反應,對身體產生不利影響。這種異常持續的STAT3激活增加了腫瘤細胞的耐受性,阻止了免疫系統的排斥反應,降低了免疫治療的有效性,增強了腫瘤發生的可能性[29]。
2.4 ETBF促進Treg和Th17細胞的累積
在正常狀態下,Treg通過高度表達IL-2受體并產生內源性IL-2來抑制IL-17的產生,因而在炎癥反應和腸道免疫調節中發揮重要作用。然而,當ETBF定植在結腸中某個特定的部位時,產生的BFT會激活STAT3通路并誘發結腸炎從而破壞腸道黏膜。同時還會導致Treg與ETBF直接接觸從而促進Treg的激活,激活后的Treg缺乏產生內源性IL-2的能力[30]。一旦Treg不能產生內源性IL-2,它只能通過消耗外源性IL-2來維持生存,Treg對外源性IL-2的消耗降低了外源性IL-2的水平,這就產生了有利于Th17細胞生長的微環境。這表明ETBF在腸道內定植促進了Treg和Th17細胞的累積,Th17細胞開始產生大量的腫瘤壞死因子-α(TNF-α)和IL-17等炎癥細胞因子,這些細胞因子在組織損傷時促進細胞生長和增殖,進而引發慢性炎癥。由致病性Th17細胞產生的IL-17不僅參與組織損傷的早期炎癥反應,而且還會促進腫瘤細胞存活和增殖、腫瘤新生血管形成和轉移,從而導致癌變[31]。同時IL-17可通過IL-6間接激活STAT3通路[28]。由此可見,當ETBF在腸道內定植時,STAT3通路激活與Treg細胞累積都與CRC發生相關。
2.5 ETBF參與CRC多種信號通路(NF-κB、WNT、MAPK)
ETBF產生的BFT參與了結腸上皮細胞的多種信號通路。當BFT激活這些信號通路時,它會給身體帶來不利影響,并可能導致CRC發生。結腸上皮細胞信號轉導可激活NF-κB、WNT和MAPK等多種信號通路[32]。BFT通過激活血紅素加氧酶-1 (HO-1)表達的腸上皮細胞NF-κB通路而導致腸黏膜產生炎癥,該通路可以防止腫瘤細胞凋亡,增強腫瘤細胞存活能力,從而導致腫瘤的發生[33]。此外,當結腸上皮細胞內的NF-κB通路長期被激活時,就會導致一氧化氮合酶活性增強而分解L-精氨酸產生一氧化氮[34]。WNT信號通路是維持結腸上皮細胞結構的重要途徑,因此WNT信號通路對于CRC的發生和進展是極其重要的[35]。隨著WNT信號通路被激活,它會削弱細胞間緊密連接,減少細胞黏附,因而使腫瘤細胞得以遷徙和轉移到其他器官[36]。基質金屬蛋白酶是鋅依賴性蛋白酶,在控制腸道屏障中的基膜及細胞外基質的合成和降解起到重要作用[37]。通過處理非基質生物活性物質來修飾趨化因子或生長因子相關的基質,以及調節其他蛋白酶的活性[38]。基質金屬蛋白酶-7(MMP-7)的分泌與腫瘤轉移和炎癥相關[39]。NF-κB和激活因子-1(AP-1)等轉錄因子可調節MMP-7的表達[40]。有研究發現,BFT刺激結腸上皮細胞可誘導MMP-7激活NF-κB和AP-1等轉錄因子,絲裂原激活蛋白激酶(MAPKs)信號通路對于MMP-7誘導至關重要[41]。
2.6 ETBF形成的生物膜在CRC中的作用
CRC發生的風險因素有很多,包括家族史、年齡、性別、個人病史、吸煙、飲食(紅肉)、肥胖、酗酒和炎癥性腸病等。然而,最近的研究也發現了一種新的危險因素,即細菌生物膜的形成,也被證實與CRC的進展相關[42]。結腸上皮增生的增加導致微生物群落關系的改變,從而改變微生物的組成和活性,進而導致生物膜的形成[43]。生物膜的形成是細菌黏附和生長所必需的,它伴隨著細胞外聚合物和黏附基質的產生而產生,這將導致細菌生長和基因表達的變化。而這些微生物形成的生物膜作為前致癌炎癥反應的觸發器,有可能最終發展成CRC[44]。生物膜會導致結腸上皮細胞E-cadherin的重新分布,增加腸道通透性,導致腸道屏障功能喪失,從而加劇腸道失調,同時生物膜的形成也降低了細菌對輻射和抗菌藥物的敏感性[45]。
與健康人相比,CRC患者結腸組織樣本中產生的生物膜更為常見。有一種理論認為,生物膜承載著不同種類的細菌,而不是入侵微生物的單一變種,并可能導致炎癥反應的增加和具有遺傳毒性的細菌源衍生物的產生。無論是健康個體還是CRC患者,腸道微生物群形成的生物膜都與促癌生物學變化一致,即結腸上皮增生增加,IL-6、STAT3激活增加,多胺增加,E-cadherin減少[46]。而多胺代謝物水平的增加被認為協同促進生物膜的形成和細胞增殖,為結腸細胞的致癌轉化的創造有利條件[46]。此外,改變結腸黏膜的滲透性和細胞代謝也會導致腫瘤微環境的變化。另一方面,在家族性腺瘤性息肉病患者中也發現了生物膜存在,這些患者遺傳了突變的APC基因,作為“腺瘤-癌序列”[47]的早期階段,由于息肉和腺瘤的形成,極易發展成CRC。
3 靶向ETBF藥物治療策略
CRC的常規治療手段包括化療和手術,兩者都有可能導致嚴重的并發癥。手術具有侵入性、死亡率高;化療容易產生耐藥性、細胞毒性和其他不良反應。ETBF被認為是導致CRC癌變的關鍵病原體,而開發靶向這些關鍵病原體的藥物可以減少不良反應或增強現有CRC化療方案的療效,成為一種實用的輔助策略。本文將對幾種作用于ETBF藥物進行簡要回顧,包括抗菌藥物、天然產物、抗炎藥物、益生菌類等。
3.1 抗菌藥物
臨床上用來治療ETBF感染的常用抗菌藥物有青霉素類(如芐青霉素)、頭孢菌素類、喹諾酮類(如諾氟沙星)、克林霉素和甲硝唑等藥物。然而,近年來不斷有抗菌藥物耐藥的相關報道,且耐藥率呈逐步升高趨勢。同時細菌形成的生物膜也會導致其對抗菌藥物的耐藥性增加,迄今為止,還沒有一種抗菌藥物被證明可以有效治療細菌生物膜相關感染。抗菌藥物如多黏菌素、亞胺培南的研究表明,它們不能完全消除生物膜,而只是減少生物膜的產生[48]。最近的研究也表明,廣譜抗菌藥物在包括CRC[8]在內的各種類型癌癥患者中并不能產生良好的臨床效果。生物膜會導致吞噬細胞和補體系統的激活減弱,從而保護致病菌不受宿主免疫系統的影響,使其對常規抗菌藥物的耐藥性增加約1000倍[49]。細菌在生物膜中的代謝狀態和有限的供氧也是導致其對抗菌藥物產生耐藥性的可能因素[50]。Jeyaraj等[51]發現,當給予致死濃度的抗菌藥物時含有生物膜的細胞比浮游細胞的突變率更高,從而增加了抗菌藥物耐藥基因通過質粒轉移的機會。雖然使用抗菌藥物治療可以殺滅ETBF等腸道致病菌,但是由于缺乏靶向性,抗菌藥物的使用會破壞胃腸道生態系統的平衡性和導致耐藥性的發生,使得CRC的臨床治療效果受到限制。
3.2 天然產物
天然產物由于化學結構的多樣性及其已知的生物活性,已成為藥物開發中十分具有潛力的候選藥物。許多天然產物已被證明具有良好的抗菌和抗生物膜活性,最近報道的幾種源自植物的天然產物顯示了其對CRC相關病原體的抗菌作用。
3.2.1 花姜酮(zerumbone) 近年來有研究表明,花姜酮具有抗誘變、抗菌、抗癌變、抗炎等多種生物活性[52]。Kim等[53]研究發現,花姜酮對不同的脆弱擬桿菌菌株,包括野生型ETBF、BFT-2過表達的ETBF和NTBF,均具有抗菌活性。研究證實花姜酮不僅可以抑制生物膜的形成,而且能夠根除已形成的生物膜。同時花姜酮對含有毒性BFT-2基因的脆弱擬桿菌菌株的生物膜形成的抑制作用比非產腸毒素菌株更有效。此外,花姜酮的抗菌膜活性可能是通過下調與生物膜形成相關的外排泵基因(bmeB12)而誘導的[53]。以上結果表明,花姜酮可作為一種抗生物膜和抗菌劑,用于治療和預防ETBF引起的生物膜相關感染,同時還是一種有效的免疫調節劑以及一種具有開發潛力的抗腫瘤藥物。
3.2.2α-葎草烯(α-caryophyllene)α-葎草烯也顯示了抑制ETBF生物膜形成的潛力[54]。α-葎草烯是一種倍半萜,存在于芳香植物包括薄荷、鼠尾草和生姜(姜科)的精油中[55]。有報道稱α-葎草烯精油具有抗炎和抗菌作用[56-57]。與花姜酮類似,α-葎草烯也通過誘導RND型外排泵基因bmeB1和bmeB3的下調來發揮抗菌活性,導致ETBF的細胞膜破壞和生物膜形成的抑制[54]。此外,α-葎草烯在動物模型中表現出顯著的抗炎活性,可減少組胺注射引起的水腫[56]。在卡拉膠注射的大鼠模型中,使用α-葎草烯全身治療可以阻止TNF-α和IL-1β的產生[56]。
3.2.3 小檗堿(berberine) Chen等[58]對小檗堿預防結直腸腺瘤復發的一項多中心、隨機、雙盲研究發現小檗堿可以降低息肉切除術后患者結腸腺瘤和息肉樣病變復發的風險。小檗堿的低成本和安全性展現了它在預防CRC方面的潛力。小檗堿(最初從中草藥黃連中提取)在中國已經使用了幾個世紀,用于治療腹瀉和腸炎。小檗堿的抗腫瘤活性在動物實驗中也有報道,它可以調節腫瘤微環境,阻斷腫瘤發生相關通路的激活,此外,研究也證實小檗堿可以通過改變微生物群的組成來預防小鼠大腸癌的發生。
3.3 抗炎藥物
金諾芬(auranofin)是一種金屬鹽,已獲得美國食品和藥品監督管理局(FDA)批準作為治療類風濕關節炎的藥物。同時它還具有抑制腸道細菌(如ETBF)生長的作用,有可能作為一種抗腫瘤藥物而被重新使用。對類似于金諾芬等已批準藥物的重新使用,可以節省開發和測試新藥所需的時間和費用,同時其安全性也得到了廣泛的驗證[59]。一項研究表明,金諾芬對p53為零的卵巢癌SKOV3細胞系具有抗腫瘤活性[60]。Jang等[59]研究了金諾芬對ETBF的抗菌和抗生物膜作用,證實了用相對較低濃度的金諾芬對ETBF有很好的抑制和根除細菌生物膜作用。經金諾芬治療后,外膜蛋白(OmpA)基因和外排泵相關基因bmeB3的表達顯著降低。后續研究需要進一步在體內模型中驗證金諾芬抑制ETBF的作用機制。
3.4 益生菌
益生菌被認為是無致病性的安全菌株,通過維持腸道微生物平衡而起到促進健康的作用,常被作為活補劑使用[60]。益生菌在治療微生物感染,促進人類健康,并在預防抗菌藥物相關的腹瀉、早產兒壞死性小腸結腸炎,治療嬰兒結腸絞痛、牙周病以及誘導或維持潰瘍性結腸炎緩解等方面表現出良好的應用前景[61]。不同益生菌混合物對產生生物膜的腸菌病原體(包括ETBF和產腸毒素的大腸埃希菌)都有抗菌膜特性[62-63]。Shin等[62]最近研究了益生菌丁酸梭菌NCTC 7423上清液對ETBF基因表達和生物膜形成的影響。丁酸梭菌對ETBF的生長表現出拮抗作用,丁酸梭菌的細菌定植因子(CFs)還抑制了生物膜的成熟,使已形成的生物膜被破壞,降低了生物膜中細胞的代謝活性[62]。
益生菌產生多種拮抗化合物,包括胞外多糖[64]、細菌素[64]和生物表面活性劑[65],這些拮抗化合物可以干擾生物膜的附著和形成,以及分解成熟的生物膜。此外,益生菌能夠改變腸道pH值,從而與致病菌競爭營養物質[66]。益生菌在進入宿主體內后,除與病原體直接相互作用外,還可以通過與免疫系統相互作用發揮免疫調節作用。研究表明,益生菌和其產生的可溶性因子可通過Toll樣受體識別調節和激活特異性免疫細胞,釋放細胞因子,從而產生免疫調節作用[67]。有研究報道聯合補充ω-3脂肪酸和微生物細胞制劑可以改善CRC患者化療的生活質量,減少炎癥生物標志物(如IL-6、TNF-α)水平,緩解化療產生的不良反應[68]。在結腸癌患者腫瘤組織和附近的黏膜中有明顯的微生物群存在,經過益生菌干預后,患者糞便中具核梭桿菌和ETBF含量顯著降低[69]。目前,益生菌的攝入在一些臨床試驗中顯示出了良好的結果,被認為是一種可行的通過調節腸道微生物群來預防CRC的方法。
4 總結和展望
隨著CRC發病率逐年升高,ETBF作為導致CRC發生的關鍵病原體,正成為近年來研究的熱點,而開發靶向這些關鍵病原體的藥物將為CRC的治療開辟新的方向。本文通過對ETBF的致病機制和靶向治療藥物進行簡要回顧,發現研發新的CRC針對性治療藥物迫在眉睫,而開發靶向腸道微生物相關藥物將成為一種潛在的預防和治療CRC的方法。結直腸特殊的生理位置,胃腸道的生理復雜性,結腸生物利用度以及干預的特定部位靶向能力等問題都為開發抗CRC藥物增加了難度。對定植于胃腸道中的具有致病性的ETBF等微生物進行針對性的干預,不僅可以消除特定的致病微生物,同時也可以最大限度地降低對胃腸道生態系統的破壞。開發靶向腸道致病菌相關藥物,將為優化精準治療方案,提高患者生存率提供可能。微生物學和腫瘤學領域未來的挑戰將包括開發快速和經濟有效的腸道微生物群失調診斷方法,以及闡明致病菌對CRC的生物效應和作用機制。
