馬鈴薯響應低溫脅迫的蛋白質(zhì)組學分析
2023-06-04 09:34:37李文翔王艦王芳
西北農(nóng)業(yè)學報 2023年4期
李文翔 王艦 王芳


摘 要 為探究馬鈴薯在蛋白質(zhì)組學水平上對低溫脅迫的響應機制,對耐低溫品種‘DR-2與低溫敏感品種‘費烏瑞它組培苗葉片蛋白質(zhì)組學進行分析。結(jié)果表明,鑒定到肽段282 790個,唯一性肽段19 099個,蛋白質(zhì)4 471個。通過Label-Free技術(shù)對葉片差異表達蛋白分析表明,低溫脅迫下‘DR-2中有38個顯著差異蛋白,‘費烏瑞它中有72個顯著差異蛋白。篩選出代謝通路差異蛋白13個,包括代謝相關(guān)蛋白5個,光合作用相關(guān)蛋白3個,防御相關(guān)蛋白2個,運輸相關(guān)蛋白3個。Pathway富集分析結(jié)果表明低溫脅迫處理下,分布在光合作用、能量代謝通路中的3-磷酸甘油醛脫氫酶、鈣網(wǎng)蛋白、熱激蛋白、肌醇-3-磷酸合酶在葉片中差異表達,該結(jié)果對研究馬鈴薯低溫響應機制具有一定參考價值。
關(guān)鍵詞 馬鈴薯;低溫脅迫;蛋白質(zhì)組學;響應機制
低溫是制約植物生長和發(fā)育最常見的非生物脅迫之一,對植物生長和生產(chǎn)均有不利影響。植物通過細胞到生理應答反應迅速啟動相關(guān)基因進行轉(zhuǎn)錄后調(diào)控,合成相關(guān)蛋白,形成多個調(diào)控網(wǎng)絡以響應低溫脅迫[1]。作為基因表達的產(chǎn)物,蛋白質(zhì)參與植物體內(nèi)大部分生理功能的調(diào)節(jié)[2]。利用非標記定量蛋白質(zhì)組學對低溫脅迫中差異表達蛋白進行研究,可了解低溫脅迫下蛋白質(zhì)的變化,揭示其作用模式,鑒定潛在目標蛋白,從而在蛋白質(zhì)水平上掌握脅迫因子傷害機制與植物適應機制。
利用蛋白質(zhì)組學方法以不同組織作為蛋白質(zhì)表達的研究已在水稻[3]、玉米[4]、小麥[5]、擬南芥[6]等植物中報道。歐文軍[7]在木薯葉片中鑒定出34個差異蛋白通過維持光合作用、能量代謝和分子伴侶等表達以響應低溫脅迫。Rocco等[8]短期低溫處理擬南芥后發(fā)現(xiàn)蛋白質(zhì)數(shù)量增加,主要參與能量產(chǎn)生和碳代謝等途徑。唐秀英等[9]對不同品種水稻低溫處理,發(fā)現(xiàn)苗期根系差異蛋白主要在碳代謝、次級代謝產(chǎn)物合成等途徑中起催化結(jié)合作用。
馬鈴薯作為一種重要的主糧作物,由于適應能力強、增產(chǎn)潛力大等特點,已成為僅次于小麥、水稻的第三大糧食作物[10],在全球范圍廣泛種植。馬鈴薯喜冷涼,但溫度低于10 ℃時對幼苗生長不利,苗期遭受低溫傷害后誘導體內(nèi)可溶性糖和蛋白質(zhì)生成,保護植株免受低溫傷害;同時光合作用受阻,導致植株停止生長,對產(chǎn)量造成影響[11]。國內(nèi)對馬鈴薯蛋白質(zhì)的研究主要集中在干旱脅迫[12]和生長發(fā)育[13]等方面,在低溫脅迫方面的研究較少。本研究以Label-Free技術(shù)為主要研究手段,利用蛋白質(zhì)組學方法挖掘馬鈴薯低溫脅迫響應因子,篩選差異表達蛋白,為后續(xù)基因克隆和低溫響應途徑研究提供參考,也為抗性品種的遺傳改良和培育提供理論依據(jù)。
1 材料與方法
1.1 供試材料
以馬鈴薯(Solanum tuberosum L.)耐低溫品種‘DR-2和低溫敏感品種‘費烏瑞它組培苗為試驗材料,由青海省農(nóng)林科學院生物技術(shù)研究所提供。
1.2 試驗方法
1.2.1 材料處理方法 組培苗在MS培養(yǎng)基上培養(yǎng)15 d后,選取長勢一致的組培苗進行試驗。對照組轉(zhuǎn)入人工氣候培養(yǎng)箱,培養(yǎng)箱溫度白天20 ℃、晚上18 ℃,光照度為10 000 lx,光照時間為12 h;處理組進行低溫脅迫,溫度為4 ℃,其余條件與對照組相同,設置3次重復。分別處理5 d后,每個重復取18株,將完整功能葉混樣后液氮冷凍放入10 mL離心管,備用。
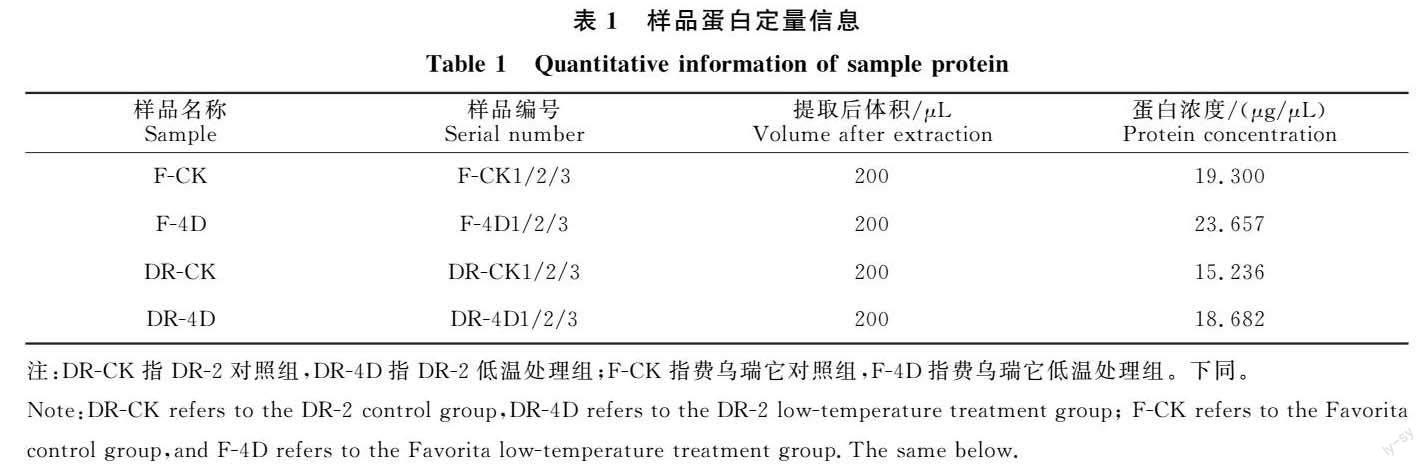
1.2.2 樣品制備 用研缽液氮冷凍研磨樣品,使用甲醇-氯仿沉淀法沉淀蛋白質(zhì),使用NanoDrop 進行蛋白定量,定量信息見表1。用FASP對全部蛋白質(zhì)進行酶解:每例樣品中分別加入適量DTT至終濃度為100 mmol/L,沸水浴5 min后室溫冷卻。轉(zhuǎn)入10? ku超濾離心管去除分子量大于10 ku的大分子蛋白,12 000×g離心15 min,棄濾液。加入100 μL IAA,震蕩混勻,室溫避光放置30 min,12 000× g離心10 min,棄濾液。加入100 μL NH4HCO3,震蕩混勻,14 000×g離心10 min,重復4次后加入40 μL Trypsin,充分振蕩,37 ℃水浴16~18 h。換新收集管,12 000×g離心10 min,收集濾液,加入適量0.1% TFA溶液,酶解后的肽段使用C18 Cartridge對上清液進行脫鹽處理,真空干燥。酶解后的肽段用0.1% FA復溶,取上清進行濃度測定,以備LC-MS/MS分析。
1.2.3 LC-MS/MS分析 每例樣品取適量肽段使用納升流速Easy nLC 1 200 色譜系統(tǒng)進行色譜分離。緩沖液:A液為 0.1%甲酸水溶液,B液為0.1%甲酸乙腈水溶液(乙腈為 80%)。色譜柱以95%的A液平衡。樣品進樣到Trap Column(100 μm×20 mm,5 μm,C18,Dr. Maisch GmbH)后經(jīng)過色譜分析柱(75 μm×150 mm,3 μm,C18,Dr. Maisch GmbH)進行梯度分離,流速為300 nL/min。液相梯度設置如下:0~3 min,B液線性梯度為2%~8%;3~98 min,B液線性梯度為8%~28%;98~108 min,B液線性梯度為28%~40%;108~110 min,B液線性梯度為40%~100%;110~120 min,B液維持在100%。肽段分離后用Q-Exactive HF-X質(zhì)譜儀進行數(shù)據(jù)依賴采集質(zhì)譜分析。分析時長為120 min,檢測模式:正離子,母離子掃描范圍:300~1 800 m/z,一級質(zhì)譜分辨率:60 000 @m/z 200,AGC target:3e6,一級Maximum IT:50 ms。肽段二級質(zhì)譜分析按照下列方法采集:每次全掃描后觸發(fā)采集20個最高強度母離子的二級質(zhì)譜圖譜(MS2 scan),二級質(zhì)譜分辨率:15 000 @ m/z 200,AGC target:1e5,二級Maximum IT:25 ms,MS2 Activation Type:HCD,Isolation window:1.6 m/z,Normalized collision energy:28。Label-free非標記定量分析委托上海拜譜生物科技有限公司進行。
1.2.4 蛋白質(zhì)鑒定與數(shù)據(jù)庫檢索 采用檢索軟件MaxQuant 1.6.0.16進行質(zhì)譜數(shù)據(jù)檢索,參數(shù)設置見表2。
1.2.5 蛋白質(zhì)定量與差異分析 MaxQuant是領先的蛋白質(zhì)組學定性定量算法,已逐漸成為該領域內(nèi)的標準解決方案之一[14]。在定量結(jié)果的顯著性差異分析中,本試驗篩選樣本組內(nèi)3次重復數(shù)據(jù)進行統(tǒng)計分析,視上下調(diào)差異倍數(shù)大于2且P小于0.05的蛋白質(zhì)為顯著差異表達蛋白質(zhì)。采用兩組樣本間的蛋白質(zhì)表達差異倍數(shù)和T檢驗得到的P兩個因素共同繪制火山圖,用于表現(xiàn)兩組樣本數(shù)據(jù)的顯著性差異。
1.2.6 生物信息學分析 GO是一個標準化基因功能分類體系,從生物過程、分子功能和細胞組分三方面對目標蛋白質(zhì)進行分類[15],GO注釋的顯著性富集分析通過Fisher精確檢驗來評價。KEGG通路富集分析以所有定性蛋白質(zhì)為背景,通過Fisher精確檢驗,來分析計算各個通路蛋白質(zhì)富集度的顯著性水平,從而確定受到顯著影響的代謝和信號轉(zhuǎn)導途徑[16-17]。
2 結(jié)果與分析
2.1 蛋白質(zhì)鑒定結(jié)果統(tǒng)計
對蛋白質(zhì)鑒定結(jié)果統(tǒng)計,其中馬鈴薯品種‘DR-2對照組中鑒定肽段共計69 666個,唯一性肽段15 635個,蛋白組數(shù)目4 284個;低溫處理組中鑒定肽段共計66 517個,唯一性肽段15 263個,蛋白組數(shù)目4 224個。馬鈴薯品種‘費烏瑞它對照組中鑒定肽段共計71 051個,唯一性肽段15 931個,蛋白組數(shù)目4 206個;低溫處理組中鑒定肽段共計75 556個,唯一性肽段16 527個,蛋白組數(shù)目4 299個。本試驗得到鑒定肽段共計282 790個,唯一性肽段共計19 099個,蛋白組數(shù)目共計4 471個。
2.2 蛋白質(zhì)定量結(jié)果統(tǒng)計
利用DAA質(zhì)譜數(shù)據(jù),分析‘DR-2低溫處理組與對照組、‘費烏瑞它低溫處理組與對照組、‘DR-2低溫處理組與‘費烏瑞它低溫處理組和‘DR-2對照組與‘費烏瑞它對照組共4個對照組中的顯著性差異表達蛋白質(zhì),分別篩選出顯著差異蛋白總數(shù)為38、72、195和193,其中上調(diào)表達蛋白與下調(diào)表達蛋白結(jié)果見圖1。
2.3 顯著性差異蛋白質(zhì)分析
紅色點為上調(diào)表達蛋白質(zhì),綠色點為下調(diào)表達蛋白質(zhì),灰色點為非顯著性差異蛋白質(zhì)。蛋白質(zhì)點的橫縱坐標為離散度來表明蛋白質(zhì)差異程度,離散度越大差異越大。各組間顯著性差異蛋白見圖2。
2.4 蛋白質(zhì)聚類分析
本試驗從樣本和蛋白質(zhì)定量信息兩個維度進行分類,紅色代表上調(diào),藍色代表下調(diào)。由圖3可以看出低溫脅迫和對照組的葉片差異蛋白上下調(diào)表達情況,每個處理的3組重復中相似性很高,說明本試驗所篩選的目標差異蛋白質(zhì)較合理。
2.5 GO富集分析
分析發(fā)現(xiàn)‘DR-2在低溫處理與對照組的差異蛋白表現(xiàn)在33個生物過程、9個細胞組分、25個分子功能途徑中。根據(jù)GO層級水平關(guān)系及富集程度篩選出主要參與低溫脅迫應答的18個途徑。由表3可知生物過程功能分類中,差異表達基因在單一生物合成及小分子代謝過程中富集度較高;L-抗壞血酸代謝過程只有一個上調(diào)表達基因。細胞組分功能分類中,細胞質(zhì)中差異表達基因顯著富集。分子功能分類中,催化活性GO term極顯著富集,且占比最多,共22個基因富集;焦磷酸酶活性過程有5個基因富集,3-磷酸肌醇合酶活性占比較少,只有1個基因表達。GO term富集說明‘DR-2葉片響應低溫脅迫的主要組分在細胞質(zhì)中,通過激活焦磷酸酶、水解酶及3-磷酸肌醇合酶等活性催化更多蛋白質(zhì)合成,參與有機酸的合成及小分子代謝、單糖代謝等過程來抵御低溫脅迫。
分析發(fā)現(xiàn)‘費烏瑞它在低溫處理與對照組的差異蛋白表現(xiàn)在61個生物過程、34個細胞組分、34個分子功能途徑中。根據(jù)GO層級水平關(guān)系及富集程度篩選出主要參與低溫脅迫應答的18個途徑。由表4可知生物過程功能分類中,雖然有機物生物合成過程差異表達基因顯著富集,但是P值相對較高;雙糖、葡萄糖代謝以及對非生物刺激、光刺激和含氧化合物等反應表現(xiàn)出抑制下調(diào)狀態(tài)。細胞組分中主要為細胞內(nèi)部分。分子功能分類中,結(jié)構(gòu)分子活性、熱激蛋白結(jié)合、3-磷酸肌醇合酶活性等極顯著差異,在氧化還原酶活性中顯著富集。GO term富集說明‘費烏瑞它的葉片對氧化還原酶活性響應較強,可能是通過光系統(tǒng)Ⅱ產(chǎn)氧復合物促進熱激蛋白的結(jié)合降低了雙糖和葡萄糖反應速率,進而引起低溫刺激應答基因的下調(diào)表達。
分析發(fā)現(xiàn)‘費烏瑞它低溫處理與‘DR-2低溫處理的差異蛋白表現(xiàn)在175個生物過程、36個細胞組分、91個分子功能途徑中。圖4表示低溫處理下,差異蛋白在3個類別中前10的顯著富集GO term。生物過程功能分類中,小分子代謝、單一生物代謝過程等顯著富集。細胞組分功能分類中,細胞質(zhì)部分及葉綠體部分顯著富集。分子功能分類中,氧化還原酶活性、催化活性等顯著富集。GO term富集說明在低溫脅迫下造成‘費烏瑞它與‘DR-2不同抗性的原因可能是由于不同馬鈴薯品種在葉片細胞質(zhì)內(nèi)與葉綠體類囊體、葉綠體膜等光合作用相關(guān)的細胞組分結(jié)合能力不同,通過催化氧化還原酶活性、水解酶活性、熱激蛋白結(jié)合等,造成對糖代謝、有機物合成、非生物刺激等途徑的不同反應導致對低溫的不同抗性。
分析發(fā)現(xiàn)‘費烏瑞它對照組與‘DR-2對照組的差異蛋白表現(xiàn)在155個生物過程、30個細胞組分、65個分子功能途徑中。圖5顯示‘費烏瑞它和‘DR-2對照組中,差異蛋白在3個類別中前10的顯著富集GO term。
2.6 KEGG通路富集分析
通過KEGG通路富集分析(圖6),4個差異蛋白在氨基酸生物合成和亞油酸代謝2個生物代謝途徑中發(fā)生了顯著性變化(P<0.05)。
如圖7所示,13個差異蛋白顯著富集在3個生物代謝途徑中,其中光合作用、半膀氨酸和蛋氨酸代謝2個途徑發(fā)生了極顯著變化(P<0.01),氨基酸生物合成途徑發(fā)生了顯著性變化(P<0.05)。
DR-4D vs DR-CK比較組鑒定出相關(guān)差異蛋白4個,F(xiàn)-4D vs F-CK比較組鑒定出相關(guān)差異蛋白12個,相關(guān)差異蛋白鑒定信息見表5。
如圖8所示,78個差異蛋白顯著富集在9個生物代謝途徑中,其中內(nèi)質(zhì)網(wǎng)中蛋白質(zhì)加工、卟啉和葉綠素代謝、氨基酸生物合成、次生代謝產(chǎn)物的生物合成、苯丙氨酸酪氨酸和色氨酸的生物合成、谷胱甘肽代謝6個途徑發(fā)生了極顯著變化(P<0.01),精氨酸生物合成、光合生物中的碳固定、丙氨酸天冬氨酸和谷氨酸代謝3個途徑發(fā)生了顯著性變化(P<0.05)。
如圖9所示,71個差異蛋白顯著富集在10個生物代謝途徑中,其中內(nèi)質(zhì)網(wǎng)中蛋白質(zhì)加工、光合作用、光合作用-觸角蛋白、精氨酸生物合成、光合生物中的碳固定5個途徑發(fā)生了極顯著變化(P<0.01),丙氨酸天冬氨酸和谷氨酸代謝、谷胱甘肽代謝、代謝途徑、氨基酸生物合成、牛黃酸代謝5個途徑發(fā)生了顯著性變化(P<0.05)。
F-4D vs DR-4D比較組鑒定出相關(guān)差異蛋白33個,F(xiàn)-CK vs DR-CK比較組鑒定出相關(guān)差異蛋白34個,相關(guān)差異蛋白鑒定信息見表6。
3 討論
3.1 能量代謝相關(guān)蛋白
3-磷酸甘油醛脫氫酶(GAPDH)是植物體中將葡萄糖及糖原分解為丙酮酸過程的一種關(guān)鍵酶,催化3-磷酸甘油醛氧化脫氫和磷酸化生成的糖酵解途徑的中間產(chǎn)物,并使得脫氫酶的輔酶NAD+生成NADH和H+,通過蘋果酸等進入線粒體,經(jīng)過氧化呼吸鏈的傳遞最終生成水和二氧化碳[18]。KEGG富集分析表明,對低溫處理與對照組分析發(fā)現(xiàn)3-磷酸甘油醛脫氫酶在‘DR-2中上調(diào)表達,在‘費烏瑞它中下調(diào)表達;在低溫處理下的‘費烏瑞它和‘DR-2中發(fā)現(xiàn)NAD脫氫酶和蘋果酸酶下調(diào)表達,與GO富集分析結(jié)果葡萄糖代謝生物過程抑制下調(diào)表達結(jié)果一致。表明在低溫脅迫下,由于馬鈴薯品種不同對3-磷酸甘油醛脫氫酶的表達不同,導致對葡萄糖的分解能力不同,推測是造成‘DR-2耐低溫性高于‘費烏瑞它的原因之一。
低溫導致植物中滲透調(diào)節(jié)物質(zhì)、酶活性和細胞結(jié)構(gòu)等改變[19],誘導植物體基因的上下調(diào)表達,進而產(chǎn)生冷脅迫相關(guān)特異蛋白[1]。肌醇-3-磷酸合酶(MIPS)作為一種磷蛋白,參與肌醇合成過程,在肌醇穩(wěn)態(tài)中起著重要的作用。MIPS的磷酸化是生物肌醇合成的新型調(diào)節(jié)機制[20]。肌醇通過自身氧化代謝生成參與細胞壁合成的木聚糖及果膠[21],促進細胞對養(yǎng)分的吸收利用,穩(wěn)定植株胞內(nèi)滲透壓以提高對低溫的耐受性。研究表明MIPS在擬南芥[22]、煙草[23]、水稻[24]中過表達增強了其對鹽、干旱和低溫脅迫的抗性; OsMIPS1的過表達增加了低溫抗性,沉默表達則降低了水稻低溫抗性[25]。植物細胞存在類三磷酸肌醇受體蛋白,參與磷脂酰肌醇系統(tǒng)信號轉(zhuǎn)導[26],通過對細胞膜液泡膜等結(jié)合調(diào)控,調(diào)節(jié)細胞分裂分化,推測本研究中低溫處理下的‘費烏瑞它和‘DR-2肌醇磷酸合酶的上調(diào)表達穩(wěn)固了植株胞內(nèi)外環(huán)境,提高了低溫抗性。
3.2 光合作用相關(guān)蛋白
熱激蛋白(HSP)是植物細胞受逆境脅迫產(chǎn)生的一種維持生命活動的功能蛋白,熱激70 ku蛋白上調(diào)表達可提高植株的抗逆性[27]。HSP70B的低表達可提高光系統(tǒng)Ⅱ的光敏感性,HSP70B的過表達具有保護作用[28]。GO富集與KEGG富集分析表明,熱激70 ku蛋白的抑制下調(diào)可能是造成‘費烏瑞它中光系統(tǒng)Ⅱ產(chǎn)氧復合物及光合作用相關(guān)生物途徑上調(diào)表達的原因。低溫通過降低代謝酶的活性,最終影響光合效率。植物光合作用電子傳遞鏈在低溫脅迫抑制后活性氧含量降低[29]。本研究發(fā)現(xiàn)在低溫脅迫下‘費烏瑞它含氧化合物反應處于抑制下調(diào)狀態(tài),但是熱激同源70 ku蛋白、psaC多肽蛋白、Psb28亞基蛋白上調(diào)表達,這可能是相關(guān)HSP與基因蛋白共同作用在一定程度上保護了低溫環(huán)境下的‘費烏瑞它植株避免凍傷致死。
3.3 其他蛋白
鈣網(wǎng)蛋白(CRT)是一種植物胞內(nèi)分子伴侶,參與鈣離子穩(wěn)態(tài)過程的保守功能蛋白[30]。Komatsu等[31]研究表明CRT基因的過表達提高了水稻耐寒性。Sharma等[32]試驗表明CRT基因可以響應水稻低溫脅迫。本研究發(fā)現(xiàn)低溫處理下的‘費烏瑞它中鈣網(wǎng)蛋白下調(diào)表達,可能降低了植株胞內(nèi)穩(wěn)態(tài),造成‘費烏瑞它低溫抗性較‘DR-2弱,這與前人的研究一致。
ACO2是一種氧化酶基因,在木瓜中證實與果實成熟期的葉片衰老相關(guān)[33]。本研究中 ACO2基因上調(diào)表達,推測是造成葉片胞內(nèi)活性氧積累,導致‘費烏瑞它低溫敏感的原因之一。LOX是一種脂氧合酶,吳錦程等[34]表明低溫脅迫下鈣離子系統(tǒng)與膜脂氧合酶活性調(diào)控相關(guān),廖晶晶[35]發(fā)現(xiàn)LOX10提高了薄皮甜瓜植株的耐旱性,本研究中LOX1.5在‘DR-2中上調(diào)表達,推測是穩(wěn)固了膜系統(tǒng)結(jié)構(gòu),提高了低溫耐受性。
4 結(jié)? 論
不同馬鈴薯品種在生長發(fā)育過程中蛋白質(zhì)的表達存在差異,試驗結(jié)果表明:分布在光合作用、能量代謝等通路中相關(guān)蛋白的差異表達是造成馬鈴薯對低溫不同抗性的主要原因。推測3-磷酸甘油醛脫氫酶、鈣網(wǎng)蛋白、熱激蛋白、肌醇-3-磷酸合酶在低溫脅迫響應途徑中十分重要,在后續(xù)試驗中可對其進行驗證。
參考文獻 Reference:
[1] 楊慧菊,蘭玉倩,王石華.植物響應低溫脅迫組學研究進展[J].山東農(nóng)業(yè)科學,2020,52(5):142-148.
YANG H J,LAN Y Q,WANG SH H.Advances in plant response to low temperature stress [J].Shandong Agricultural Science,2020,52(5):142-148.
[2] LEE H E,SHIN D,SANG R P,et al. Ethylene responsive element binding protein 1 (StEREBP1) from? Solanum tuberosum increases tolerance to abiotic stress in transgenic potato plants[J].Biochemical & Biophysical Research Communications,2007,353(4):863-868.
[3] VO K,RAHMAN M M.Proteomics and metabolomics studies on the biotic stress responses of rice:an? update[J].Rice,2021,14(1):1-16.
[4] 石 江,趙 琳,朱月清,等.玉米幼苗葉片響應熱脅迫的蛋白質(zhì)組學分析[J].浙江農(nóng)業(yè)學報,2018,30(6):893-908.
SHI J,ZHAO L,ZHU Y Q,et al.Proteomic analysis of maize seedling leaves in response to heat stress [J].Zhejiang Agricultural Journal,2018,30(6):893-908.
[5] MICHALETTI A,NAGHAVI M R,TOORCHI M,et al. Metabolomics and proteomics reveal drought-stress responses of leaf tissues from spring-wheat[J].Scientific Reports,2018,8(1):5710-5728.
[6] 席景會.低溫脅迫下擬南芥差異蛋白質(zhì)組學研究[D].吉林:吉林大學,2007.
XI J H.Study on differential proteomics of Arabidopsis? under low temperature stress [D]. Jinlin:Jilin University,2007.
[7] 歐文軍.木薯葉片響應低溫脅迫的轉(zhuǎn)錄組和蛋白質(zhì)組研究[D].海口:海南大學,2014.
OU W J. Transcriptome and? proteome study of cassava? leaves in? response to? low? temperature? stress[D]. Haikou:Hainan Unitersity,2014.
[8] ROCCO M,ARENA S,RENZONE G,et al. Proteomic?analysis of temperature stress-responsive proteins in Arabidopsis thaliana rosette leaves[J].Molecular? Biology? Systems,2013,9(6):1257-1267.
[9] 唐秀英,王會民,龍起樟,等.低溫脅迫對水稻苗期根系影響的蛋白質(zhì)組學研究[J].華北農(nóng)學報,2019,34(6):82-88.
TANG X? Y,WANG H? M,LONG Q ZH,et al. Proteomics study on the effect of? low temperature stress on the root system of rice seedling[J].Acta Agriculturae Boreali-Sinica,2019,34(6):82-88.
[10] DEVAUX,ANDR K,PETER O. Potatoes for sustainable global? food? security[J].Potato Research,2014,57(3/4):185-199.
[11] 楊超英.馬鈴薯種質(zhì)資源的耐凍性鑒定和耐凍機理研究[D].西寧:青海大學,2014.
YANG CH Y.Study on the freezing tolerance identification and? freezing tolerance? mechanism of? potato? germplasm? resources[D].Xining:Qinghai University,2014.
[12] 張鳳軍,葉景秀,師 理,等.干旱脅迫下不同抗旱水平馬鈴薯葉片蛋白質(zhì)組學分析[J].江蘇農(nóng)業(yè)科學,2018, 46(9):23-28.
ZHANG F J,YE J X,SHI L,et al.Proteomic analysis of potato leaves with different drought resistance levels under drought stress [J].Jiangsu Agricultural Science,2018,46(9):23-28.
[13] REMPELOS L,COOPER J ,WILCOCKSON S,et al. Quantitative proteomics to study the response of potato to contrasting fertilisation regimes[J].Molecular Breeding,2013,31(2):363-378.
[14] 王 丹. SH-SY5Y細胞BPDE蛋白加合物研究[D].太原:山西醫(yī)科大學,2019.
WANG D.Study on BPDE protein adducts in SH-SY5Y cells [D]. Taiyuan:Shanxi Medical University,2019.
[15] ASHBURNER M,BALL C? A. Gene ontology:tool for the unification of biology.The gene ontology consortium[J].Nature? Genetics,2000,25(1):25-29.
[16] 蔡英杰.轉(zhuǎn)ClNAC9基因露地菊的耐鹽堿性研究[D].哈爾濱:東北林業(yè)大學,2016.
CAI Y J.Study on salt and alkaline tolerance of ClNAC9 transgenic chrysanthemum [D]. Harbin:Northeast Forestry University,2016.
[17] KANEHISA M,GOTO S. KEGG for integration and interpretation of large-scale molecular data sets[J].Nucleic Acids Research,2012,40(4):109-114.
[18] 孫永梅,劉麗杰,馮明芳,等.植物在低溫脅迫下的糖代謝研究進展[J].東北農(nóng)業(yè)大學學報,2015,46(7):95-102,108.
SUN Y M,LIU L J,F(xiàn)ENG M F,et al.Research progress of plant glucose metabolism under low temperature stress [J].Journal of Northeast Agricultural University,2015,46(7):95-102,108.
[19] 陳思琪,孫敬爽,麻文俊,等.植物低溫脅迫調(diào)控機制研究進展[J].中國農(nóng)學通報,2022,38(17):51-61.
CHEN S Q,SUN J SH,MA W J,et al.Research progress on the regulation mechanism of plant low temperature stress [J].Chinese Agronomy Bulletin,2022,38(17):51-61.
[20] DERANIEH R M,HE Q,CARUSO J A,et al.Phosphorylation regulates myo-inositol-3-phosphate synthase:a novel regulatory mechanism of inositol biosynthesis [J].Journal of Biological Chemistry,2013,288(37) 26822-26833.
[21] 王 超.響應大豆胞囊線蟲脅迫的GmMIPS基因的表達分析及亞細胞定位的研究[D].沈陽:沈陽農(nóng)業(yè)大學,2019.
WANG CH.Expression analysis and subcellular localization of GmMIPS gene in response to soybean cyst nematode stress [D].Shenyang:Shenyang Agricultural University,2019.
[22] ROHIT J,MANGU V R,NIRANJAN B. Arabidopsis plants constitutively overexpressing a myo-inositol 1-phosphate synthase gene (SaINO1) from the halophyte smooth cordgrass exhibits enhanced level of tolerance to salt stress[J].Plant Physiology and Biochemistry,2013,65(6):61-66.
[23] MARC R. K,HEATHER K. Low-temperature perception leading to gene expression and cold tolerance in higher plants[J].New Phytologist,2012,195(4):737-751.
[24] HARMEET K,POOJA V,BHANU P P,et al.Ectopic expression of the ABA-inducible dehydration-responsive chickpea l - myo -inositol 1-phosphate synthase 2 (CaMIPS2 ) in Arabidopsis enhances tolerance to salinity and dehydration stress[J].Planta,2013,237(1):321-335.
[25] 趙志宏. OsMIPS在水稻生長發(fā)育及逆境脅迫響應中的功能分析[D].廣州:華南農(nóng)業(yè)大學,2012.
ZHAO ZH H.Functional analysis of OsMIPS in rice growth and development and stress response [D]. Guangzhou:South China Agricultural University,2012.
[26] 顧海科.水稻和擬南芥中類三磷酸肌醇受體蛋白的免疫學證據(jù)[D].北京:中國農(nóng)業(yè)大學,2004.
GU H K.Immunological evidence of inositol triphosphate like receptor protein in rice and Arabidopsis [D]. Beijing:China Agricultural University,2004.
[27] 王明強,張道遠.植物熱激蛋白70基因家族及其生物學功能研究進展[J].基因組學與應用生物學,2015,34(2):421-428.
WANG M Q,ZHANG D Y.Research progress of plant heat shock protein 70 gene family and its biological function [J].Genomics and Applied Biology,2015,34(2):421-428.
[28] ROCCO M,ARENA S,RENZONE G,et al.Proteomic?analysis of temperature stress-responsive proteins in Arabidopsis thaliana rosette leaves[J].Molecular BioSystems,2013,9(6):1257-1267.
[29] POWLES S B,BERRY J A,BJRKMAN O. Interaction between light and chilling temperature on the inhibition of photosynthesis in chilling-sensitive plants[J].Plant Cell & Environment,2010,6(2):117-123.
[30] 孫旭紅,周煥斌,王道文,等.植物鈣聯(lián)蛋白與鈣網(wǎng)蛋白的研究進展[J].中國農(nóng)學通報,2018,34(9):48-52.
SUN X H,ZHOU H? B,WANG D W,et al.Research progress of plant cadherin and calreticulin [J].Chinese Agronomy Bulletin,2018,34(9):48-52.
[31] KOMATSU S,YAMADA E,F(xiàn)URUKAWA K.Cold stress changes the concanavalin A-positive glycosylation pattern of proteins expressed in the basal parts of rice leaf sheaths[J].Amino Acids,2009,36(1):115-123.
[32] SHARMA A,ISOGAI M.A novel interaction between calreticulin and ubiquitin-like nuclear protein in rice[J].Plant Cell,2004,45(6):684-692.
[33] CHEN Y T,LEE Y R,YANG C Y,et al. A novel papaya ACC oxidase gene (CP-ACO2) associated with late stage fruit ripening and leaf senescence[J].Plant Science,2003,164(4):531-540.
[34] 吳錦程,吳畢莎,黃審劍,等.枇杷幼果PLD和LOX對低溫脅迫的響應[J].植物科學學報,2015,33(2):203-209.
WU J CH,WU B SH,HUANG SH? J,et al. Response of young loquat fruit PLD and LOX to low temperature stress[J].Acta Plant Science,2015,33(2):203-209.
[35] 廖晶晶.薄皮甜瓜CmLOX10在植物成熟衰老和干旱響應中的作用[D].沈陽:沈陽農(nóng)業(yè)大學,2018.
LIAO? J? J.The role of muskmelon CmLOX10 in plant maturation,senescence and drought response[D]. Shenyang:Shenyang Agricultural University,2018.
Abstract? In order to explore the response mechanism of potato to low temperature stress at the proteomic level,the leaf proteomic of the plantlets of the low-temperature-tolerant variety ‘DR-2 and the low-temperature sensitive variety ‘Favorite were analyzed in this study,the results showed that there were? 282 790 peptides,19 099 unique peptides and 4 471 proteins,and? there were? 38 differentially expressed proteins in ‘DR-2 and 72 differentially expressed proteins in ‘Favorite under low temperature stress by Label-Free technique. And 13 of them were screened outin metabolic pathways,including 5 metabolism-related proteins,3 photosynthesis-related proteins,2 defense-related proteins and 3 transport-related proteins. Pathway enrichment analysis results showed that glyceraldehyde-3-phosphate dehydrogenase,calreticulin,heat shock protein,and inositol-3-phosphate synthase distributed in photosynthesis and energy metabolism pathways were differentially expressed in leaves under low temperature stress. This study providesa reference for study? on the low temperature response mechanism of potato.
Key words Potato;Low temperature stress; Proteomics; Response mechanism
