地方小麥品種貴協(xié)3號赤霉病抗性QTL的定位與效應(yīng)鑒定
2023-05-30 02:46:14鄭首航何員江鄒鳳亮杜小英歐俊梅雷加容閆海生賈海燕
麥類作物學(xué)報 2023年5期
鄭首航,何員江 ,陶 軍 ,張 華,吳 舸,鄒鳳亮 ,杜小英,歐俊梅,雷加容 ,閆海生,賈海燕,任 勇
(1.綿陽市農(nóng)業(yè)科學(xué)研究院,四川綿陽,621000; 2.南京農(nóng)業(yè)大學(xué)農(nóng)學(xué)院,江蘇南京,210095)
由鐮刀菌引起的赤霉病對小麥生產(chǎn)危害嚴(yán)重,可導(dǎo)致小麥產(chǎn)量和品質(zhì)下降[1]。近年來,由于全球氣候變暖、免耕技術(shù)的推廣以及秸稈還田等原因,國內(nèi)外小麥赤霉病的發(fā)生和流行程度日趨嚴(yán)重,引起國際廣泛關(guān)注[2]。該病在中國常年發(fā)生面積在267萬~333萬hm2,且有逐漸擴(kuò)大的趨勢[3],流行區(qū)域由傳統(tǒng)的長江中下游、黃淮麥區(qū)向華北麥區(qū)蔓延。由于赤霉病抗性受環(huán)境影響大以及抗病機(jī)制復(fù)雜,致使小麥抗赤霉病育種進(jìn)展緩慢,目前大面積推廣的品種赤霉病抗性普遍較差[4]。在已發(fā)現(xiàn)的赤霉病抗源中,僅有中國地方品種望水白和蘇麥3號表現(xiàn)較為突出[5]。挖掘抗病基因,解析分子機(jī)制以及培育抗赤霉病小麥品種,是小麥抗赤霉病育種工作的重中之重。
小麥在受到赤霉病病原菌侵染后,會產(chǎn)生一系列復(fù)雜的信號途徑激活應(yīng)答反應(yīng),誘導(dǎo)抗病相關(guān)基因的表達(dá),進(jìn)而引起蛋白以及代謝水平的變化,抵御病原菌的侵襲。小麥對赤霉病的抗性包括抗侵入(Type Ⅰ)、抗擴(kuò)展(Type Ⅱ)、抗毒素積累(Type Ⅲ)、降低病粒率(Type Ⅳ)和耐病性(Type Ⅴ)五種類型[6],是受多基因控制的數(shù)量性狀。隨著小麥基因組學(xué)的發(fā)展,借助DNA分子標(biāo)記和遺傳圖譜,已定位到400多個抗赤霉病基因/QTLs[7],分布在小麥21條染色體上,這些基因/QTLs主要來自蘇麥3號及其衍生系和望水白[7-8],其中Fhb1~Fhb7已被精細(xì)定位[9-16]。前人在CM82036[17]、DH181[18]、W14[19]、周麥27[20]等品種中均鑒定到Fhb5基因,是目前抗性最強(qiáng)、最穩(wěn)定的抗侵入相關(guān)基因[21]。
貴協(xié)3號是貴州省農(nóng)業(yè)科學(xué)院旱糧研究所2008年用以色列野生二粒小麥(Triticumdicoccoides)為母本,用光稃野燕麥 (AvenafatuaL.var.glabratapat)為父本進(jìn)行遠(yuǎn)緣雜交,獲得的F1代與貴農(nóng)22回交獲得的普通小麥材料。2013年,經(jīng)國家小麥產(chǎn)業(yè)技術(shù)體系在江蘇海安、南京和湖北武漢三地的赤霉病抗性鑒定中,貴協(xié)3號表現(xiàn)為中抗至抗病,2014年在海安試驗點也表現(xiàn)出較好的抗性,病情指數(shù)低于抗病對照品種蘇麥3號[22],但目前尚未有貴協(xié)3號抗赤霉病QTL的定位報道。本研究以感赤霉病品種綿麥96-5和貴協(xié)3號構(gòu)建的含有196個株系的DH群體為材料,結(jié)合兩年兩個地區(qū)的赤霉病嚴(yán)重度數(shù)據(jù),挖掘與赤霉病抗性相關(guān)的QTL,并分析其聚合效應(yīng),以期了解貴協(xié)3號抗赤霉病的遺傳基礎(chǔ),為小麥抗赤霉病育種提供幫助。
1 材料與方法
1.1 供試材料及田間種植
試驗材料為貴協(xié)3號和綿麥96-5及其雜交F1代通過花藥培養(yǎng)、染色體加倍獲得的含有196個株系的DH群體。貴協(xié)3號由貴州省農(nóng)業(yè)科學(xué)院旱糧研究所提供,綿陽96-5為四川省綿陽市農(nóng)業(yè)科學(xué)研究所自育品系。
于2018和2019年將DH群體及其親本分別種植在南京農(nóng)業(yè)大學(xué)試驗基地和四川省綿陽市農(nóng)業(yè)科學(xué)研究院試驗田(分別用2018NJ、2019MY表示),進(jìn)行赤霉病抗侵入表型鑒定,以貴協(xié)3號和綿麥96-5分別作為抗、感病對照。四種強(qiáng)致病菌種F15、F301、F7136以及GFP標(biāo)記過的菌株WT-GFP由南京農(nóng)業(yè)大學(xué)應(yīng)用基因組實驗室提供。
采用隨機(jī)區(qū)組試驗設(shè)計,每三行設(shè)置一空行,每個株系2個重復(fù),行寬1.5 m,行距0.3 m,每行播種25粒。
1.2 田間接種及抗性鑒定
將小麥籽粒在清水中浸泡過夜,在沸水中煮30 min,晾干表面水分后分裝至500 mL組培瓶中,121 ℃滅菌30 min后,于烘箱中烘干3~5 min,即為麥粒培養(yǎng)基。將四種強(qiáng)致病菌種F15、F301、F7136以及GFP標(biāo)記過的菌株WT-GFP接種在麥粒培養(yǎng)基中,室溫培養(yǎng)10 d左右,待培養(yǎng)基表面布滿白色菌絲后,于開花前4 d在田間進(jìn)行撒播,撒播小麥病麥粒后,盡量保持濕潤,于開花后10 d進(jìn)行抗性表型鑒定,調(diào)查病小穗數(shù)和總小穗數(shù),每個株系調(diào)查20個穗,嚴(yán)重度計算采用下面公式:嚴(yán)重度(%)=病小穗數(shù)/總小穗×100%[21],取平均值記為株系的嚴(yán)重度。3次重復(fù)。鑒定圃周圍試驗田未進(jìn)行其他菌株和其他病原菌接種。
1.3 統(tǒng)計分析
根據(jù)赤霉病嚴(yán)重度,將DH群體的196個株系分為三種不同的表型:抗病親本類型(嚴(yán)重度在10%以下)、感病親本類型(嚴(yán)重度在10%~50%之間)和不同于親本的其他類型(嚴(yán)重度在50%以上)[23]。
1.4 基因型分型、連鎖圖譜的構(gòu)建和QTL分析
采用SPSS Statistics 17.0對表型數(shù)據(jù)進(jìn)行t檢驗、相關(guān)性分析等。用CTAB法[24]提取DH群體及親本的DNA,用澳大利亞基因芯片公司的55K DArT標(biāo)記基因芯片雜交技術(shù)平臺對196個DH株系及親本進(jìn)行全基因組掃描,利用QTL IciMapping V4.1軟件的BIN功能對冗余標(biāo)記進(jìn)行刪除,篩選出兩親本間存在多態(tài)性的標(biāo)記,并進(jìn)行連鎖圖譜的構(gòu)建,利用完備區(qū)間作圖法(ICIM)對QTL進(jìn)行定位分析;通過軟件的1000 permutation功能計算LOD閾值,P值設(shè)定為0.05,檢測QTL的顯著性。與中國春最新參考基因組(IWGSC RefSeq v1.0)進(jìn)行比對,獲得標(biāo)記的物理位置。
2 結(jié)果與分析
2.1 DH群體的赤霉病嚴(yán)重度鑒定結(jié)果
感病親本綿麥96-5在2018年江蘇南京和2019年四川綿陽試驗基地的赤霉病平均嚴(yán)重度為36%和53%,而抗病親本貴協(xié)3號的平均嚴(yán)重度為13%和20%,DH群體的平均嚴(yán)重度為5%~93%,該群體后代株系的嚴(yán)重度均呈現(xiàn)連續(xù)性變異(圖1),分布頻率基本符合正態(tài)分布,表明該群體的赤霉病嚴(yán)重度屬于數(shù)量性狀,由多個基因控制。根據(jù)孟德爾遺傳分析法和數(shù)量性狀分析法,估算該群體至少含有2~4個抗侵入基因/QTLs(表1和表2)。
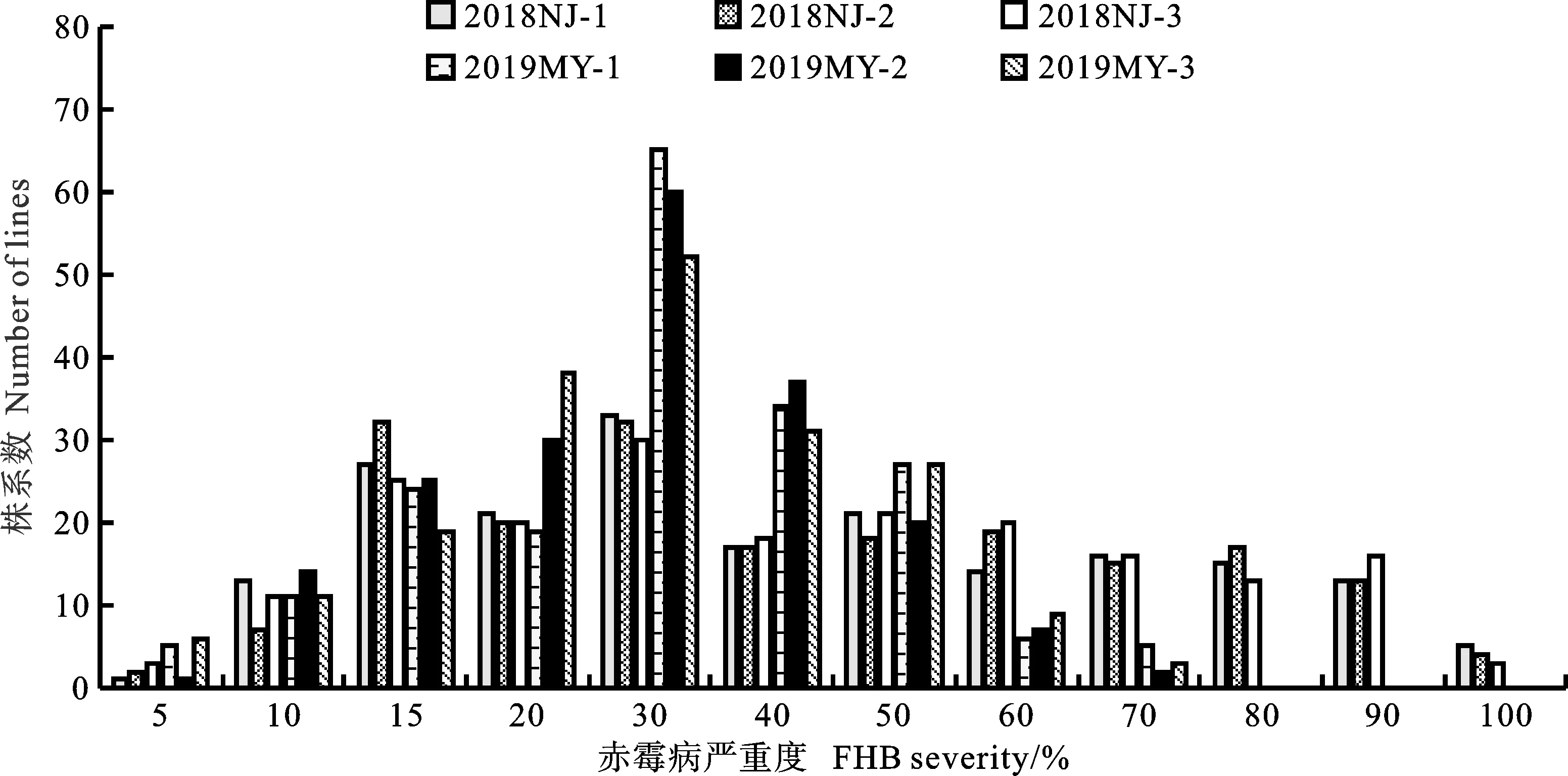
2018NJ:2018年南京;2019MY:2019年綿陽。表1~3同。1、2和3為3個重復(fù)。表1同。2018NJ:Naijing of 2018; 2019MY:Mianyang of 2019. The same in tables 1~3. 1,2 and 3 were three replicates. The same in table 1.圖1 DH群體赤霉病嚴(yán)重度的頻率分布Fig.1 Frequency distribution of FHB severity in DH population
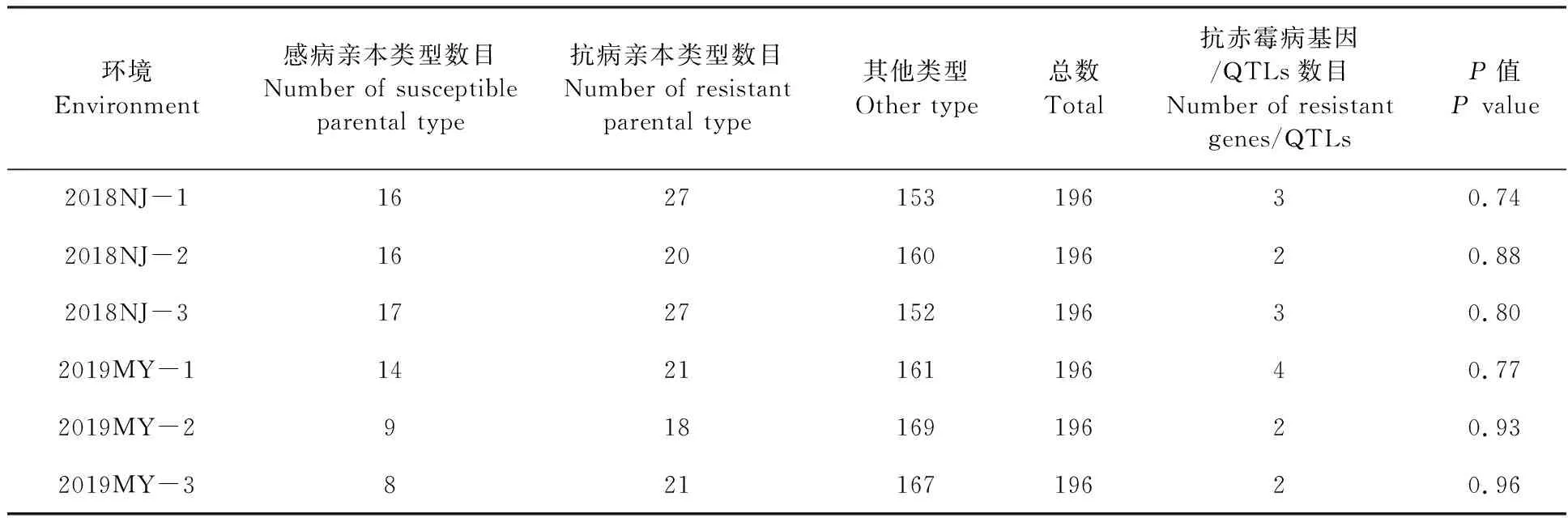
表1 孟德爾遺傳分析法估算DH群體中含有的抗赤霉病基因數(shù)目Table 1 Number of FHB resistant genes in DH population evaluated by Mendelian genetic analysis method

表2 數(shù)量性狀分析法估算DH群體中含有的抗赤霉病基因數(shù)目Table 2 Number of FHB resistance genes in DH population by quantitative genetic analysis method
2.2 遺傳連鎖圖譜的構(gòu)建
利用覆蓋小麥21條染色體的大約373 60個標(biāo)記的DArT-Seq芯片對親本及196個株系進(jìn)行基因分型,結(jié)果發(fā)現(xiàn)有13 472個DArT-Seq標(biāo)記位點(占標(biāo)記總數(shù)的36%)在親本間具有多態(tài)性。利用QTL IciMapping V4.1軟件的BIN功能,對冗余標(biāo)記進(jìn)行刪除,最終獲得1 432個DArT-Seq標(biāo)記。這些標(biāo)記構(gòu)成的遺傳連鎖圖譜覆蓋小麥全基因組,圖譜全長為15 195.8 cM,平均圖距為10.6 cM。
2.3 抗赤霉病QTL的定位結(jié)果
共檢測到3個抗赤霉病QTLs,其中QTL-FHB.GX-2B在2B染色體上,與AX-111081804標(biāo)記緊密連鎖;QTL-FHB.GX-5B在5B染色體上,與AX-111012188標(biāo)記緊密連鎖;QTL-FHB.GX-7A在7A染色體上,位于標(biāo)記AX-110915586 ~AX-108934756之間,區(qū)間長度為16 cM (圖2)。這3個QTLs的抗性等位基因均來自于抗病親本貴協(xié)3號,LOD值分別為12.7、8.5和10.6,分別可解釋1.5%、1.2%和1.3%的表型變異;這3個抗赤雷病QTLs在2018南京和2019綿陽兩個環(huán)境下均能檢測到,抗性表現(xiàn)較為穩(wěn)定。
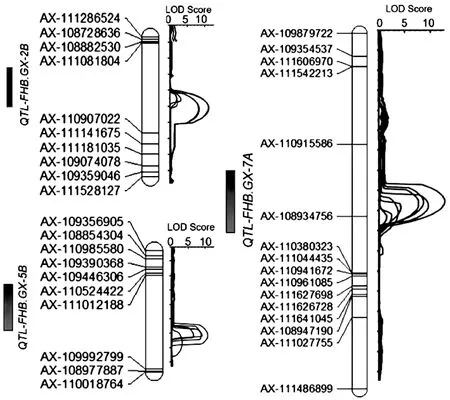
圖2 抗赤霉病QTL在染色體上的分布Fig. 2 Distribution of QTLs for resistance to FHB on chromosomes of wheat
2.4 QTL的加性互作效應(yīng)
利用與3個QTL緊密連鎖的分子標(biāo)記的基因型數(shù)據(jù),分析其累加效應(yīng),結(jié)果在196個株系中共發(fā)現(xiàn)8種抗赤霉病單倍型(表3)。其中,32個株系含有3個抗赤霉病QTL,2018南京和2019綿陽兩個環(huán)境的平均嚴(yán)重度分別為24%和30% (表3);72個株系含有2個抗赤霉病QTL,2018南京和2019綿陽兩個環(huán)境的平均嚴(yán)重度分別為26%~31% 和36%~41%;68個株系只含有1個QTL,2018南京和2019綿陽兩個環(huán)境的平均嚴(yán)重度分別為27%~32%和39%~46%;24個株系不含有檢測到的抗赤霉病QTL,2018南京和2019綿陽兩個環(huán)境的年平均嚴(yán)重度分別為33%和50%。這說明含有多個抗赤霉病QTL株系的赤霉病嚴(yán)重度明顯低于含有單個和不含抗赤霉病QTL的株系。
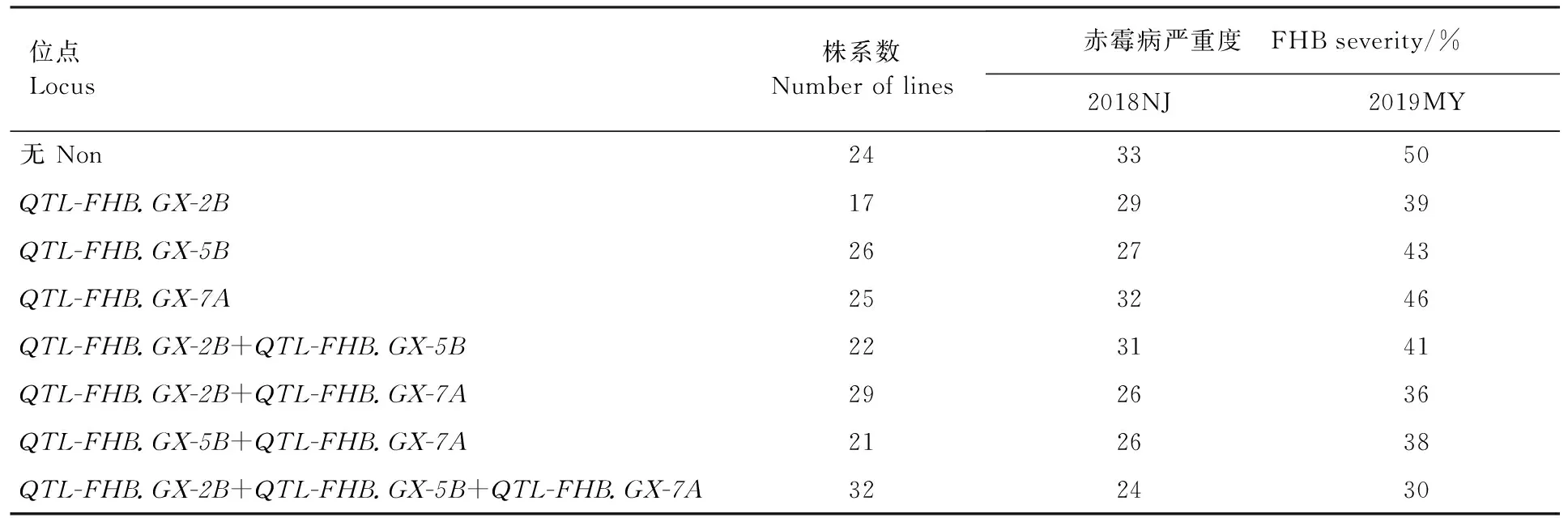
表3 QTL組合對赤霉病嚴(yán)重度的影響Table 3 Effect of QTL combinations to FHB severity
3 討論
本研究利用數(shù)量性狀分析法以及孟德爾遺傳分析法,預(yù)測到 DH群體(貴協(xié)3號×綿麥96-5)的抗赤霉病性狀受3~4個加性基因控制,進(jìn)一步利用QTL IciMappingV4.1軟件共定位到3個抗赤霉病QTL,與數(shù)量性狀分析法估算群體中的基因數(shù)目基本一致。本研究在7A染色體上定位到的QTL-FHB.GX-7A(物理區(qū)間為15 681 840~91 358 341 bp),與Zhang等[25]在7A染色體上定位到的QTL(區(qū)間為21 763 191~33 614 065 bp)區(qū)間存在部分重疊,但尚不能確定是否為同一個基因。本研究定位到的3個QTL的抗性等位基因均來自于抗病親本貴協(xié)3號,表型變異解釋率為1.2%~1.5%,推測貴協(xié)3號的赤霉病抗性是多個微效基因的累加效應(yīng),后續(xù)將會采取相關(guān)試驗進(jìn)行進(jìn)一步的驗證。本研究同時也進(jìn)一步證明,以色列野生二粒小麥與光稃野燕麥遠(yuǎn)緣雜交獲得的貴協(xié)3號,對小麥赤霉病具有良好的抗性。在目前我國赤霉病危害不斷蔓延的情況下,貴協(xié)3號可作為抗病資源在小麥抗病育種中加以利用。
在小麥抗赤霉病研究中,經(jīng)國內(nèi)外科學(xué)家的不懈努力,已經(jīng)取得了長足的進(jìn)展。如抗赤霉病基因Fhb1在美國、日本以及我國江蘇等地區(qū)均已得到成功應(yīng)用;抗性資源蘇麥3號和望水白抗赤霉病位點的有利等位變異,也已進(jìn)行了分子標(biāo)記輔助選擇聚合育種[26]。近幾年我國不斷加強(qiáng)對赤霉病的防控工作,已經(jīng)培育出一批達(dá)到抗或中抗赤霉病的小麥新品種。但由于全球氣溫變暖和種植制度的不斷改變,小麥赤霉病在我國已經(jīng)從長江中下游、黃淮麥區(qū)擴(kuò)展到全國各大麥區(qū),加強(qiáng)小麥抗赤霉病的研究以及抗赤霉病新品種的選育任重而道遠(yuǎn)。
