抗氰烯菌酯假禾谷鐮孢突變體的誘導及其生物學特性
2023-05-30 14:46:50逯燕騰李新蕾鄧淵鈺曹淑琳李偉孫海燕陳懷谷
植物保護 2023年2期
逯燕騰 李新蕾 鄧淵鈺 曹淑琳 李偉 孫海燕 陳懷谷
摘要 為評估引起小麥莖基腐病的病原菌假禾谷鐮孢Fusarium pseudograminearum對氰烯菌酯的抗性風險,對5株敏感菌株進行了室內藥劑馴化,獲得33株抗性突變體,突變頻率為16.5%,其對氰烯菌酯的抗性水平范圍為7.39~1 665.76倍,3株表現低抗,4株表現中抗,26株表現高抗;發現在myosin-5基因上存在11種抗性突變類型,其中217位的絲氨酸突變為亮氨酸(S217L)、420位的谷氨酸突變為賴氨酸(E420K)和135位的丙氨酸突變為蘇氨酸(A135T)為主要突變類型,其比例分別為45.5%、15.2%和9.1%。S217L型抗性突變體的產孢量顯著下降,菌絲生長速率和致病力與親本菌株無顯著差異。E420K型抗性突變體的菌絲生長速率和致病力顯著下降,產孢量與親本菌株無顯著差異。A135T型抗性突變體的菌絲生長速率和產孢量與親本菌株無顯著差異。研究結果表明假禾谷鐮孢在藥劑選擇壓力下易形成氰烯菌酯的抗性群體,對氰烯菌酯存在中到高等的潛在抗性風險,其myosin-5的點突變與其對氰烯菌酯的抗性相關。
關鍵詞 小麥莖基腐病;?假禾谷鐮孢;?氰烯菌酯;?抗藥性;?生物學特性;?myosin-5
中圖分類號: S 435.121.49
文獻標識碼:?A
DOI:?10.16688/j.zwbh.2021707
Abstract In order to assess the resistance risk of Fusarium pseudograminearum to phenamacril, a total of 33 phenamacril-resistant mutants were obtained through spontaneous selection with a mutation frequency of 16.5%, and the resistance level of these mutants to phenamacril ranged from 7.39 to 1 665.76 times, with three mutants showing low resistance, four mutants showing moderate resistance and 26 mutants showing high resistance. Eleven types of resistance mutations were found in myosin-5 gene. A serine (S) to leucine (L) transition at position 217 (S217L), a glutamic (E) to lysine (K) transition at position 420 (E420K), and an alanine (A) to threonine (T) transition at position 135 (A135T) were major mutation types, with a frequency of 45.5%, 15.2% and 9.1%, respectively. Compared with the parental strains, the spore production of the type S217L resistant mutants was significantly decreased, but there was no significant difference in mycelial growth rate and pathogenicity. Mycelial growth rate and pathogenicity of the type E420K resistant mutants were significantly decreased, and no significant difference was observed in spore production. The point mutation of myosin-5 was related to its resistance to phenamacril. The mycelial growth rate and spore production of A135T resistant mutants were not significantly different from the parental strains. The results suggested that F.pseudograminearum could easily form the phenamacril-resistant population under the pressure of fungicide selection, and had a moderate or high risk of resistance to phenamacril.
Key words wheat crown rot;?Fusarium pseudograminearum;?phenamacril;?resistance;?biological characteristics;?myosin-5
小麥莖基腐病(crown rot, CR)是由多種鐮刀菌引起的土傳病害[1-3],在美國、加拿大、澳大利亞、中國等地廣泛發生[4-8],我國小麥莖基腐病的主要病原菌為假禾谷鐮孢Fusarium pseudograminearum、亞洲鐮孢F.asiaticum和禾谷鐮孢F.graminearum等。不同地區引起莖基腐病的優勢種不同,河南、河北、山西及山東等地以假禾谷鐮孢為優勢種,而安徽、江蘇等地以禾谷鐮孢復合種為主[1,7-8]。
小麥莖基腐病從小麥分蘗到成熟期均可發生,感病小麥莖基部葉鞘和莖稈變褐,莖節壞死,嚴重時植株死亡、枯白(孕穗期),出現白穗,小麥產量下降[8]。2010年前,該病害在我國偶有發生,危害不嚴重。近年來由于秸稈還田使土壤內病原菌不斷積累、不合理使用化肥導致土壤鹽漬化嚴重,該病害在我國黃淮麥區逐年加重,對小麥安全生產構成嚴重威脅[9-11]。據報道,2013年-2016年間小麥莖基腐病在河南省部分重病田造成小麥產量損失高達38.0%~61.3%[12-13];在河北省嚴重地塊白穗率高達20%~50%[14];2019年山東省鄒城市小麥莖基腐病地塊發生率高達66.8%[15];2017年-2019年,陜西省小麥莖基腐病發生面積均達到10萬 hm2以上[16]。
化學防治是目前控制該病害發生的主要方法之一。紀莉景等研究發現6種種衣劑包衣處理對小麥苗期莖基腐病病株率和病情指數的防效分別在60.4%~100.0%和45.5%~100.0%,隨著小麥生育期的推移,種衣劑的防治效果均呈下降趨勢[17]。張守成等研究發現種子處理或返青期噴施不同殺菌劑對小麥莖基腐病均有較好防效,但不同殺菌劑的防控效果及持效期有明顯差異[18]。
氰烯菌酯是一種2-氰基丙烯酸酯類殺菌劑,由江蘇省農藥研究所于1998年自主研發。它對鐮刀菌具有專化抗菌活性,已登記用于小麥赤霉病和水稻惡苗病的防治[19-20]。據報道,氰烯菌酯能抑制禾谷鐮孢肌球蛋白myosin-5馬達結構域的ATP酶活性,從而導致肌動蛋白絲聚合受阻,影響囊泡運輸,使菌絲細胞不能極性生長[21-23]。肌球蛋白myosin-5基因的突變導致了禾谷鐮刀菌對氰烯菌酯的抗性。目前該藥劑還未在小麥莖基腐病防治上登記使用,也未有假禾谷鐮孢對其抗性的研究報道。本研究以假禾谷鐮孢為研究對象,對其進行氰烯菌酯抗性突變體的誘導,并對抗性機理和突變體生物學特性進行研究,以期為氰烯菌酯在小麥莖基腐病防治上的應用提供參考。
1?材料與方法
1.1?供試材料
供試菌株: 5株分別從山東壽光、江蘇鹽城和連云港采集的假禾谷鐮孢菌株CF1558、CF1566、CF15128、CF15263和CF15281,由江蘇省農業科學院植物保護研究所小麥病害防控團隊分離純化、保存和鑒定。
供試藥劑:95%氰烯菌酯原藥,由江蘇省農藥研究所股份有限公司提供,用甲醇溶解配制成1×104μg/mL的母液,于4℃保存,備用。
馬鈴薯葡萄糖瓊脂(PDA)培養基:馬鈴薯200 g、葡萄糖20 g、瓊脂粉15 g,加蒸餾水定容至1 L,用于菌株的培養及保存;羧甲基纖維素鈉(CMC)培養基:羧甲基纖維素鈉15 g、NH4NO3 1 g、KH2PO4 1 g、MgSO4·7H2O 0.5 g、酵母提取物1 g,加蒸餾水定容至1 L,用于菌株產孢培養。
1.2?方法
1.2.1?假禾谷鐮孢抗性突變體誘導
將5株假禾谷鐮孢分別接種到PDA平板上預培養4 d后,于菌落邊緣打取直徑5 mm的菌餅,將菌餅菌絲面朝下接于含5 μg/mL氰烯菌酯的PDA平板上[24],每平板接種10個菌餅,每菌株接種40個,25℃黑暗培養。培養過程中出現的扇形突變菌落即為疑似抗性突變體,將其轉接到含5 μg/mL氰烯菌酯的PDA平板上進行抗性驗證,能夠生長的即為抗性突變體,命名為R-親本菌株編號后兩位-序號。計算抗性突變頻率。
抗性突變頻率=(抗性突變體數/藥劑馴化的菌餅數)× 100%。
1.2.2?抗性水平測定
將抗性突變體及其親本菌株分別接種到PDA平板上,25℃預培養4 d后,在菌落邊緣打取直徑為5 mm的菌餅,親本菌株轉接到含0、0.125、0.25、0.5、1、2、4 μg/mL氰烯菌酯的PDA平板上,抗性突變體接于含0、0.625、1.25、2.5、5、10、20、40、80、160 μg/mL氰烯菌酯的PDA平板上,25℃黑暗培養4 d,十字交叉法測量菌落直徑取平均值。每菌株3次重復。根據各突變體的EC50與其親本菌株的EC50比值計算抗性倍數,根據抗性倍數將各突變體的抗性水平劃分為敏感、低抗、中抗和高抗[25],其中:抗性倍數≤3的為敏感菌株(S);3<抗性倍數≤10的為低抗菌株(LR);10<抗性倍數≤100的為中抗菌株(MR);抗性倍數>100的為高抗菌株(HR)。
1.2.3?抗性遺傳穩定性測定
從藥劑馴化獲得的抗性突變體中每種突變類型各選取1株進行測定。將-80℃保存6個月后的11株抗性突變體在無藥PDA平板上繼代培養4代,25℃黑暗培養5 d,按1.2.2節方法中的十字交叉法測定抗性突變體對氰烯菌酯的敏感性(EC50),計算抗性倍數,并根據抗性突變體的生長速度確定其遺傳穩定性。
1.2.4?myosin-5 基因擴增及序列分析
將抗性突變體及其親本菌株在PDA平板上25℃恒溫培養4 d后,收集菌絲,采用CTAB法提取菌絲基因組DNA[26]。使用myosin-5基因序列特異性引物5-F(5′-CGACAAATTCACTTCAGGCTC-3′),5-R(5′-TTGCCTGTCTGCTACCATCA-3′),以提取的基因組DNA為模板, 使用南京諾唯贊生物科技有限公司的Max Super-Fidelity DNA Polymerase進行PCR擴增,擴增程序為:95℃預變性3 min;95℃變性15 s,58℃退火15 s,72℃延伸60 s,循環35次;72℃延伸5 min,4℃保存。擴增產物采用1%瓊脂糖凝膠電泳檢測,將目的擴增產物送至生工生物工程 (上海) 股份有限公司測序,分析突變體myosin-5基因的堿基及氨基酸變化情況。
1.2.5?抗性突變體的生物學特性研究
1.2.5.1?菌絲生長速率測定
將抗性突變體及其親本菌株分別沿菌落邊緣,用直徑為5 mm的打孔器打取菌餅,接種于無藥PDA平板上,25℃黑暗培養4 d后,十字交叉法測量菌落直徑[27]。每菌株3次重復。
1.2.5.2?產孢量測定
將抗性突變體及其親本菌株沿菌落邊緣打取直徑為5 mm的菌餅,放入裝有50 mL CMC的培養瓶中,每瓶接種5個菌餅,25℃、150 r/min振蕩培養,誘導產孢。5 d后無菌條件下過濾獲得孢子懸浮液,適量無菌水稀釋至合適濃度,用血球計數板在顯微鏡下計算孢子數量。每菌株3次重復。
1.2.5.3?致病力測定
抗性突變體及其親本菌株對苗期小麥的致病力測定參照Gardiner等[28]的方法。采用1.2.5.2方法誘導產孢,將孢子懸浮液濃度調整至1×105個/mL。將消毒后的‘揚麥158小麥種子置于鋪有單層濾紙的培養皿中,清水保濕,在25℃恒溫培養箱中進行催芽。2 d后選取萌發一致的種子浸蘸孢子懸浮液10 s,用無紡布包裹保持均勻間距的10粒種子豎直放置于裝有適量水的廣口塑料瓶中,25℃、L∥D=12 h∥12 h光暗交替的培養箱培養,培養過程中清水保濕。每處理3次重復,14 d后記錄幼苗發病情況。參照Bovill等[29]的方法根據第1葉鞘下莖稈褐變長度,對小麥莖基腐病發生的嚴重度進行分級,0級:無病;1級:褐變長度為25%以下;2級:褐變長度為25%~50%;3級:褐變長度為51%~75%;4級:褐變長度為76%~100%。計算各處理的病情指數。
病情指數=100×Σ(各級病株數×病級值)/(調查總株數×4)。
1.3?數據分析
采用DPS數據處理系統LSD法對試驗數據進行統計分析,測序結果用DNAMAN軟件進行序列對比分析。
2?結果與分析
2.1?假禾谷鐮孢抗性突變體的類型
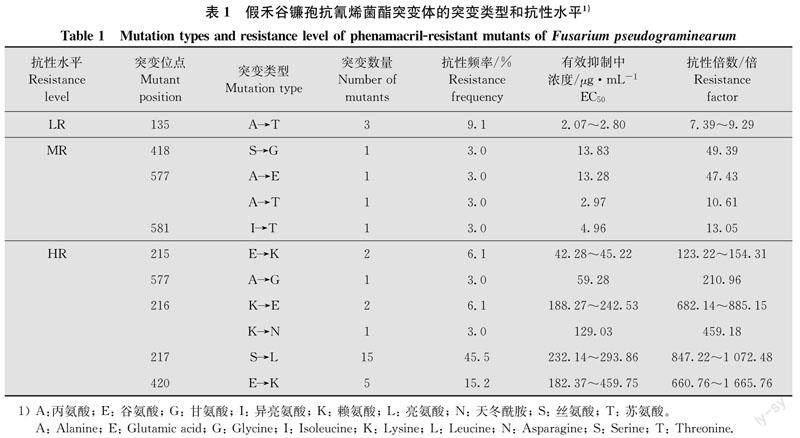
5株親本菌株經藥劑馴化25 d后,共獲得33株抗性突變體,平均突變頻率為16.5%。通過對抗性突變體及其親本菌株的myosin-5基因序列分析,發現11種突變類型,其中217位絲氨酸(serine)突變為亮氨酸(leucine) (S217L)、420位谷氨酸(glutamic acid)突變為賴氨酸(lysine) (E420K)和135位丙氨酸(alanine)突變為蘇氨酸(threonine) (A135T)為主要突變類型,其比例分別為45.5%、15.2%和9.1%(表1)。
2.2?假禾谷鐮孢抗性突變體的抗性水平
5株親本菌株對氰烯菌酯的EC50為0.27~0.38 μg/mL,33株抗性突變體的EC50為2.07~459.75 μg/mL,抗性水平范圍為7.39~1 665.76倍。33株抗性突變體中3株表現低抗,4株表現中抗,26株表現高抗。3種主要突變類型中,S217L型和E420K型表現高抗,A135T型表現低抗(表1)。
2.3?假禾谷鐮孢抗性突變體的遺傳穩定性
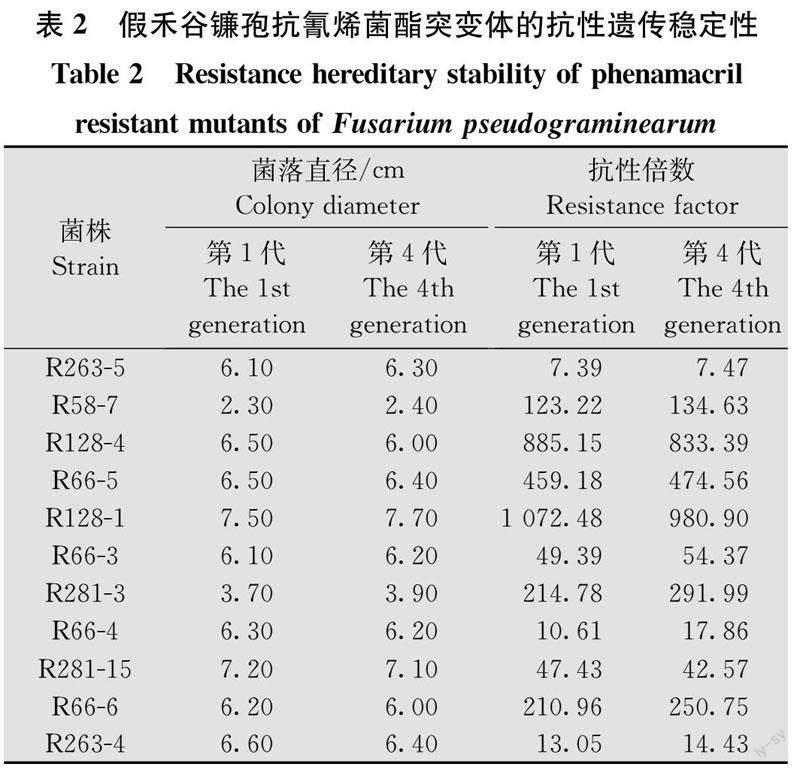
經藥劑馴化的11株抗性突變體在無藥的PDA培養基上繼代培養4代后,第1代和第4代的菌落生長速度無明顯差異,對氰烯菌酯的EC50也無明顯變化(表2),表明室內藥劑馴化獲得的假禾谷鐮孢抗氰烯菌酯突變體可穩定遺傳。
2.4?假禾谷鐮孢抗性突變體的生物學特性
2.4.1?菌絲生長速率
抗性突變體及其親本菌株培養4 d后,親本菌株菌落直徑在6.07~7.40 cm之間,抗性突變體菌落直徑在2.30~7.57 cm之間(表3)。差異顯著性分析結果表明,E420K、E215K和K216E型抗性突變體的菌絲生長速率顯著低于其親本菌株(表3,圖1),其他抗性突變體與其親本菌株沒有顯著性差異 (P>0.05)。
2.4.2?產孢能力
S217L、E215K和I581T型抗性突變體的產孢量顯著低于其親本菌株(表3),其他類型突變體的產孢量與其親本菌株相似,不存在顯著差異(P>0.05)。
2.4.3?致病力
與親本菌株相比,S217L、S418G和A577T型抗性突變體的致病力無顯著差異,其他突變體的致病力顯著下降(P<0.05) (圖2),病情指數下降范圍在5.37~48.44(表3)。
3?結論與討論
小麥莖基腐病是近年來我國小麥生產中發展迅速、危害嚴重的病害之一。該病害在小麥整個生育期都可發生,目前還未有完善的防治體系。周淼平等研究發現黃淮麥區大部分小麥推廣品種對小麥莖基腐病表現感病或高度感病,無高抗品種,只有極少數表現中抗[30]。因此在缺乏好的抗病品種情況下,化學防治是確保小麥穩產的重要保障。目前小麥莖基腐病化學防治研究主要集中于篩選有效藥劑和明確藥劑防效[15-18],對藥劑的抗性風險評估和抗性機制研究還很少。前期研究結果表明,氰烯菌酯對假禾谷鐮孢等小麥莖基腐病菌具有較好的抑制作用,對于田間病害的發生也具有較好的控制效果。明確小麥莖基腐病菌對氰烯菌酯的抗性風險和抗性機制對于該藥劑在防治小麥莖基腐病中的開發應用有重要意義。
已有研究表明,氰烯菌酯的作用靶標是病原菌的肌球蛋白-5(myosin-5)[23,31]。肌球蛋白是真核細胞中肌動蛋白依賴的ATP酶的馬達蛋白,在細胞質運動、細胞極性、轉錄因子調控、胞內運輸和信號轉導上有著極其重要的作用[23,31]。病原菌對氰烯菌酯產生抗性與肌球蛋白-5點突變相關。周明國教授團隊研究發現小麥赤霉病菌的肌球蛋白-5至少有12個氨基酸殘基可發生總頻率為23.9%的不同抗藥性水平變異[32],其中最常見的突變類型是217位的絲氨酸突變為亮氨酸(S217L)和420位的谷氨酸突變為賴氨酸(E420K)[33]。李斌研究發現水稻惡苗病菌對氰烯菌酯產生田間抗性的主要原因是肌球蛋白-5的219位由絲氨酸突變為脯氨酸(S219P)和絲氨酸突變為亮氨酸(S219L)[34]。本研究發現假禾谷鐮孢對氰烯菌酯的抗性至少存在11種突變類型,主要突變類型與小麥赤霉病菌相似,都為S217L和E420K型,這表明不同鐮刀菌對氰烯菌酯的抗性機制相似。
氰烯菌酯作用位點單一,其抗性風險備受關注。本文研究發現在離體條件下假禾谷鐮孢對氰烯菌酯易產生抗性,平均突變頻率為16.5%,這與亞洲鐮孢對氰烯菌酯的突變頻率相近,這可能與氰烯菌酯的作用靶標相關,且其作用靶標肌球蛋白-5的馬達結構域在藥劑作用下容易發生改變,從而導致抗性的產生。在獲得的抗性突變體中主要突變類型(S217L和E420K)的抗性水平高,且S217L型抗性突變體的菌絲生長速率、致病力與其親本相似,表明假禾谷鐮孢對氰烯菌酯存在潛在的抗性風險。S217L型抗性突變體的產孢能力顯著低于其親本菌株,E420K型抗性突變體的菌絲生長速率、致病力顯著低于其親本菌株,表明這兩種主要抗性突變體的適合度弱于親本菌株,將來在田間流行的風險較低。周明國教授團隊發現小麥赤霉病菌對氰烯菌酯的主要抗性突變類型(S217L)也是產孢能力低于親本菌株[33],推測與肌球蛋白-5結構域內類似“口袋”的結構有關,在誘導過程中這類區域的變化可以直接影響抗藥水平,也有可能間接影響病菌的其他表型。大部分在室內誘導下表現出高抗的抗性菌株的產孢量發生了顯著的降低,減弱了突變菌株在田間的適應性,從而降低了抗性菌株在田間的種群比例[35],這可能是氰烯菌酯在小麥赤霉病防治上使用近十年還一直未有抗藥性報道的原因之一。
氰烯菌酯于2012年同時登記用于小麥赤霉病和水稻惡苗病的防治,但目前僅有水稻惡苗病菌對其有田間抗性的報道[31]。這兩種病原菌對氰烯菌酯田間抗性風險的差異可能與病原菌本身特性相關,也有可能與施藥方式、施藥劑量相關,氰烯菌酯在小麥赤霉病防治上主要是揚花期噴霧,在水稻惡苗病防治上主要是浸種。小麥莖基腐病的化學防治既可以用藥劑種子包衣,也可以在小麥拔節前噴施藥劑,不同用藥方式是否會導致病原菌對藥劑的抗性風險存在差異需要進行進一步的研究。
本研究尚處于室內試驗階段,所得結果僅可為假禾谷鐮孢對氰烯菌酯的抗性風險評估提供參考,后續還將深入開展研究。
參考文獻
[1]?DENG Yuanyu, LI Wei, ZHANG Peng, et al. Fusarium pseudograminearum as an emerging pathogen of crown rot of wheat in eastern China [J]. Plant Pathology, 2020, 69(2): 240-248.
[2]?DYER A T, JOHNSTON R H, HOGG A C, et al. Comparison of pathogenicity of the Fusarium crown rot (FCR) complex (F.culmorum, F. pseudograminearum and F. graminearum) on hard red spring and durum wheat [J]. European Journal of Plant Pathology, 2009, 125(3): 387-395.
[3]?CHAKRABORTY S, OBANOR F, WESTECOT T R, et al. Wheat crown rot pathogens Fusarium graminearum and F.pseudograminearum lack specialization [J]. Phytopathology, 2010, 100(10): 1057-1065.
[4]?MITTER V, FRANCL L J, ALI S, et al. Ascosporic and conidial inoculum of Gibberella zeae play different roles in Fusarium head blight and crown rot of wheat in Australia and the USA [J]. Australasian Plant Pathology, 2006, 35(4): 441-452.
[5]?BENTLEY A R, CROMEY M G, FARROKHI-NEJAD R, et al. Fusarium crown and root rot pathogens associated with wheat and grass stem bases on the South Island of New Zealand [J]. Australasian Plant Pathology, 2006, 35(5): 495-502.
[6]?SUKUMAR C, FRIDAY O, RHYANNYN W, et al. Wheat crown rot pathogens Fusarium graminearum and F.pseudograminearum lack specialization [J]. Phytopathology, 2010, 100(10): 1057-1065.
[7]?張向向,孫海燕,李偉,等.我國冬小麥主產省小麥莖基腐鐮孢菌的組成及其致病力[J].麥類作物學報,2014,34(2):272-278.
[8]?LI Honglian, YUAN Hongxia, FU Bo, et al. First report of Fusarium pseudograminearum causing crown rot of wheat in Henan, China [J]. Plant Disease, 2012, 96(7): 1065.
[9]?王莉莉,朱凱,孫瑩,等.醚菌酯對假禾谷鐮孢的抑制作用及對小麥莖基腐病的防效研究[J]. 植物病理學報: 2022, 52(3):?434-442.
[10]胡蓉,鄭露,劉浩,等.秸稈還田對水稻根際微生物多樣性和水稻紋枯病發生的影響[J].植物保護學報,2020,47(6):1261-1269.
[11]楊云,賀小倫,胡艷峰,等.黃淮麥區主推小麥品種對假禾谷鐮刀菌所致莖基腐病的抗性[J].麥類作物學報,2015,35(3):339-345.
[12]賀小倫,周海峰,袁虹霞,等.河南和河北冬小麥區假禾谷鐮孢的遺傳多樣性[J].中國農業科學,2016,49(2): 272-281.
[13]徐飛,宋玉立,周益林,等.2013-2016年河南省小麥莖基腐病的發生危害情況及特點[J].植物保護,2016,42(6):126-132.
[14]紀莉景,栗秋生,王連生,等.河北省小麥冠腐病發生現狀及病原菌初探[J].植物保護,2016,42(5):154-157.
[15]趙艷麗,王祥會,胡英華,等.種子包衣技術防治小麥莖基腐病的田間藥效試驗[J].中國農技推廣,2020,36(5): 63-65.
[16]魏會新,郭海鵬,賀亞紅,等.陜西省小麥莖基腐病防控技術研究[J].陜西農業科學,2021,67(1): 1-3.
[17]紀莉景,王亞嬌,栗秋生,等.防治小麥莖基腐病種衣劑的篩選及全生育期防控效果[J].河北農業科學,2020,24(2):42-47.
[18]張守成,吳國峰,徐加健,等.不同殺菌劑處理對小麥莖基腐病的防控效果[J].中國農技推廣,2020,36(2):60-63.
[19]王龍根,倪玨萍,王鳳云,等.新殺菌劑JS399-19的生物活性研究[J].農藥,2004(8):380-383.
[20]祝燕麗,黃勁松,董濤海. 25%氰烯菌酯(勁護)懸浮劑浸種對稻種安全性及防病效果研究[J].現代農業科技,2012(24):141.
[21]BENINGO K A, LILLIE S H, BROWN S S. The yeast kinesin-related protein Smy1p exerts its effects on the class V myosin Myo2p via a physical interaction [J]. Molecular Biology of the Cell, 2000, 11(2): 691-702.
[22]TRYBUS K M. Myosin V from head to tail [J]. Cellular and Molecular Life Sciences, 2008, 65(9): 1378-1389.
[23]ZHENG Zhitian, HOU Yiping, CAI Yiqiang, et al. Whole-genome sequencing reveals that mutations in myosin-5 confer resistance to the fungicide phenamacril in Fusarium graminearum [J/OL]. Scientific Reports, 2015, 5: 8248. DOI: 10.1038/srep08248.
[24]高啟迅.小麥赤霉病菌對常用殺菌劑的抗性研究[D].杭州: 浙江大學,2016.
[25]原征. 小麥莖基腐病菌對咯菌腈的抗性風險評估及抗性機理研究[D].南京:南京農業大學,2020.
[26]吳志紅,汪天虹,黃衛,等.簡便易行的絲狀真菌染色體DNA提取法[J].菌物系統,2001(4):575-577.
[27]BECHER R, HETTWER U, KARLOVSKY P, et al. Adaptation of Fusarium graminearum to tebuconazole yielded descendants diverging for levels of fitness, fungicide resistance, virulence, and mycotoxin production [J]. Phytopathology, 2010, 100(5): 444-453.
[28]GARDINER D M, MCDONALD M C, COVARELLI L, et al. Comparative pathogenomics reveals horizontally acquired novel virulence genes in fungi infecting cereal hosts [J/OL]. PLoS Pathogens, 2012, 8(9): e1002952. DOI: 10.1371/journal.ppat.1002952.
[29]BOVILL W D, HORNE M, HERDE D, et al. Pyramiding QTL increases seedling resistance to crown rot (Fusarium pseudograminearum) of wheat (Triticum aestivum) [J]. Theoretical and Applied Genetics, 2010, 121(1): 127-136.
[30]周淼平,姚金保,張鵬,等.小麥抗莖腐病種質篩選及鑒定新方法的建立[J].植物遺傳資源學報,2016,17(2):377-382.
[31]ZHANG Chengqi, CHEN Yun, YIN Yanni, et al. A small molecule species specifically inhibits Fusarium myosin I [J]. Environmental Microbiology, 2015, 17(8): 2735-2746.
[32]周明國. 殺菌劑氰烯菌酯新靶標的發現及其產業化應用[EB/OL]. (2018-03-20) [2021-12-20]. https:∥kxyjy.njau.edu.cn/info/1063/6456.htm.
[33]LI B, ZHENG Z, LIU X, et al. Genotypes and characteristics of phenamacril-resistant mutants in Fusarium asiaticum [J]. Plant Disease,2016,100(8): 1754-1761.
[34]李斌. 亞洲鐮孢菌(Fusarium asiaticum)對氰烯菌酯的抗藥性遺傳多樣性及其適合度研究[D].南京:南京農業大學,2017.
[35]HOU Yiping, QU Xiangpu, MAO Xuewei, et al. Resistance mechanism of Fusarium fujikuroi to phenamacril in the field [J]. Pest Management Science, 2018, 74(3): 607-616.
(責任編輯:田?喆)
