基于4種生態位模型的長芒莧潛在適生區預測
2023-05-30 09:17:23張心怡趙健李志鵬王苗苗陳宏
植物保護 2023年2期
張心怡 趙健 李志鵬 王苗苗 陳宏
摘要 惡性入侵雜草長芒莧Amaranthus palmeri S.Watson對農業生產和生物多樣性造成嚴重威脅。預測其潛在適生區對糧食安全和生物多樣性保護至關重要。不同模型由于算法不同對長芒莧潛在適生區的預測結果存在差異。本研究綜合4種生態位模型 (MaxEnt、GARP、BIOCLIM和DOMAIN) 預測長芒莧在我國的潛在適生區以提高預測準確性。結果表明,4種模型的平均AUC值均大于0.85。MaxEnt和DOMAIN的平均Kappa值大于0.81;BIOCLIM和GARP的平均Kappa值大于0.69。MaxEnt的預測精度和穩定性要略勝一籌。BIOCLIM和MaxEnt的預測結果較為收斂,DOMAIN和GARP預測的適生區范圍較廣,由此預測的長芒莧潛在適生區分別占我國陸地總面積的20.66%和32.38%,48.39%和49.76%。綜合預測結果,長芒莧在我國的適生區主要集中在中東部地區,西北和東北地區則是長芒莧存在的邊緣環境地區。
關鍵詞 長芒莧;?潛在適生區;?生態位模型;?模型評價
中圖分類號: S 451
文獻標識碼:?A
DOI:?10.16688/j.zwbh.2022399
Abstract Amaranthus palmeri S.Watson, a malignant invasive weed, poses a serious threat to agricultural production and biodiversity. Predicting potential suitable areas is crucial for food security and biodiversity conservation. Different models give different predictions on the potential suitable areas of A.palmeri due to different algorithms. In this study, four ecological niche models (MaxEnt, GARP, BIOCLIM and DOMAIN) were used to comprehensively predict the potential suitable areas of A.palmeri in China to improve the accuracy of prediction. The results showed that the average AUC values of the four models were all greater than 0.85; the average Kappa values of MaxEnt and DOMAIN were greater than 0.81, and the average Kappa values of BIOCLIM and GARP were greater than 0.69. The prediction accuracy and stability of MaxEnt were better. In addition, the prediction results by BIOCLIM and MaxEnt were relatively convergent, while DOMAIN and GARP predicted a wider distribution of suitable areas. The predicted potential suitable areas of A.palmeri accounted for 20.66%, 32.38%, 48.39% and 49.76% of the total land area in China, respectively. Based on comprehensive prediction results, the suitable areas of A.palmeri in China were mainly concentrated in the central and eastern regions, and the northwest and northeast regions were the marginal environmental areas.
Key words Amaranthus palmeri;?potential suitable areas;?ecological niche model;?model evaluation
生物入侵是威脅生物多樣性的主要原因之一[1],入侵植物對生物多樣性的影響尤為顯著[2]。隨著全球化進程加快,跨國貿易、交通、旅游等行業的快速發展為生物入侵提供了更多樣化的途徑,生物入侵風險日趨增加、形勢日益嚴峻[3]。研究表明,全球范圍內生物入侵的最低總成本已達到約1.3萬億美元[4],生物入侵造成的損失不僅表現在經濟方面,更嚴重的是外來入侵物種會對生態環境產生不可逆轉的破壞和影響。長芒莧Amaranthus palmeri S. Watson是莧科莧屬異株莧亞屬的一種C4高光效一年生草本植物,原生于美國西南部和墨西哥西北部[5]。我國于1985年首次在北京發現長芒莧[6],目前該植物在天津、河北、河南、湖南、廣西等地都有記錄。長芒莧種子萌發率高、植株生長迅速、適應性廣,可快速積累生物量,與本土植物相比競爭優勢強[7],從而可以在侵入地成功建立種群并導致生物多樣性下降[8]和作物減產[9]。目前的防治手段如:除草劑、深耕和輪作等防控措施均無法長期有效地控制長芒莧的擴散[7,9]。相關研究表明,在雜草災害暴發前進行早期預警防控比在雜草災害暴發后根除更為經濟有效[10],而適生區的預測是做好雜草災害早期預警的重要依據和參考。
生態位模型 (ecological niche models, ENMs) 已被廣泛應用于預測物種的適生區[11-12]。ENMs基于算法運算判斷物種的生態需求,將物種分布信息和環境變量關聯,以概率的形式反映物種對生境的偏好程度[13-14]。目前常用的模型按應用方式可分為兩大類:一類基于R平臺的Biomod2程序包調用,如隨機森林 (random forest, RF)、人工神經網絡 (artificial neural network, ANN)、廣義線性模型 (generalized linear model, GLM)、廣義相加模型 (generalized additive model, GAM)等;另一類可以通過相應的軟件直接進行應用,如生物氣候分析與預測系統 (bioclimate analysis and prediction system, BIOCLIM)、DOMAIN距離模型 (domain distance, DOMAIN)、CLIMEX氣候匹配模型 (CLIMEX match climates regional, CLIMEX)、最大熵模型 (maximum entropy model, MaxEnt)、規則集遺傳算法 (genetic algorithm for rule-set production, GARP)等,這類已封裝成軟件的預測模型可操作性更強。不同模型對同一事件的分析結果不同,同一模型也會由于采用的數據、參數選擇等的不同表現出不同的結果[15]。近年來,利用多種模型綜合分析物種的適生區已成為此類研究的重要方法。
相關研究表明,在生態位保守的前提下,應用生態位模型預測物種的適生區才具有合理性[11]。李慧琪等[16]的研究指出,長芒莧在我國的生態位具有保守性,因此,選擇生態位模型預測長芒莧的適生區在方法上是可行的。目前國內外對長芒莧適生區預測的相關研究基本上是基于單一模型預測,如CLIMEX[17-18]、MaxEnt [16]和BIOCLIM [19]等,但不同模型預測結果存在較大的差異。因此,本研究通過比較4種生態位模型 (MaxEnt、GARP、BIOCLIM和DOMAIN) 對長芒莧潛在適生區預測的異同,對4種模型的預測結果進行綜合分析以提高預測的科學性,從而明確長芒莧在我國的潛在適生區,量化長芒莧在我國不同區域的風險程度,以期為長芒莧的監測預警提供具有現實意義的參考。
1?材料與方法
1.1?軟件和數據來源
1.1.1?相關軟件
所用4種生態位模型軟件分別是:MaxEnt 3.4.1用于MaxEnt模型預測、Desktop GARP 1.1.6用于GARP模型預測、DIVA-GIS 7.5用于BIOCLIM和DOMAIN模型預測。另外,DIVA-GIS 7.5、ArcGIS 10.2和IBM SPSS 21軟件用于數據處理。
1.1.2?地圖數據
世界地圖和中國地圖的分析底圖從自然資源部標準地圖服務系統 (http:∥bzdt.ch.mnr.gov.cn/) 下載。
1.1.3?分布數據
長芒莧的分布數據從多個公開的數據庫獲取,包括全球生物多樣性信息平臺 (GBIF: https:∥www.gbif.org/)、美國西南環境信息網:亞利桑那州—新墨西哥州(SEINet: https:∥swbiodiversity.org/seinet/index.php)、國際農業與生物科學研究中心(CABI: https:∥www.plantwise.org/knowledgebank/)、國家標本平臺(NSII: http:∥www.nsii.org.cn/)、中國數字植物標本館(CVH: https:∥www.cvh.ac.cn/)、中國植物圖像庫(PPBC: http:∥ppbc.iplant.cn/)。另外,還有部分數據來自文獻記錄[6,16,20-25]。
1.1.4?環境數據
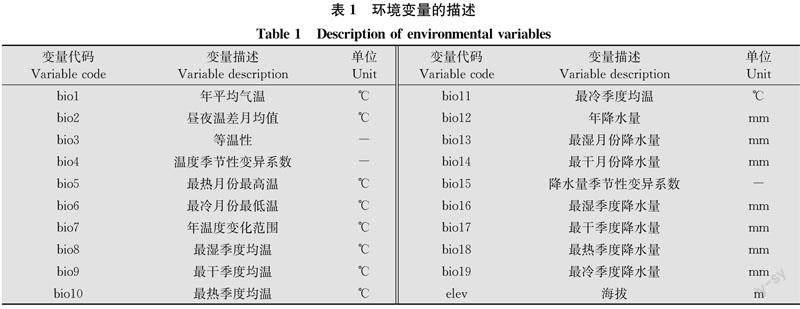
物種在較大地理空間尺度下的分布主要與氣候相關[11],而海拔高度可能通過影響溫度來影響物種的分布[26]。因此選取19個生物氣候變量和海拔作為原始環境變量(表1)。生物氣候變量包括年趨勢、季節性以及極端或限制性氣溫和降雨變化,具有更廣泛的生物學意義。從世界氣候網站 (WorldClim: https:∥www.worldclim.org/data/index.html) 下載所需數據,版本為WorldClim 2.1。為了保證較高精度和模型良好的運行能力,選擇2.5 arc minutes。
1.2?數據處理
1.2.1?分布數據
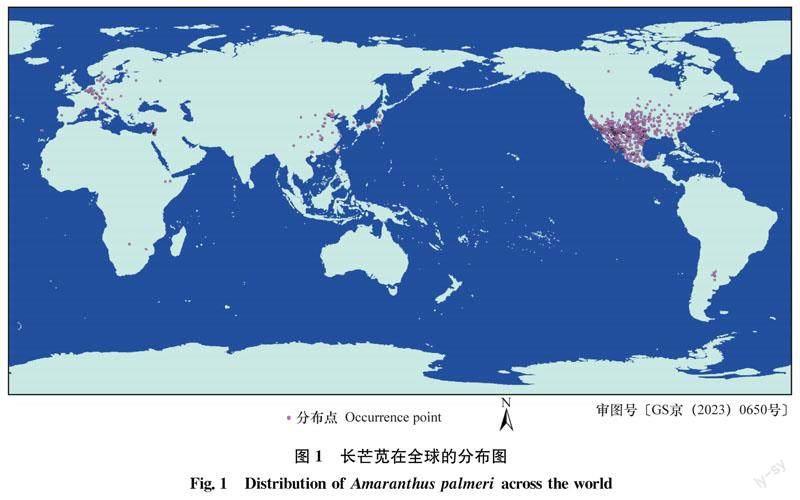
刪除原始分布數據中錯誤、重復以及十進制經緯度為整數值的數據,對于有詳細地理位置而沒有地理坐標的記錄,通過國家地理信息公共服務平臺——天地圖 (https:∥map.tianditu.gov.cn/) 確定其經緯度信息。為降低冗余數據造成的過擬合現象,參考環境變量的空間分辨率,在ArcGIS中創建2.5 minutes分辨率的漁網 (Data Management Tools-Feature Class-Create Fishnet),每個網格中僅保留一個分布點。最終獲得1 679條有效數據(圖1)。將數據保存為shp格式。
將shp格式的分布數據導入DIVA-GIS,隨機選取75%的分布數據作為訓練集用于模型預測,剩下的25%分布數據和隨機生成的1倍于分布點的非分布點作為測試集用于模型驗證,設置10次重復。最終生成一個訓練集(train.shp)和一個測試集(test.shp),包含10組相對應的訓練數據和測試數據。再將shp格式的訓練集轉換為csv格式用于MaxEnt和GARP模型預測,shp格式的訓練集用于BIOCLIM和DOMAIN模型預測,shp格式的測試集用于所有模型驗證。
1.2.2?環境變量
環境變量的原始數據為tif格式,在ArcGIS中將其轉換為asc格式。高度相關的環境變量會導致過擬合,同時降低模型運算效率,因此對環境變量進行篩選。在ArcGIS中提取分布點的生物氣候變量信息,將其導入SPSS進行雙變量相關分析,選擇Pearson相關系數對生物氣候變量進行多重共線性檢驗。當兩個變量Pearson相關系數大于0.8時,結合長芒莧的生物生理學及模型應用等研究保留一個相對更具有生物學意義的變量。
MaxEnt模型所需環境變量的格式為asc格式。在Desktop GARP中將asc格式的環境變量轉換為raw格式,再生成dxl環境數據集,用于GARP模型預測。在DIVA-GIS中將asc格式的環境變量轉換為grd格式,并創建stack環境數據集,用于BIOCLIM和DOMAIN模型預測。
1.3?模型預測
MaxEnt模型:將csv格式的訓練集和asc格式的環境變量導入軟件,利用刀切法衡量環境變量的重要性,將輸出格式設置為邏輯輸出,輸出文件類型為asc。其他參數保持默認。
GARP模型:將csv格式的訓練集和dxl格式的環境數據集導入Desktop GARP,設置100%的分布數據比例用于訓練。因為GARP模型輸出的預測結果為布爾值,所以設置20次重復。在ArcGIS中將每個訓練集的20個輸出結果疊加并取均值,轉換為[0,1]的連續概率值。
BIOCLIM和DOMAIN模型:將shp格式的訓練集導入DIVA-GIS,通過Modeling-Bioclim/Domain加載stack環境數據集,依次運行兩種模型。
1.4?模型評價
ROC曲線下的面積值AUC和Kappa統計量是模型評價常用的兩種指標,AUC的取值范圍為 [0,1],Kappa的取值范圍為 [-1,1]。在實際應用中,當AUC>0.5時,模型的預測效果優于隨機預測效果,當Kappa>0時對一致性的判斷才有意義,且AUC和Kappa值越大,說明預測的準確度越高,一致性越好[12]。由于物種分布率和診斷閾值的影響,Kappa統計值往往存在偏差,AUC值不受閾值的影響且其應用也更廣泛[27]。本研究以AUC值為主要評價指標,以Kappa輔助判斷模型的一致性。在DIVA-GIS中可以實現兩種評價指標的輸出(modeling-evaluation-create evaluation file & show ROC/Kappa)。
1.5?適生區劃分
以4種生態位模型的10組預測分布圖的均值作為最終的預測結果。由于不同模型的算法不同,尚沒有統一的適生區劃分方法。目前利用多個生態位模型進行比較的研究中,多采用自然間斷點法 (natural breaks, Jenks) 對預測結果進行重分類[28-29],所以本研究基于自然間斷點法將長芒莧在我國的潛在分布劃分為4類適生區:非適生區、低適生區、中適生區和高適生區。但是DOMAIN模型的輸出值是分類置信度而非概率估計[30],如果采用Jenks進行劃分,會造成對預測結果的過度解讀,并且利用Jenks劃分的結果與DOMAIN運行后默認輸出的適生區分布相差甚遠。DOMAIN模型需要自定義閾值把研究區域的相似性值轉化為物種的分布區并推導生成異域分布區[30-31]。本研究參考Carpenter等[31]的研究,并結合DOMAIN的輸出結果與長芒莧實際分布的匹配度,對模型運行的默認輸出結果進行調整來劃分適生區。
重分類后,通過“屬性-符號系統-唯一值”統計不同適生區的像元計數,并計算各適生區的像元占比,即為不同適生區所占中國陸地總面積的百分比。
2?結果與分析
2.1?環境變量相關性分析篩選結果
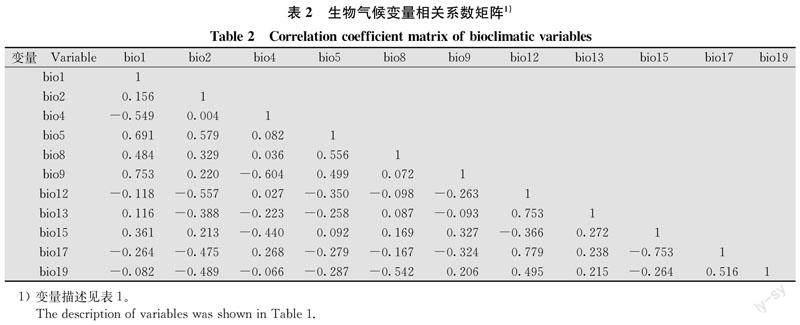
考慮到C4植物的分布與高溫密切相關[32],且長芒莧在年均溫較低的東北溫帶地區基本只進行營養生長[25],結合相關模型預測研究中年均溫對模型的貢獻較大[16,33]等信息,保留年均溫(bio1),剔除與其高度相關的最冷月最低溫(bio6)、最熱季度均溫(bio10)、最冷季度均溫(bio11),并保留與bio10高度相關的最熱月最高溫(bio5),與bio11高度相關的最干季度均溫(bio9)。基于長芒莧種子在成熟后一年內對發芽溫度要求不同,且發芽能力受到溫度季節性的顯著影響[34],保留溫度季節性變異系數(bio4),剔除與之高度相關的等溫性(bio3)、溫度年變化范圍(bio7)。
與降水相關的環境變量中僅有兩組變量表現出高度相關,即最濕月降水量(bio13)、最濕季度降水量(bio16)和最熱季度降水量(bio18)兩兩相關;最干月降水量(bio14)和最干季度降水量(bio17)高度相關。長芒莧起源于索諾拉沙漠,其出現往往與干旱環境有關,且作為機會主義者,可以快速響應環境中可利用的水分[35-36]。有研究表明在高濕度條件下,觀察到長芒莧種子上有真菌群落的存在,這可能導致種子腐爛,活性降低,從而影響長芒莧的發生[37]。因此,結合相關性分析結果,保留bio13和bio17。
保留所有與其他變量不具有高度相關性的變量。本研究對生物氣候變量的相關性分析篩選結果如表2所示。最終保留11個生物氣候變量,并添加海拔變量作為地形因子。
2.2?4種生態位模型評估
4種模型的AUC和Kappa值如表3所示。對4種模型的AUC和Kapp值分別進行方差分析,由于原始AUC數據不符合Levene方差齊性,所以對AUC值進行反正弦平方根轉換,轉換后的數據符合方差齊性 P (AUC)=0.322, P (Kappa)=0.567,顯著性均大于0.05。由LSD多重比較結果可知,MaxEnt和DOMAIN模型的AUC (P=0.106) 和Kappa (P=0.981)無顯著性差異,GARP和BIOCLIM 模型的Kappa (P=0.714) 無顯著差異。AUC和Kappa的平均值均表現為:MaxEnt>DOMAIN>BIOCLIM>GARP。AUC的標準偏差:MaxEnt<DOMAIN<BIOCLIM<GARP,Kappa 的標準偏差:MaxEnt<BIOCLIM<GARP<DOMAIN。綜合結果,MaxEnt和DOMAIN模型的預測準確度要優于BIOCLIM和GARP,并且MaxEnt模型較其他3種模型略勝一籌。
2.3?長芒莧在我國的潛在適生區
利用4種生態位模型對長芒莧在我國的潛在分布預測結果如圖2所示,適生區面積占比如圖3所示。MaxEnt預測結果顯示,長芒莧在我國的適生區主要集中在中東部地區,高度適生區集中在華北平原,低、中、高適生區分別占我國陸地總面積的13.00%、11.45%、7.93%。GARP預測結果顯示長芒莧分布較廣且我國中東部地區均表現為高度適生區,此外西北地區也有一定的適生區分布,低、中、高適生區分別占我國陸地總面積的8.48%、7.30%、33.98%。BIOCLIM的預測結果最為收斂,適生區主要分布在22°N~40°N,98° E~122°E之間,高度適生區僅分布在河南大部分地區以及安徽、湖北和陜西部分地區,低、中、高適生區分別占我國陸地總面積的12.69%、5.68%、2.29%。DOMAIN的預測適生區范圍最廣,但相比于GARP可以更好地擬合適生區的分布,我國中東部、西北以及東北地區均表現出一定的適生性,高度適生區較分散,低、中、高適生區分別占我國陸地總面積的28.90%、15.21%、4.28%。綜合結果,GARP和DOMAIN模型預測的適生區分布較廣,MaxEnt和BIOCLIM預測的適生區分布較為收斂。4種模型顯示共同的適生區集中在我國中東部。
2.4?影響長芒莧分布的主導環境變量
MaxEnt模型可以通過刀切法衡量環境變量的相對重要性,環境變量的相對貢獻百分比如表4所示,各環境變量的正規化訓練增益值如圖4所示。結果顯示,與溫度相關的氣候因子對長芒莧的分布起主導作用,相對貢獻占所有環境變量的77.21%,其中僅年均溫(bio1)就占41.37%,晝夜溫差月均值(bio2)和溫度季節性變異系數(bio4)也對長芒莧的分布有一定的影響。由圖4可知,當剔除最冷季度降水量(bio19)時,正規化訓練增益值下降最多,說明其可能包含其他變量沒有的信息。
3?討論
3.1?4種生態位模型的適用性評估
根據方差分析和LSD多重比較,MaxEnt與DOMAIN模型的預測精度沒有顯著差異,但MaxEnt的AUC和Kappa平均值略高、標準偏差較小,模型更穩定。雖然GARP和BIOCLIM模型的預測精度較低,但也具有一定的參考價值。另外,在運行效率方面,MaxEnt具有絕對的優勢,運行速度快且不容易出錯;DOMAIN運行速度慢且容易出現卡頓現象;BIOCLIM表現一般;GARP的結果處理極為繁瑣。因此,研究以MaxEnt和DOMAIN為主,GARP和BIOCLIM為輔,綜合分析長芒莧在我國的適生區。
本研究所用的4種模型都可以通過對應的軟件直接運行,操作界面簡潔明了。一方面,MaxEnt、DOMAIN、BIOCLIM模型的輸出結果所占內存均在5 GB以下,而GARP模型由于需要進行布爾值到邏輯值的轉換,設置了每組數據20次重復,導致最終所占內存高達40 GB。另一方面,僅MaxEnt模型在預測時可以設置利用刀切法衡量環境變量的重要性并直接輸出結果,DOMAIN、BIOCLIM和GARP模型雖然可以通過對單個變量或者多個變量組合進行預測,但需要耗費較長的時間,不建議在較大地理空間尺度下預測時進行這樣的操作。因此,在較大地理空間尺度下設置較多重復時,應優先考慮MaxEnt、DOMAIN和BIOCLIM模型,建議進行預運行(如先設置1次重復)估計所需內存大小,以避免內存不夠導致任務中斷或失敗。另外,當需要分析主導環境變量時,建議優先使用MaxEnt模型。
3.2?4種生態位模型預測結果比較
對于預測結果較為收斂的MaxEnt和BIOCLIM模型來說,MaxEnt可以更好地擬合長芒莧在我國的分布,而BIOCLIM模型對長芒莧實際分布的擬合較差。長芒莧已經擴散到北京市的多個區,但BIOCLIM的預測結果顯示北京為非適生區。BIOCLIM將環境包絡 (environmental envelope) 定義為歐幾里得空間 (Euclidean space) 中的直線體積,基于范圍獨立處理每個氣候軸,這可能會導致生態不合理的預測[31]。GARP模型預測的長芒莧高適生區范圍最大。在對金錢松Pseudolarix amabilis[28]和斑體花蜱Amblyomma maculatum[38]等物種的適生區預測中,GARP也表現出相似的預測結果。這可能是因為GARP的輸出結果為布爾型,只要長芒莧在某地表現出一定的適生性,結果即為適生;也可能是由于重復設置不夠而導致分類不明顯。然而GARP預測的低、中適生區也恰恰反映了長芒莧存在的邊緣環境條件。DOMAIN模型預測的適生區范圍也較大,這是因為DOMAIN模型通過Gower算法計算點與點的相似性,利用相似矩陣對研究區域進行賦值和分類[31],受分布點影響較大,只要某地區有實際分布,基本能表現出一定范圍的適生區[29]。本研究的取樣點包括了我國西北地區長芒莧的記錄(甘肅和內蒙古,數據分別來源于PPBC和CVH),其他學者在東北溫帶地區觀察到長芒莧的發生[25]也進一步說明了長芒莧在我國西北和東北地區存在一定的適生性。
3.3?環境變量對長芒莧分布的影響
研究結果顯示長芒莧的分布主要受溫度影響,尤其是年均溫。結合本文2.1對溫度相關變量進行篩選的信息可知,年均溫(bio1)可能是限制長芒莧在高緯度地區擴散的主要因子。溫度季節性變異系數(bio4)也對長芒莧的發生有一定的影響。有研究表明,相較于恒溫,長芒莧在交替溫度下的發芽率更高,且隨著一定范圍內溫度的升高,發芽率也升高[39-40]。我國由南向北、由東向西,晝夜溫差逐漸變大。當單獨考慮晝夜溫差時,長芒莧在我國西北地區應表現出較高的適生性。預測結果顯示長芒莧在我國的潛在適生區主要集中在中東部,說明晝夜溫差可能與其他環境變量共同作用影響長芒莧的分布,對長芒莧的分布起一定的“修飾”作用而非主導作用。結合本文2.1對降雨相關變量進行篩選的信息可知,長芒莧的萌發生長對土壤水分有一定要求,最冷季度降水量(bio19)可能通過影響土壤含水量進一步影響長芒莧的發生,從而導致最冷季度降水量對長芒莧分布有一定貢獻。
長芒莧的地理分布橫跨熱帶和溫帶氣候區域,具有較強的氣候適應性。長芒莧在壓力條件下的高度可塑性[41]和廣泛的環境適應性[25]可以使其在邊緣條件下持續存在。而未來氣候變化下,溫度逐漸升高,長芒莧有極大可能向高緯度擴張并建立更廣泛的繁殖種群。
3.4?前景與展望
雖然長芒莧已經在我國多個省份發生,但已有分布數據記錄卻很少。一方面可能是某地發現長芒莧后及時根除后未記錄,另一方面可能是人們對于長芒莧的認識和關注度不夠,無相關發生記錄。此外,數據更新和共享不及時導致信息滯后也是一個原因。我國進境口岸于2010年首次報道檢出長芒莧,從收集到的分布數據來看,在2010年之前長芒莧僅在北京市有分布記錄。2010年,我國口岸相關單位開展異株莧亞屬雜草的檢疫鑒定技術培訓,此后,長芒莧的檢出次數近乎呈直線上升[42],其他省份也逐漸有長芒莧的分布記錄。因此,加強長芒莧等重要入侵物種的識別、鑒定技能培訓與公眾科普,提高基層人員對重要入侵物種,特別是新發入侵物種的認知和處置能力,對于加強我國入侵防控工作具有重要的現實意義。
目前,長芒莧在我國尚未大面積入侵為害,但長芒莧極易混雜在收獲的作物中并隨糧食調運進行遠距離傳播,擴散風險較高。我國是農業生產大國,也是生物多樣性最為豐富的國家之一,一旦長芒莧在多地大面積暴發,將嚴重威脅我國糧食安全和生態安全。準確評估長芒莧在我國的潛在適生區對于不同地區制定長芒莧預警和防控策略有較強的指導意義。長芒莧在我國京津冀地區呈暴發性增長,這些地區是長芒莧發生的高風險區,應積極采取防除措施阻止其進一步擴散暴發。在河南、湖北、湖南、廣西、廣東、浙江、江蘇等地區發現長芒莧有零星分布記錄,在山西、陜西、江西、上海等地區暫時未發現長芒莧的分布記錄,但預測結果顯示,長芒莧在這些地區表現出不同程度的適生性,有不同程度的入侵風險,建議開展普查工作摸清長芒莧的實際分布情況并建立早期監測和預警體系。同時,構建對疫情的快速響應和實時控制體系以及加強長芒莧的科普宣傳也有利于“早發現,早預警,早應對”。
4?結論
本研究利用長芒莧在全球的分布記錄和結合生物學相關特性篩選的環境變量,基于MaxEnt、GARP、BIOCLIM和DOMAIN模型預測了長芒莧在我國的潛在適生區。綜合模型評價指標和模型運行性能,MaxEnt最優,DOMAIN稍遜,BIOCLIM和GARP次之。4種模型預測的適生區重合區域集中在我國中東部地區,而西北和東北地區則是長芒莧在我國發生的邊緣環境地區。本研究采用4種模型和2種評價指標,不同模型之間進行互補,較單一模型預測結果更為可靠,可為長芒莧早期預警與防控決策提供參考。
參考文獻
[1]?SEEBENS H, BLACKBURN T M, DYER E E, et al. Global rise in emerging alien species results from increased accessibility of new source pools [J]. Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(10): E2264-E2273.
[2]?鞠瑞亭, 李慧, 石正人, 等. 近十年中國生物入侵研究進展[J]. 生物多樣性, 2012, 20(5): 581-611.
[3]?謝聯輝, 尤民生, 侯有明, 等. 生物入侵——問題與對策[M]. 北京:科學出版社, 2011: 6.
[4]?DIAGNE C, LEROY B, VAISSIERE A C, et al. High and rising economic costs of biological invasions worldwide [J]. Nature, 2021, 592(7855): 571-576.
[5]?SAUER J. Recent migration and evolution of the dioecious Amaranths [J]. Evolution, 1957, 11(1): 11-31.
[6]?李振宇. 長芒莧——中國莧屬一新歸化種[J]. 植物學通報, 2003(6): 734-735.
[7]?ROBERTS J, FLORENTINE S. A review of the biology, distribution patterns and management of the invasive species Amaranthus palmeri S.Watson (Palmer amaranth): Current and future management challenges [J]. Weed Research, 2021, 62(2): 113-122.
[8]?LI Yiming, MO Xunqiang, LI Qiannan, et al. Effects of an invasive plant Palmer amaranth (Amaranthus palmeri) on herb communities [J/OL]. IOP Conference Series: Earth and Environmental Science, 2021, 791(1): 012202. DOI: 10.1088/1755-1315/791/1/012202.
[9]?WARD S M, WEBSTER T M, STECKEL L E. Palmer amaranth (Amaranthus palmeri): a review [J]. Weed Technology, 2013, 27(1): 12-27.
[10]JARNEVICH C S, HOLCOMBE T R, BARNETT D T, et al. Forecasting weed distributions using climate data: a GIS early warning tool [J]. Invasive Plant Science and Management, 2010, 3(4): 365-375.
[11]PHILLIPS S J, ANDERSON R P, SCHAPIRE R E. Maximum entropy modeling of species geographic distributions [J]. Ecological Modelling, 2006, 190(3): 231-259.
[12]JIMENEZ-VALVERDE A, PETERSON A T, SOBERON J, et al. Use of niche models in invasive species risk assessments [J]. Biological Invasions, 2011, 13(12): 2785-2797.
[13]朱耿平, 劉國卿, 卜文俊, 等. 生態位模型的基本原理及其在生物多樣性保護中的應用[J]. 生物多樣性, 2013, 21(1): 90-98.
[14]許仲林, 彭煥華, 彭守璋. 物種分布模型的發展及評價方法[J]. 生態學報, 2015, 35(2): 557-567.
[15]FANG Yaqin, ZHANG Xuhui, WEI Haiyan, et al. Predicting the invasive trend of exotic plants in China based on the ensemble model under climate change: a case for three invasive plants of Asteraceae [J/OL]. Science of the Total Environment, 2021, 756: 143841. DOI: 10.1016/j.scitotenv.2020.143841.
[16]李慧琪, 趙力, 祝培文, 等. 入侵植物長芒莧在中國的潛在分布[J]. 天津師范大學學報(自然科學版), 2015, 35(4): 57-61.
[17]徐晗, 宋云, 范曉虹, 等. 3種異株莧亞屬雜草入侵風險及其在我國適生性分析[J]. 植物檢疫, 2013, 27(4): 20-23.
[18]KISTNER E J, HATFIELD J L. Potential geographic distribution of Palmer amaranth under current and future climates [J/OL]. Agricultural & Environmental Letters, 2018, 3(1): 170044. DOI: 102134/ae12017.12.0044.
[19]趙光明. 連云港口岸進口植物產品截獲有害生物分析及重要有害生物風險分析[D]. 揚州: 揚州大學, 2018.
[20]李象欽, 韋春強, 潘玉梅, 等. 廣西三種新記錄歸化植物及其入侵性分析[J]. 廣西植物, 2017, 37(6): 806-810.
[21]吳堯晶, 周柳, 肖順勇, 等. 湖南省6種新記錄外來植物及其入侵性分析[J]. 湖南師范大學自然科學學報, 2018, 41(3): 25-29.
[22]莫訓強, 孟偉慶, 李洪遠. 天津3種外來植物新記錄——長芒莧、瘤梗甘薯和鉆葉紫菀[J]. 天津師范大學學報(自然科學版), 2017, 37(2): 36-38.
[23]陸麗華, 周慶, 劉生峰, 等. 重慶口岸4種重要外來入侵植物的鑒定[J]. 時代農機, 2017, 44(12): 150-151.
[24]張梅. 二種莧屬入侵植物對本地植物功能性狀的生態影響研究[D]. 天津: 南開大學, 2020.
[25]曹晶晶, 王瑞, 李永革, 等. 外來入侵植物長芒莧在中國不同地區的表型變異與環境適應性[J]. 植物檢疫, 2020, 34(3): 25-31.
[26]AZRAG A G A, PIRK C W W, YUSUF A A, et al. Prediction of insect pest distribution as influenced by elevation: Combining field observations and temperature-dependent development models for the coffee stink bug, Antestiopsis thunbergii (Gmelin) [J/OL]. PLoS ONE, 2018, 13(6): e0199569. DOI: 10.1371/journal.pone.0199569.
[27]王運生, 謝丙炎, 萬方浩, 等. ROC曲線分析在評價入侵物種分布模型中的應用[J]. 生物多樣性, 2007(4): 365-372.
[28]王國崢, 耿其芳, 肖孟陽, 等. 基于4種生態位模型的金錢松潛在適生區預測[J]. 生態學報, 2020, 40(17): 6096-6104.
[29]張琴, 張東方, 吳明麗, 等. 基于生態位模型預測天麻全球潛在適生區[J]. 植物生態學報, 2017, 41(7): 770-778.
[30]張雷, 劉世榮, 孫鵬森, 等. 基于DOMAIN和NeuralEnsembles模型預測中國毛竹潛在分布[J]. 林業科學, 2011, 47(7): 20-26.
[31]CARPENTER G, GILLISON A N, WINTER J. DOMAIN: a flexible modeling procedure for mapping potential distributions of plants and animals [J]. Biodiversity Conservation, 1993, 2(6): 667-680.
[32]EHLERINGER J R, CERLING T E, HELLIKER B R. C4 photosynthesis, atmospheric CO2, and climate [J]. Oecologia, 1997, 112: 285-299.
[33]RUNQUIST R D B, LAKE T, TIFFIN P, et al. Species distribution models throughout the invasion history of Palmer amaranth predict regions at risk of future invasion and reveal challenges with modeling rapidly shifting geographic ranges [J/OL]. Scientific Reports, 2019, 9: 2426. DOI: 10.1038/s41598-018-38059-9.
[34]JHA P, NORSWORTHY J K, RILEY M B, et al. Annual changes in temperature and light requirements for germination of Palmer amaranth (Amaranthus palmeri) seeds retrieved from soil [J]. Weed Science, 2010, 58(4): 426-432.
[35]EHLERINGER J. Ecophysiology of Amaranthus palmeri, a sonoran desert summer annual [J]. Oecologia, 1983, 57(1/2): 107-112.
[36]PISKACKOVA T A R, REBERRG-HORTON S C, RICHARDSON R J, et al. Windows of action for controlling Palmer amaranth (Amaranthus palmeri) using emergence and phenology models [J]. Weed Research, 2021, 61(3): 188-198.
[37]FRANCA L X, DODDS D M, REYNOLDS D B, et al. Influence of flooding period and seed burial depth on Palmer amaranth (Amaranthus palmeri) seed germination [J]. Pest Management Science, 2020, 76(11): 3832-3837.
[38]馬德龍, 李超, 周若冰, 等. 基于規則集遺傳算法模型的斑體花蜱在中國適生區預估[J]. 中國媒介生物學及控制雜志, 2022, 33(2): 262-267.
[39]STECKEL L E, SPRAGUE C L, STOLLER E W, et al. Temperature effects on germination of nine Amaranthus species [J]. Weed Science, 2004, 52: 217-221.
[40]吉美靜, 崔海蘭, 陳景超, 等. 影響長芒莧種子萌發因素的研究[J]. 雜草學報, 2021, 39(3): 8-14.
[41]KORRES N E, NORSWORTHY J K. Palmer amaranth (Amaranthus palmeri) demographic and biological characteristics in wide-row soybean [J]. Weed Science, 2017, 65(4): 491-503.
[42]楊靜, 伏建國, 廖芳, 等. 全國口岸近年來截獲莧屬雜草疫情分析[J]. 植物檢疫, 2015, 29(3): 93-96.
(責任編輯:楊明麗)
