
油棕C2H2基因家族鑒定及低溫脅迫下的表達分析
2023-05-30 23:09:04程秋如劉子凡曾憲海
熱帶作物學報 2023年3期
程秋如 劉子凡 曾憲海

關鍵詞:油棕;C2H2 型鋅指蛋白;基因家族;低溫脅迫
中圖分類號:S565.9 文獻標識碼:A
鋅指蛋白(zink finger protein, ZFP)是一類廣泛存在于植物中的轉錄因子,最早在非洲爪蟾的卵母細胞中發現,具有手指狀結構,通過與Zn2+結合來穩定此結構[1]。根據鋅指蛋白二級結構中組氨酸(H)和膀胱氨酸(C)的數目和位置,可將其劃分為C2H2、C3H、C2C2、C4、C6、C8、C2HC、C2HC5 等類型[2]。C2H2 型鋅指蛋白轉錄因子是目前研究最多的鋅指蛋白,廣泛存在于真核生物中, C2H2 型鋅指蛋白中含有特殊的X2-Cys-X(2-4)-CysX12-His-X(3-5)-His 保守結構域,其中C 表示半胱氨酸,H 表示組氨酸,X 表示任意氨基酸,P 表示苯丙氨酸,L 表示亮氨酸[3-4]。多數C2H2 鋅指蛋白含有植物特有的一段高度保守的QALGGH 基序,該基序對DNA 的結合活性至關重要,C2H2 型鋅指蛋白不僅能與DNA、RNA結合還可以與蛋白質相互作用,在植物的轉錄調控、RNA 代謝等途徑中發揮重要功能[5-8]。
C2H2型鋅指蛋白最早在矮牽牛中發現[9],隨后在其他植物的研究中逐漸增多,如擬南芥[10],水稻[11],楊樹[12],煙草[13],小麥[14],番茄[15]、木薯[16],馬鈴薯[17],大麻[18]等,這些都證明C2H2型鋅指蛋白參與了植物的營養生長、生殖發育及對逆境脅迫的響應。C2H2 型鋅指蛋白在植物的抗寒中發揮重要作用。SAKAMOTO 等[7]研究發現擬南芥C2H2 家族的STZ、AZF1 和AZF3 基因在低溫脅迫下被顯著誘導表達。過表達ZAT12 基因可以有效的提高轉基因擬南芥的耐低溫能力[19]。ZHANG 等[20] 研究發現在擬南芥中過表達SlCZFP1 基因可以增強轉基因擬南芥的耐低溫能力。在枳中發現PtrZPT2-1 基因在低溫脅迫下表達量顯著升高,且過表達PtrZPT2-1 基因的轉基因煙草表現出較強的抗寒性[21]。在大豆中發現C2H2 型鋅指蛋白家族的SCOF-1 基因可以誘導下游COR 基因的表達,可以增強轉基因擬南芥的抗寒能力[22]。矮牽牛中ZPT2-2 基因可以增強轉基因植物的耐低溫能力[23]。在煙草中轉OSISAP1 基因可以提高轉基因煙草的抗寒能力[24]。在水稻中ZFP245、ZFP18 基因在低溫下被顯著誘導表達[25],過表達OsZFP151 可以增強轉基因水稻的耐低溫能力[26],上述研究結果說明,C2H2 基因在響應植物的低溫脅迫過程中發揮著積極作用,而油棕C2H2 型鋅指蛋白家族是否參與低溫脅迫的應答尚不明析。因此,在油棕基因組中鑒定C2H2 型鋅指蛋白并明確其在低溫脅迫中的功能對油棕抗寒研究是非常重要的。
油棕(Elaeis guineensis Jacq.)是一種重要的熱帶木本油料作物,也是世界上產油效率最高的油料作物,年均產油量高達4.27 t/hm2,被譽為“世界油王”[27]。油棕原產于熱帶非洲,喜高溫多雨的氣候,適宜在年均氣溫為25~28 ℃的地區生長[28],我國熱區地處熱帶北緣,年平均氣溫19.0~25.4 ℃,極端最低溫達0~5 ℃,熱量低于油棕原產區,嚴重影響了油棕的生長發育和產量[29]。本研究以擬南芥C2H2 氨基酸序列作為探針,在油棕基因組數據庫中進行本地BLASTP 比對,篩選出油棕C2H2 型鋅指蛋白家族,對其進行生物信息學分析,并根據轉錄組數據和qRT-PCR 方法分析油棕C2H2 家族基因在低溫脅迫下的表達情況,以期為后續油棕C2H2 家族基因的低溫脅迫研究提供理論依據。
1 材料與方法
1.1 材料
供試材料為本課題組前期篩選的云南大理(東經100°72′,北緯25°50′,海拔1381 m,年均溫14.9 ℃,極端最低溫–3.3~2.3 ℃[30])抗寒高產油棕成齡樹的自交子一代耐寒材料和冷敏感材料各3 株[31],苗齡約為3 年,現存葉片數為5~10片,株高100~150 cm。
將耐寒材料和冷敏感材料放置于中國熱帶農業科學院橡膠研究所冷庫中進行4 ℃低溫處理(每天光照時間12 h),分別在低溫處理0、1、3、5 d 時進行取樣。采集植株第2~3 片葉葉軸中部兩邊的小葉作為樣品,將葉樣用去離子水洗凈后再用錫箔紙嚴密包裹然后放到液氮中速凍,最后轉移到–80 ℃超低溫冰箱保存,用于總RNA 提取,每個處理3 個重復。
本研究將耐寒材料命名為CT(cold tolerance),冷敏感材料命名為CS(cold sensitive),CT 低溫處理0、1、3、5 d 的樣品命名為CT0、CT1、CT3、CT5,CS 低溫處理0、1、3、5 d 的樣品命名為CS0、CS1、CS3、CS5,每個處理3個重復。
1.2 方法
1.2.1 油棕C2H2 家族成員鑒定及理化性質分析從 NCBI(https://www.ncbi.nlm.nih.gov/)網站下載油棕基因組序列和基因組蛋白質序列,從TAIR數據庫中下載176 個擬南芥C2H2 型鋅指蛋白家族氨基酸序列,以擬南芥C2H2 氨基酸序列作為探針,在油棕基因組數據庫中進行本地BLASTP比對,閾值設置為1e–5,獲得與擬南芥C2H2 型鋅指蛋白相似度高的油棕蛋白質全長序列。將獲得的油棕C2H2 基因序列在pfam(http://pfam.xfam.org/)、Batch Web CD-Search Tool(https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi ) 和SMART9.0(http://smart.embl-heidelberg.de/)等在線軟件進行結構域驗證,篩選含有C2H2 結構域的蛋白序列確定為油棕C2H2 型鋅指蛋白家族成員。在ExPASy 網站( http://web.expasy.org/protparam/)對獲得的油棕C2H2 型鋅指蛋白家族成員進行氨基酸數、分子量(MW)、等電點(pI)、不穩定系數等理化性質分析。使用WOLFPSORT網站(http://www.genscript.com/wolf-psort.html)進行亞細胞定位。
1.2.2 系統進化樹的構建及染色體定位 將獲得的油棕和擬南芥的C2H2 型鋅指蛋白序列使用MEGA7.0 軟件基于NJ(Neighbor join)法構建系統進化樹,bootstrap 值為1000 次。以油棕基因組注釋信息為依據,使用TBtools 軟件[33]進行油棕C2H2 鋅指蛋白家族基因的染色體定位。
1.2.3 基因結構與保守基序分析 使用在線軟件GSDS2.0(http://gsds.gao-lab.org/)進行基因結構分析,并使用TBtools 軟件[32]進行可視化圖片的輸出。保守基序使用在線軟件MEME(http://meme-suite.org/)分析,并使用TBtools 軟件[32]進行可視化圖片的輸出。
1.2.4 啟動子順式元件分析 使用 TBtool 軟件[32]提取油棕C2H2 型鋅指蛋白家族成員的啟動子序列, 使用PlantCARE ( http://bioinformatics.psb.ugent.be/webtools/plantcare/htm)在線網站進行順式元件預測,并使用TBtools 軟件[32]進行可視化圖片的輸出。
1.2.5 低溫脅迫下EgC2H2 基因家族成員的表達分析 根據前期進行轉錄組測序(未發表數據)的結果,從轉錄組測序數據中提取油棕C2H2 型鋅指蛋白家族成員的FPKM 值,利用TBtools 軟件[32]繪制熱圖。
根據轉錄組數據分析篩選16 個顯著差異表達的EgC2H2 基因進行qRT-PCR 分析。采用RNAprep Pure 多糖多酚植物總RNA 提取試劑盒提取RNA,用瓊脂糖凝膠電泳檢測RNA 質量,使用Agilent2100 對RNA 的濃度和A260/A280 進行檢測。使用thermo scientific RevertAid First StrandcDNA Kit 試劑盒進行cDNA 反轉錄。使用CFX96Real-time PCR System(Bio-Rad, United States)儀器進行qRT-PCR 驗證,每個樣品3 個重復,反應體積為20 μL,包括ChamQ Universal SYBR qPCRMaster Mix 10.0 μL、正向及反向引物(10 umol/L)各0.4 μL、cDNA(10 ng/L)1.0 μL 以及ddH2O8.2 μL。qRT-PCR 反應程序:95 ℃預變性30 s,95 ℃變性5 s,60 ℃退火延伸30 s,40 個循環。Egβ-actin 為內參基因[33],利用NCBI 中的primer-BLAST 設計特異性引物,并由擎科生物科技有限公司海南分公司合成,引物序列見表1。采用2–CT 方法,計算基因的表達量,使用Originpro2018 軟件進行繪圖。
1.3 數據處理
使用DPS 軟件進行數據分析,用Duncans法進行差異顯著性分析。
2 結果與分析
2.1 油棕C2H2家族成員鑒定及理化性質分析
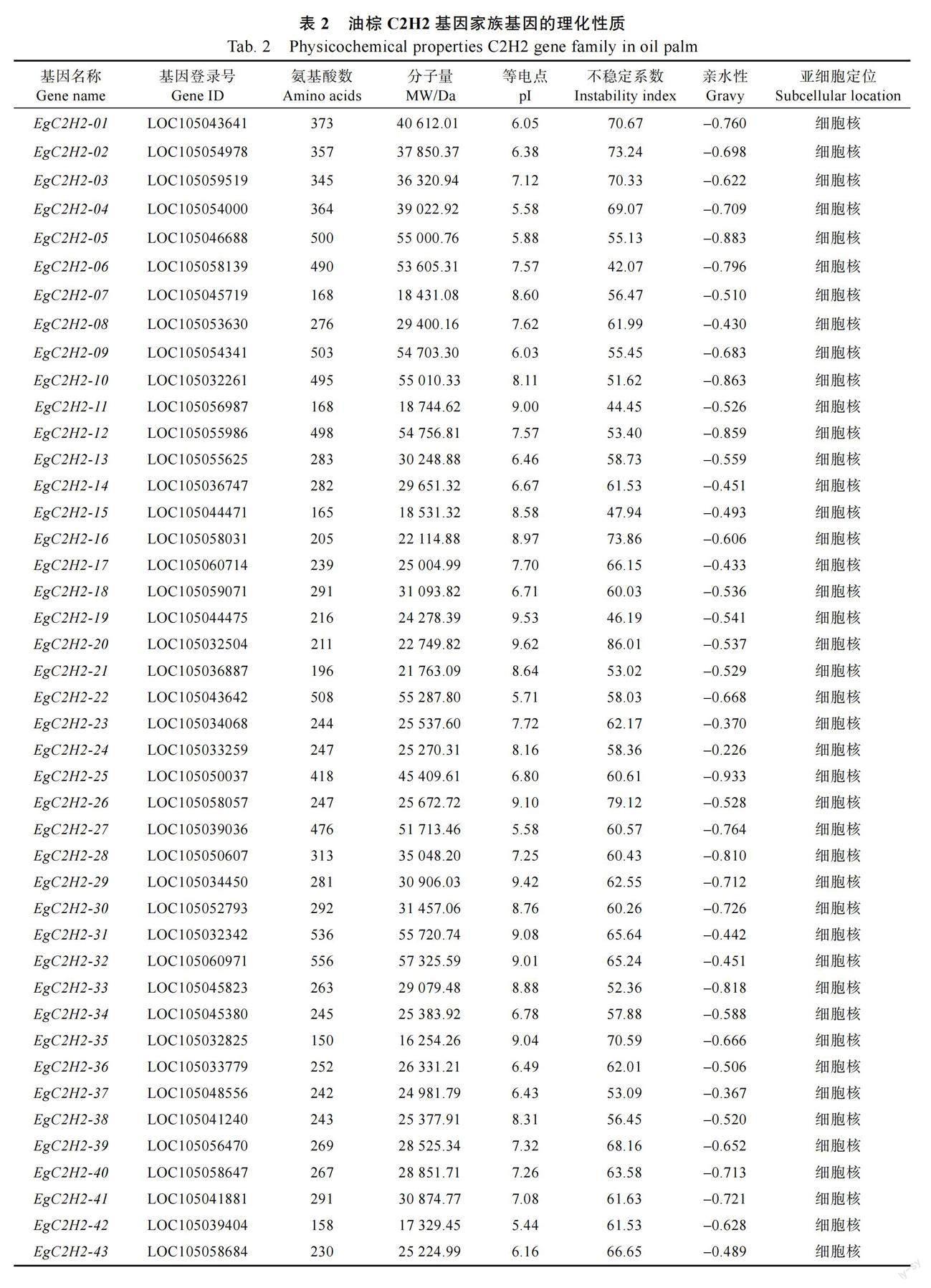
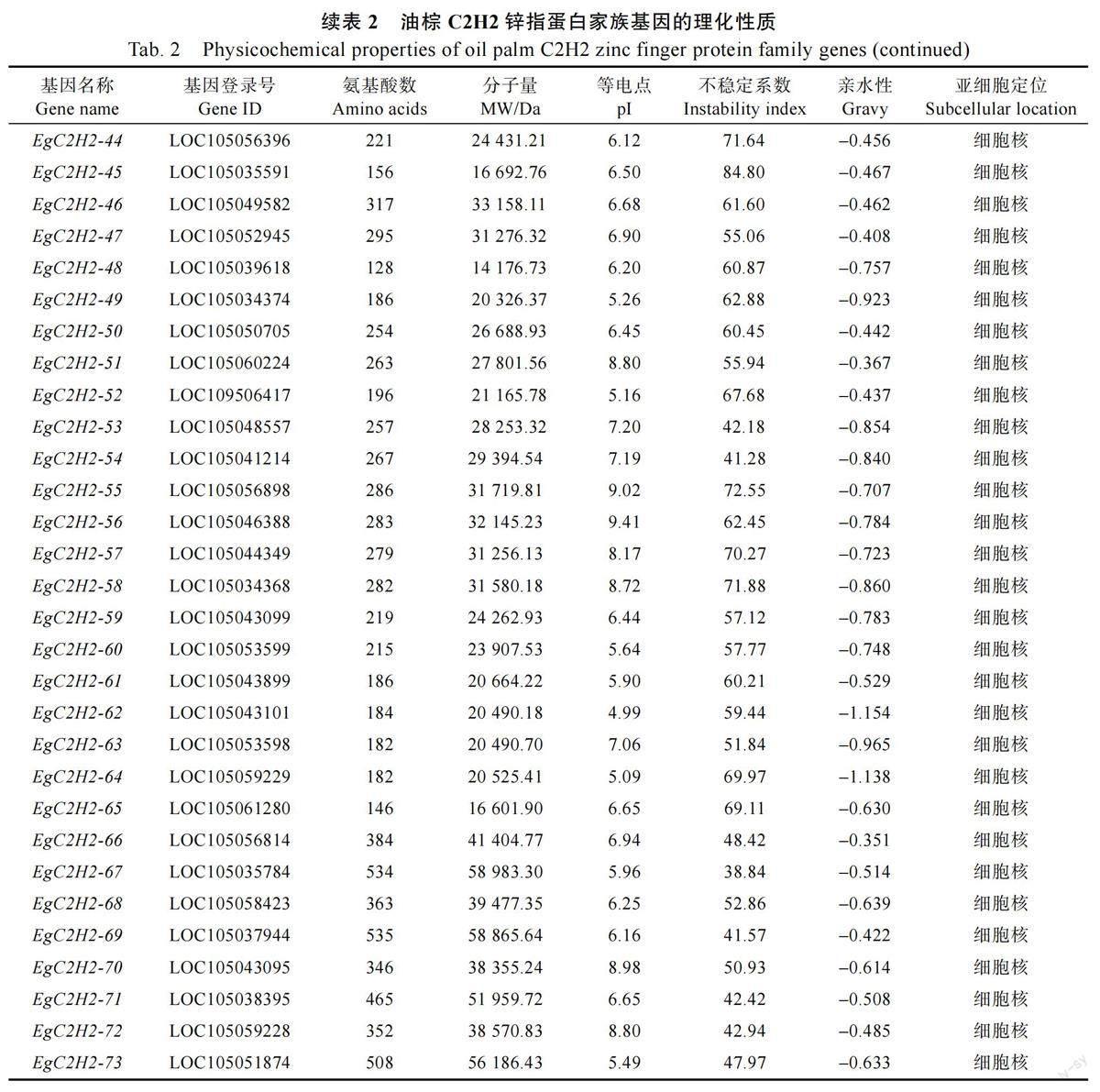
根據擬南芥C2H2 氨基酸序列在油棕基因組數據庫中進行BLASTP 比對、篩選和驗證,共獲得73 個油棕C2H2 型鋅指蛋白家族成員,并將其命名為EgC2H2-01~EgC2H2-73,如表2。在油棕C2H2 基因編碼的蛋白質中,氨基酸的數目為128~556 個,分子量為14 176.73~58 983.30 Da,等電點為4.99~9.62,不穩定系數為38.84~86.01,除EgC2H2-67 屬于穩定蛋白,其他均為不穩定蛋白,親水性為-1.154~-0.226,亞細胞定位顯示C2H2 基因家族所有成員均定位于細胞核。
2.2 C2H2 家族成員進化樹分析
利用MEGA7.0 軟件對油棕和擬南芥C2H2 氨基酸序列構建系統進化樹,根據進化樹分析結果(圖1),可以將C2H2 基因分成4 個亞家族,分別命名為Ⅰ~Ⅳ。擬南芥和油棕C2H2 基因在4 個亞家族中均有分布,第Ⅰ亞家族C2H2 基因最多,有59 個成員,第Ⅱ亞家族有56 個成員,第Ⅲ亞家族有12 個成員,第Ⅳ亞家族最少,有9個成員。油棕C2H2 與擬南芥C2H2 發生明顯聚類,表明C2H2 型鋅指蛋白在進化上具有保守性。
2.3 油棕C2H2 基因家族染色體定位
根據油棕基因組注釋文件,利用TBtools 軟件對油棕C2H2 家族基因的染色體位置進行可視化圖片的輸出。如圖2 所示,73 個油棕C2H2 家族中的55 個基因不均勻的分布于16 條染色體上,其中Chrom04、Chrom05 和Chrom11 染色體上分布的基因數目最多,均分布有6 個EgC2H2 基因,Chrom02 和Chrom15 染色體上均分布5 個EgC2H2基因,Chrom08、Chrom13 和Chrom16 染色體上均分布有4 個EgC2H2 基因,Chrom03 染色體上分布有3 個EgC2H2 基因,Chrom01、Chrom07、Chrom10、Chrom12 和Chrom14 染色體上均分布有2 個EgC2H2 基因,Chrom06 和Chrom09 染色體上基因數目最少,均分布有1 個EgC2H2 基因,18 個EgC2H2 基因未確定染色體位置。
2.4 油棕C2H2 基因家族基因結構分析
本研究對EgC2H2 家族基因結構進行了分析(圖3),73 個EgC2H2 基因含有1~4 個外顯子,EgC2H2-42 和EgC2H2-66 含有2 個外顯子,EgC2H2-19 和EgC2H2-33 含有3 個外顯子,EgC2H2-31 、EgC2H2-32 、EgC2H2-53 和EgC2H2-54 含有4 個外顯子,其他EgC2H2 基因僅含有1 個外顯子。此外,9 個EgC2H2 基因含有內含子,其他均無內含子。
2.5 油棕C2H2 基因家族保守基序分析
利用在線網站MEME 搜索獲得了73 個EgC2H2 基因的10 個保守基序(圖4),其中Motif1、Motif 2 和Motif 6 含有C2H2 型鋅指蛋白保守結構域,都存在典型的QALGGH 基序,屬于Q型C2H2 鋅指蛋白;Motif 1 和Motif 2 含有21 個保守氨基酸,Motif 6 含有26 個保守氨基酸。EgC2H2 基因家族成員含有1 個或多個保守結構域,其中Motif 1 在EgC2H2 基因家族分布最廣,62 個EgC2H2 基因含有Motif 1,說明Motif 1 在EgC2H2 基因家族發揮關鍵作用,7 個EgC2H2 基因同時具有Motif 1、Motif 2 和Motif 6。此外Motif7、Motif 9 和Motif 5 在EgC2H2 基因家族中也分布較多,說明EgC2H2 基因家族保守結構域的種類和數量比較豐富。Motif 10 和Motif 5 為第Ⅰ亞家族所特有,且每個成員均含1 個Motif 5,Motif8 為第Ⅲ亞家族特有,同一亞家族成員的保守結構域具有較高的一致性。
2.6 油棕C2H2 基因家族啟動子順式元件分析
對油棕C2H2 基因家族起始位點上游2000 bp的啟動子序列進行順式元件分析(圖5),結果表明,油棕C2H2 基因家族含有多種啟動子順式元件,除EgC2H2-36 和EgC2H2-06 外,所有C2H2啟動子均含有1 個或多個啟動子順式元件。光響應元件在EgC2H2 基因家族分布最廣,除EgC2H2-36、EgC2H2-41、EgC2H2-21 和EgC2H2-06 外,其他EgC2H2 基因均含有光響應元件,此外,EgC2H2 基因家族還含有大量與植物激素和逆境脅迫有關的響應元件,如83.6%的EgC2H2 家族成員含有厭氧誘導響應元件,75.3%的EgC2H2家族成員含有脫落酸響應元件,47.9%的EgC2H2家族成員含有生長素響應元件,58.9%的EgC2H2家族成員含有赤霉素響應元件,69.9%的EgC2H2家族成員含有茉莉酸甲酯誘導響應元件,54.8%的EgC2H2 家族成員含有干旱響應元件,50.7%的EgC2H2 家族成員含有水楊酸響應元件,42.5%的EgC2H2 家族成員含有低溫響應元件,此外,部分EgC2H2 家族成員還含有防御與脅迫響應元件。表明EgC2H2 家族在響應逆境脅迫中發揮重要作用。
2.7 油棕C2H2 基因家族表達分析
對耐寒材料和低溫敏感材料低溫脅迫0、1、3、5 d 的轉錄組數據進行分析,使用TBtools 軟件繪制油棕C2H2 基因家族表達模式的熱圖(圖6)。結果顯示,低溫脅迫下,EgC2H2 基因在不同材料之間和同一材料不同處理時間之間的表達量有明顯差異,說明EgC2H2 家族基因能夠被低溫誘導。低溫脅迫下,EgC2H2 基因家族中有10個基因未表達,其他基因均有表達,且在CT 的表達量變化較CS 顯著,上調表達基因中大量基因在CT3 和CT5 中表達量相對較高,少量基因在CS3 和CS5 中表達量高,下調表達基因中大量基因在CT0 和CT1 中表達量高,少量基因在CS0和CS1 中表達量高。隨低溫脅迫時間的延長,EgC2H2-11、EgC2H2-07、EgC2H2-23、EgC2H2-17、EgC2H2-21、EgC2H2-33、EgC2H2-67、EgC2H2-16和EgC2H2-28 的表達量在CT 中持續升高,在CT5處理下表達量最高。EgC2H2-25、EgC2H2-26 和EgC2H2-30 的表達量在CS 中持續升高,在CS5處理下表達量最高。隨低溫脅迫時間的延長,EgC2H2-43、EgC2H2-39、EgC2H2-66、EgC2H2-69、EgC2H2-14 和EgC2H2-50 表達量持續降低,在低溫處理5 d 時表達量最低。
2.8 油棕C2H2 基因家族成員在低溫脅迫下qRT-PCR 分析
根據轉錄組數據分析篩選了16 個顯著差異表達的EgC2H2 基因進行熒光定量PCR 分析(圖7)。結果表明,低溫脅迫處理后,16 個EgC2H2基因在同一材料不同處理時間下差異顯著,且變化趨勢與轉錄組數據基本一致,EgC2H2-20、EgC2H2-24、EgC2H2-71、EgC2H2-27 和EgC2H2-19 基因的表達量隨脅迫時間的延長在CT 中呈先升高再降低的趨勢,在低溫處理3 d 時達到最高水平,且表達量明顯高于CS,但在CS中則呈持續升高的趨勢。EgC2H2-07、EgC2H2-26、EgC2H2-11、EgC2H2-73、EgC2H2-17、EgC2H2-16、EgC2H2-30、EgC2H2-21、和EgC2H2-23 基因的表達量隨脅迫時間的延長呈升高的趨勢,其中EgC2H2-26 和EgC2H2-30 基因低溫脅迫后在CS 中的表達量高于CT,EgC2H2-11、EgC2H2-17、EgC2H2-16、EgC2H2-21 和EgC2H2-23 基因低溫脅迫后在CT 中的表達量顯著高于CS,且升高的幅度較大,低溫脅迫5 d 時表達量分別是CS 的2.0、3.7、2.4、5.5 和1.4 倍。EgC2H2-11、EgC2H2-17、EgC2H2-21、EgC2H2-19 和EgC2H2-23 基因低溫脅迫下表達量上升的幅度遠遠高于其他基因,可能與耐寒材料較強的抗寒能力有關。
3 討論
鋅指蛋白是植物中最大的轉錄因子家族之一,C2H2 型鋅指蛋白轉錄因子是目前研究較多的鋅指蛋白,在植物的生長發育以及高鹽、低溫、干旱等非生物脅迫響應中扮演重要作用[23, 34-35]。目前國內外關于C2H2 型鋅指蛋白在其他植物中的報道越來越多,如矮牽牛、擬南芥、水稻、楊樹、小麥、馬鈴薯等[9-12, 14, 17],然而油棕C2H2型鋅指蛋白家族的研究還未見報道。本研究對油棕C2H2 型鋅指蛋白家族的全基因組鑒定,共獲得73 條含有C2H2 鋅指蛋白保守結構域的油棕C2H2 基因,比擬南芥(176 個)[10]、煙草(116個)[13]、水稻(189 個)[11]、小麥(122 個)[14]、楊樹(109 個)[12]、番茄(92)[15]和木薯(85)[16]的C2H2 基因家族成員數目少,說明不同物種間數量差異較大,C2H2 基因在進化過程中可能發生了基因復制。
構建系統進化樹發現,油棕C2H2 基因家族可以分為4 個亞家族,每個亞家族的結構域類型存在高度的一致性,第Ⅲ、Ⅳ亞家族的EgC2H2基因均含有zf-C2H2 結構域,而第Ⅰ、Ⅱ亞家族的EgC2H2 基因均含有典型的zf-C2H2-6 結構域,且第Ⅰ和第Ⅱ亞家族成員數量占總數的85%,說明zf-C2H2-6 結構域是油棕C2H2 家族中發揮作用的重要結構域。保守基序分析發現,每個成員均至少含有1 個鋅指蛋白保守結構域,不同亞家族motif 類型和數量存在明顯差異,但同一亞家族C2H2 成員的保守基序和數量具有較高的一致性,推測同一亞家族基因功能接近或相同。Motif1、Motif 2 和Motif 6 結構域中均含有植物C2H2鋅指蛋白特有的一段高度保守的QALGGH 基序,早期研究報道QALGGH 基序是與DNA 的結合的重要基序,在植物的生長發育中發揮重要作用[7-8]。基因結構分析發現,油棕C2H2 基因家族的基因結構簡單,大部分成員含有1 個外顯子,且數量差異較小,僅極少數成員含有內含子,屬于內含子缺失型,說明油棕C2H2 基因家族在進化過程中可能發生了內含子丟失的現象,這與在大麻、木薯、小麥、煙草等植物中的研究結果類似[13-14, 16, 18]。C2H2 基因家族的外顯子和內含子數目在不同物種間具有較高的保守性,有研究報道,內含子缺失型基因可以更快速地響應逆境脅迫[36]。啟動子順式元件預測分析發現油棕C2H2家族含有大量與植物激素和逆境脅迫有關的響應元件,這些結果與其他植物發現的順式元件結果類似[12, 14, 35],進一步說明油棕C2H2 基因家族參與非生物脅迫過程。
大量研究表明,C2H2 型鋅指蛋白在植物的抗寒中發揮重要作用。過表達C2H2 基因可以增強擬南芥、水稻、煙草等多種植物的抗寒性[19-21, 24-26]。本研究對不同低溫處理時間的轉錄組數據分析顯示,油棕C2H2 基因家族成員中63 個基因在不同處理時間之間的表達量有明顯差異,說明油棕C2H2 基因家族響應低溫脅迫,這些基因大多數在耐寒材料中高表達,而在冷敏感材料中低表達,這可能與耐寒材料較強的抗寒能力有關。qRT-PCR 結果顯示,16 個EgC2H2 基因在低溫脅迫下的表達量均發生了顯著變化,除EgC2H2-26、EgC2H2-09 和EgC2H2-30 外,其他基因的表達量變化趨勢均顯著高于冷敏感材料,推測EgC2H2基因在耐寒材料中感受低溫的速度快于冷敏感材料,耐寒材料可能通過增加EgC2H2 基因的快速表達加快了低溫信號的傳遞,并誘導下游冷響應基因的表達,從而使植株表現出較強的抗寒能力。本研究還發現, EgC2H2-11 、EgC2H2-17 、EgC2H2-21、EgC2H2-19 和EgC2H2-23 顯著上調表達與谷子中SiC2H2-78 的表達模式類似[37],且表達量顯著高于冷敏感材料,表明耐寒材料可能通過提高這些基因的表達量來抵御低溫傷害,預示著這些基因在低溫脅迫中起正向調控作用,可以作為抗寒候選基因進一步深入研究。
