皺木耳漆酶生物信息學分析*
2023-05-28 02:03:56李雪松孫達鋒岳萬松劉春麗袁紹保羅孝坤劉紹雄
中國食用菌 2023年2期
李雪松,孫達鋒,華 蓉,岳萬松,劉春麗,袁紹保,羅孝坤,劉紹雄**
[1.云南云菌(集團)有限公司,云南 昆明 650221;2.陸良爨鄉綠圓菇業有限公司,云南 陸良 655601;3.中華全國供銷合作總社昆明食用菌研究所,云南 昆明 650221]
皺木耳(Auricularia delicata)隸屬于真菌界(Kingdom Fungi)擔子菌門(Basidiomycota)傘菌綱(Agaricomycetes)木耳目(Auriculariales)木耳科(Auriculariaceae)木耳屬(Auricularia),在我國是一種傳統的食(藥)用菌,具有抗氧化、抗病毒、保肝解毒等功效,常用于治療各種胃腸道和肝臟疾病[1]。目前,黑木耳(Auricularia heimuer)、毛木耳(Auricularia cornea)等木耳品種已被廣泛栽培及推廣,但木耳屬其他物種的資源開發利用相對較少。與黑木耳、毛木耳相比,皺木耳耳片質地較厚,子實層具有明顯的網狀褶皺,口感更加松脆爽口[2]。皺木耳的野生資源豐富,主要分布于我國的西南地區,特別是云貴地區。目前,對于皺木耳生物學特性以及生長發育調控基因相關的基礎研究較少,這成為了限制皺木耳栽培及推廣的因素之一。
漆酶是一種含銅的多酚氧化酶,與抗壞血酸氧化酶、哺乳動物血漿銅藍蛋白、膽紅素氧化酶等同屬于藍色多銅氧化酶家族MCO(multi copper oxidase,MCO)[3]。目前,在細菌、真菌、植物及動物中均有關于漆酶的研究報道。其中,真菌漆酶研究最多,已研究的物種包括雜色云芝(Coriolus versicolor)、糙皮側耳(Pleurotus ostreatus)、變色栓菌(Trametes versicolor)、灰蓋鬼傘(Coprinopsis cinerea)等。真菌漆酶主要參與其形態建成、病原菌致病、色素合成、應激反應等多種生物學過程,對其生長發育十分重要[4-5]。同時,由于漆酶具有廣泛的底物作用性,可作為綠色生物催化劑應用于工業、農業、食品、環保、醫藥等多個領域,具有廣闊的市場前景[6]。現有的研究表明,真菌漆酶基因一般包含多個拷貝且通常成簇排列,每一個拷貝的漆酶同源基因可能行使著某一特定的功能,而特定的功能往往與其特有的結構直接相關[7]。因此,對皺木耳漆酶基因的結構展開分析,將有助于了解其生物學功能及作用機制。
本研究對皺木耳的漆酶進行了生物信息學分析,分析了漆酶基因結構、蛋白結構及理化性質。這些結果進一步明確了皺木耳漆酶基因的差異及相關的功能,對闡明皺木耳的生長發育過程、漆酶的開發利用等具有重要意義。同時,通過對皺木耳生長發育過程進行研究,也為規范栽培、優良菌株選育等提供基礎支撐。
1 材料與方法
1.1 材料
從NCBI 數據庫中下載皺木耳的基因組序列、基因注釋文件、蛋白序列文件,灰蓋鬼傘、雙色蠟蘑(Laccaria bicolor)、草菇(Volvariella volvacea)等的漆酶蛋白序列。皺木耳、雙孢蘑菇(Agaricus bisporus)、黑木耳、變色栓菌等的漆酶參考序列從UniProt 數據庫獲取。
1.2 皺木耳漆酶序列的鑒定
首先,使用HMMER 軟件構建漆酶的保守結構域模型,E 值設置為e<10-10,比對皺木耳的所有蛋白序列,作為HMM 部分漆酶的結果。其次,為保證候選漆酶基因的準確性和完整性,以參考漆酶序列為目標序列,利用BLAST 軟件從皺木耳的蛋白文件中搜索漆酶,E 值設置為e<10-4,篩選出的序列為BLAST 部分漆酶的結果。然后,將HMM 和BLAST的結果合并,并將合并后的結果通過BLASTP 軟件與UniProtKB 數據庫比對,E 值小于10-10的被認為是候選漆酶。最后,再利用Pfam 數據庫和CDD 數據庫預測這些候選蛋白的結構域,若存在結構域,則認為該候選蛋白屬于漆酶;若未能檢測出結構域,則認為其不屬于漆酶。
1.3 皺木耳漆酶蛋白的理化性質分析
利用ProtParam 工具預測漆酶蛋白的相關理化性質,指標包括氨基酸數目、分子量、理論等電點、蛋白質疏水性等。漆酶蛋白的亞細胞定位分析使用PSLDoc[8]工具完成。
1.4 皺木耳漆酶基因結構和蛋白結構分析
通過皺木耳的基因組注釋文件獲取漆酶基因的結構信息,使用Tbtools[9]構建基因結構圖。利用SOPMA[10]預測皺木耳漆酶蛋白的二級結構。同時,使用SWISS-MODEL 對漆酶的氨基酸序列進行同源建模,生成蛋白質三級結構。基于MEME[11]工具對皺木耳漆酶蛋白的模體序列進行分析,其中motifs 的最大數量設置為10,其他參數采用默認值。使用Pfam 數據庫和CDD 數據庫開展結構域預測,取2 種預測結果的并集作為最終漆酶蛋白的結構域結果。
1.5 皺木耳漆酶蛋白的系統發育分析
從NCBI 數據庫和UniProt 數據庫中下載雙孢蘑菇、黑木耳、毛木耳、灰蓋鬼傘、雙色蠟蘑、射脈菌(Phlebia radiata)、糙皮側耳、鳳尾菇(Pleurotus sajorcaju)、毛栓菌(Trametes hirsuta)、變色栓菌、長絨毛栓菌(Trametes villosa)、草菇的漆酶蛋白序列。使用MUSCLE 軟件對所有的漆酶序列進行比對,然后使用MEGA[12]構建系統進化樹,自展值設置為1 000。
2 結果與分析
2.1 皺木耳漆酶序列的鑒定結果
通過HMMER 程序構建模型,利用該模型從皺木耳基因組中檢索到13 個蛋白序列含有2 個及以上多銅氧化酶的結構域。通過BLAST 程序比對UniProtKB 數據庫,發現有11 個可能是皺木耳漆酶的蛋白序列。最后,合并HMMER 和BLAST 的結果,再通過篩選序列長度、3 個多銅氧化酶結構域等特征,獲得8 個皺木耳漆酶蛋白,詳見表1。
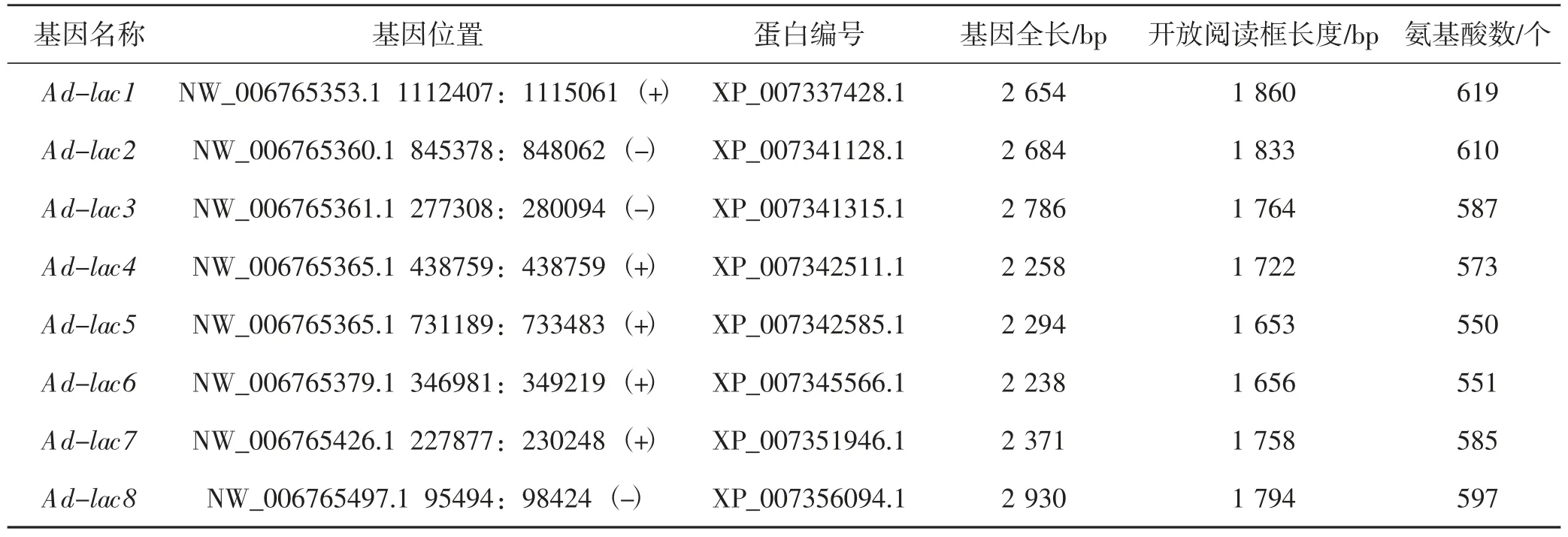
表1 皺木耳漆酶基因信息分析Tab.1 Information of laccase gene in Auricularia delicata
如表1世紀,皺木耳的8 個漆酶基因長度為2.2~2.9 Kb,編碼蛋白的長度也較為相近,最長為619 個氨基酸,最短為550 個氨基酸;除Ad-lac3與Ad-lac4位于同一段scaffold 上外,其余的漆酶基因都位于不同的scaffold 上。
2.2 皺木耳漆酶蛋白的理化性質分析結果
對皺木耳漆酶蛋白的理化性質進行預測,分析結果詳見表2。
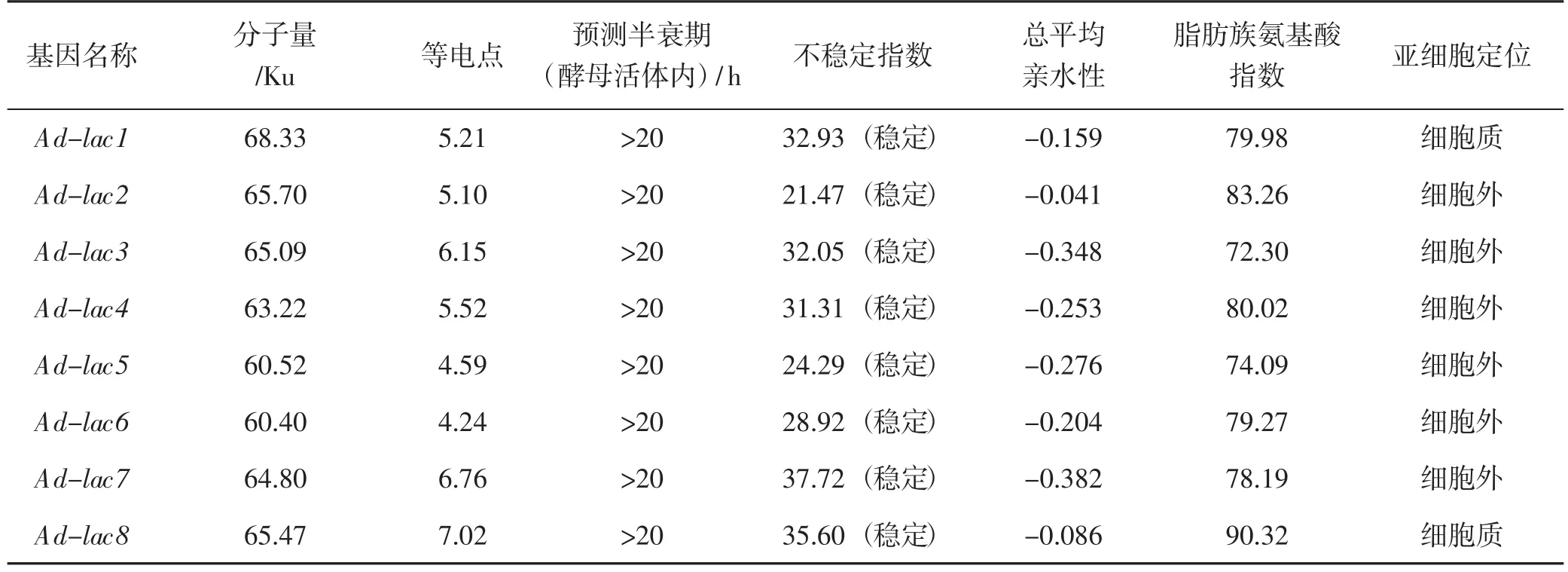
表2 皺木耳漆酶基因氨基酸的理化分析Tab.2 The physico-chemical analysis of amino acid in Auricularia delicata
由表2 可知,這些基因所表達的蛋白分子量為60.40~68.33 Ku,等電點為4.24~7.02,大多數在堿性范圍,說明蛋白分子中富含酸性氨基酸。半衰期(酵母活體內)是判斷蛋白在細胞內是否穩定的指標之一,大于20 h 的為穩定蛋白;預測結果表明皺木耳的漆酶蛋白均較穩定。總平均親水性GRAVY(grand average of hydropathy,GRAVY)指所有氨基酸親水值的總和與氨基酸個數之間的比值,負值代表親水性,負值越大表明親水性越強,而正值代表疏水性,正值越大表明疏水性越強[13]。皺木耳漆酶蛋白序列間的GRAVY 值雖存在差異,但均為親水性蛋白。亞細胞定位的分析結果表明,有6 個皺木耳的漆酶蛋白均分泌到細胞外,可能是作為分泌蛋白在細胞外分解酶;而Ad-lac1與Ad-lac8定位于細胞質中,說明皺木耳漆酶基因的生理功能存在差異。
2.3 皺木耳漆酶基因結構和蛋白結構分析結果
利用本研究中皺木耳漆酶的蛋白序列構建8 個漆酶的系統發育樹,結果詳見圖1。

圖1 皺木耳漆酶基因的系統進化樹和基因結構Fig.1 Phylogenetic tree and gene structures of Auricularia delicata laccase
由圖1 可知,皺木耳的漆酶蛋白基因均含有10個以上的CDS 編碼區域。系統發育關系較近的皺木耳漆酶蛋白的基因結構是相似的,如Ad-lac4和Ad-lac5的CDS 數目和排列幾乎一致,這也從側面體現了漆酶基因具有一定的保守性。
蛋白質二級結構是指不同氨基酸間C=O 和N-H基團間氫鍵形成的穩定結構,主要為α-螺旋和β-折疊。利用SOPMA 軟件,預測皺木耳8 條漆酶基因編碼蛋白質的二級結構,結果詳見表3。
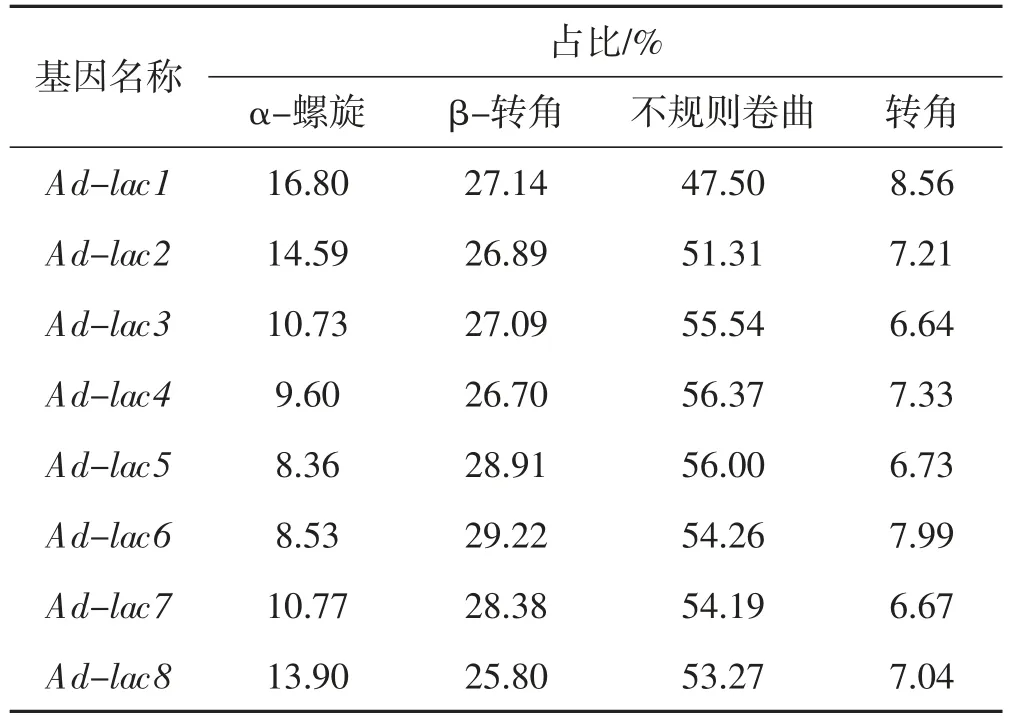
表3 皺木耳漆酶蛋白的二級結構Tab.3 The secondary structure of protein sequence in Auricularia delicata
由表3 可知,8 條序列的結構類型主要是不規則卷曲。其中,α-螺旋(Alpha helix)占8.36%~14.59%、β-轉角(Beta turn)占25.80%~29.22%、不規則卷曲(Random coil)占47.50%~56.37%,不規則卷曲所占比例遠高于其他的類型。
模體(motif)是蛋白質分子結構中介于二級結構與三級結構之間的結構層次,又稱超二級結構,是蛋白質分子具有特定功能或作為獨立結構域一部分的二級結構聚合體[14]。基因所有或者大多數成員共有的motif 極可能是該家族執行重要功能或組成結構不可缺少的部分[15]。所以,識別基因家族共同的motif 就能刻畫該基因家族的特征,從而可以利用這些特征來發掘基因家族新成員[14]。本研究中皺木耳漆酶蛋白的基序情況詳見圖2。
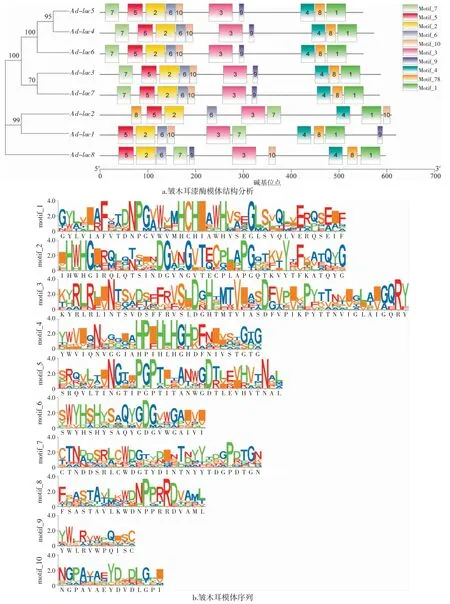
圖2 皺木耳漆酶基因的模體Fig.2 Motif of the Auricularia delicata laccase genes
如圖2世紀,皺木耳的8 個漆酶蛋白均含有10個motif。其中,motif_1 中的HCHxxxH、motif_2 中的HWH、motif_4 中的HPxHLH、motif_6 中的HSH與報道的真菌漆酶的特征序列和草菇的漆酶蛋白序列具有一致性[16]。
在較大的蛋白質分子中,由于多肽鏈上相鄰的超二級結構緊密聯系,進一步折疊形成1 個或多個相對獨立的致密三維實體,即結構域(domain)[17]。一般每個結構域由100~200 個氨基酸殘基組成,各有不同的空間構象,并具有不同的生物學功能。根據已有的研究報道,真菌漆酶一般都含有3 個多銅離子氧化酶結構域,排列為Cu-oxidase_3、Cu-oxidase 和Cu-oxidase_2。在多銅氧化酶的研究中發現了3 種光譜上不同的類型:1 型(藍色)、2型(正常)和3 型(耦合雙核),利用銅離子特有的氧化還原能力,對還原性底物進行單電子氧化,同時傳遞4 個電子,將作為第二底物的氧氣還原成水[18]。本研究中得到的8 個皺木耳的漆酶蛋白序列的情況詳見圖3。
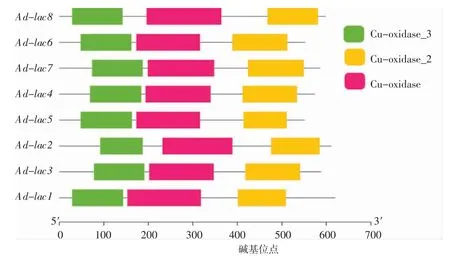
圖3 皺木耳漆酶基因的結構域Fig.3 Domain of the Auricularia delicata laccase genes
如圖3世紀,皺木耳的漆酶蛋白序列也同樣有這樣的結構域和相應的排列順序,進一步說明篩選出的皺木耳漆酶也應具有相同的生物學功能。
通過Swiss-Model 對皺木耳的漆酶蛋白序列進行三維結構同源建模,結果詳見圖4。
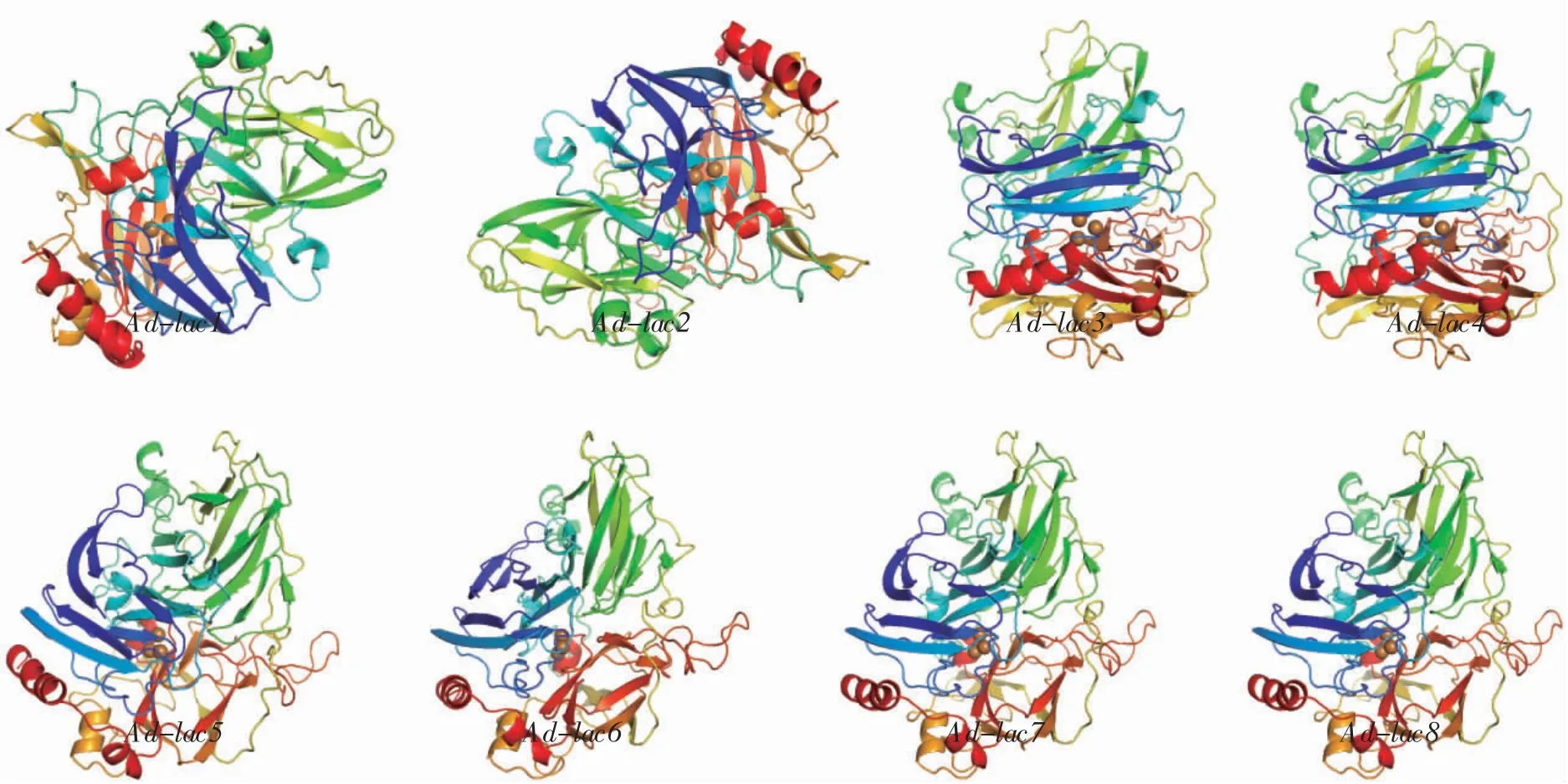
圖4 皺木耳漆酶蛋白三維結構Fig.4 Protein 3D structure of Auricularia delicata laccase
如圖4世紀,8 個漆酶蛋白大部分都具有十分相似的三維結構。每個蛋白由3 個杯狀(Cu-oxidase)結構域(Domain 1、Domain 2、Domain 3)組成,每個結構域均有1 個β 桶狀結構。8 個皺木耳漆酶蛋白的三維結構由多個α-螺旋和β-折疊構成,都具有對應的銅離子結合位點和催化的活性中心。
2.4 皺木耳漆酶蛋白的系統發育分析結果
通過合并8 條皺木耳的漆酶蛋白序列和下載的69 條蛋白序列,總計77 條漆酶序列,構建系統發育樹,結果詳見圖5。
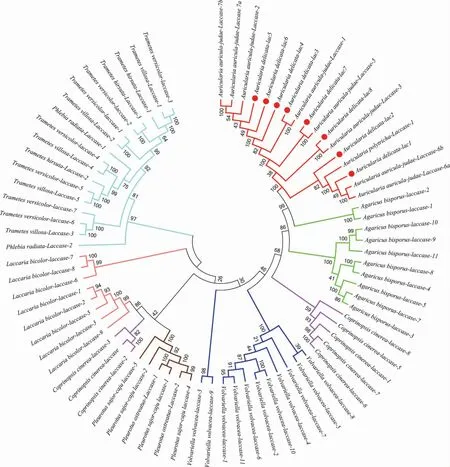
圖5 皺木耳及其他物種漆酶蛋白序列的系統發育樹Fig.5 Laccase protein sequences phylogenetic tree of Auricularia delicata and other species
如圖5世紀,77 條漆酶主要分為7 個類群,主要的大類群是木耳屬與栓菌屬(Trametes),其他的各個類群的聚類關系與其物種相對應。本研究中篩選出的皺木耳漆酶與木耳屬的黑木耳、毛木耳的漆酶聚為一枝(枝長顏色為紅色),表明皺木耳的漆酶與木耳屬其他物種漆酶的基因之間具有較高的保守性。其中,Ad-lac3、Ad-lac7和Ad-lac8各在1 個分枝上;Ad-lac1和Ad-lac2聚為一枝;Ad-lac4、Ad-lac5和Ad-lac6聚為一枝。結合皺木耳漆酶的基因結構和蛋白結構的分析結果,發現親緣關系較近的漆酶蛋白的基因結構具有一致性。
3 結論與討論
針對皺木耳的漆酶進行了生物信息學分析,結果表明在皺木耳基因組中有8 個漆酶基因,分布在基因組的不同位置,這與目前報道的擔子菌和子囊菌中漆酶基因通常成簇排列是不一致的[19-21]。原因可能是皺木耳的基因組測序時間較早,測序組裝技術具有局限性,導致基因組碎片化程度較高及注釋不完全,最終使篩選結果不全面。進一步分析了皺木耳漆酶蛋白氨基酸的定位與理化性質,發現8 個皺木耳的漆酶蛋白理化性質總體差異較小,但Adlac1與Ad-lac8基因所表達的蛋白不是細胞外的分泌蛋白,2 個漆酶蛋白可能是在細胞內行使功能。在皺木耳的基因結構分析結果中也可以看出,皺木耳不同漆酶的基因結構不一致。漆酶在真菌的形態建成、色素合成、應激反應等生長發育過程中起重要的作用,不同物種的漆酶也應該具有一定的保守性。皺木耳的8 個漆酶蛋白的二級結構以不規則卷曲為主,這與黑木耳漆酶蛋白情況相似。同時,對皺木耳8 個漆酶模體的分析發現,HCHxxxH、HWH、HPxHLH、HSH 等序列的特征與草菇等的漆酶蛋白序列具有一致性;結構域分析發現,多銅離子氧化酶結構域的排列特征與毛栓菌、變色栓菌等的研究結果一致[16]。8 條漆酶蛋白序列的三維結構特征與二級結構、模體、結構域的結果一致。這些特征都表明篩選出的8 個漆酶蛋白具有生物學功能,在皺木耳的生長發育中扮演著重要的角色。而系統發育樹的結果則說明,同一類群物種的漆酶具有較近的親緣關系,可能受相似的選擇壓力作用,具有相似的生物學功能,以便適應相應的生存環境。
目前皺木耳的研究仍聚集在生物學特性、馴化及栽培上,本研究中首次較全面地從皺木耳基因組中篩選出漆酶序列,對皺木耳漆酶基因結構、蛋白質結構等進行了分析,闡釋了皺木耳漆酶基因的特性和各基因之間的差異。同時,了解了皺木耳漆酶不同成員之間的相互關系。而對于皺木耳的每一個漆酶的生物學功能,還需要進行基因克隆以及表達分析來驗證。研究結果為闡明皺木耳的生長發育過程、漆酶的開發利用等提供了理論支持和試驗依據。
猜你喜歡
哲學評論(2021年2期)2021-08-22 01:53:34
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
現代企業(2015年9期)2015-02-28 18:56:50
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
終身教育研究(2014年5期)2014-02-28 01:23:06
