寧夏賀蘭山東麓葡萄灰霉病菌類群分析
2023-05-25 08:20:44李曉慶閆思遠顧沛雯
中外葡萄與葡萄酒 2023年3期
關鍵詞:生長
李曉慶,閆思遠,顧沛雯
(寧夏大學農學院,寧夏銀川 750021)
寧夏賀蘭山東麓地區是我國最大的葡萄酒地理標志保護產區,隨著葡萄種植面積不斷擴大,由葡萄灰葡萄孢(Botrytis cinerea)引起的葡萄灰霉病已成為寧夏賀蘭山東麓葡萄產業重大病害之一[1]。據調查,不同年份造成的葡萄產量損失在20%~50%[2]。灰葡萄孢可以侵染葡萄的葉片、花穗、果實和穗軸等,環境適宜時會在病斑表面產生鼠灰色霉層,直接影響葡萄的產量和品質[3-4]。
葡萄灰葡萄孢屬于復合種,由于繁殖快、遺傳變異大和適合度高的特點[5],導致其產生很多類群。不同類群菌株之間致病性和抗藥性均存在較大差異,這給葡萄灰霉病的有效防治帶來了巨大的挑戰,因此對灰葡萄孢進行類群劃分十分重要。蔣忠洪[6]根據生長特征將來自湖南不同地區的223株灰霉病菌株劃分為菌絲型、菌核型、孢子型3種類型。張艷杰等[3]對來自全國不同地區的143個灰葡萄孢菌株進行形態學劃分,其中40.56%為菌絲型、59.44%為菌核型,根據其生長速率可分為高、中、低3個等級,形態學分布與采集地地理距離無明顯相關性。馮曉菲[7]通過菌落類型、生長速率和離體草莓葉片致病力對來自四川195株草莓灰霉病菌株進行分析,表明不同菌株之間存在差異。馮寶珍[8]從運城保護地茄子病株中分離純化出15個灰葡萄孢菌株,依據菌株的生長速率、菌核的大小、形狀,將其劃分為4個類群,分別為菌絲型、孢子型、細小菌核型和大菌核型。
近年來,隨著寧夏賀蘭山東麓產區葡萄種植面積擴大,葡萄灰霉病的危害也越來越嚴重,但賀蘭山東麓灰霉病菌的類群劃分研究較少,對灰霉病類群進行系統劃分顯得尤為重要。本研究以從寧夏賀蘭山東麓地區不同葡萄品種病果中分離的13株葡萄灰霉病菌為研究對象,對比其分別在馬鈴薯葡萄糖瓊脂培養基(PDA)、改良馬丁瓊脂培養基和燕麥培養基上的形態和生長速率、產孢量、微菌核以及致病性差異,并通過分子生物學的方法,對其進行類群劃分,以期摸清該地區灰霉病菌的種群結構,為寧夏賀蘭山東麓葡萄灰霉病的有效防治提供理論依據。
1 材料與方法
1.1 供試材料
葡萄灰霉病病果采集:2021年6—9月,在寧夏賀蘭山東麓葡萄6個酒莊共采集葡萄灰霉病果30份,用自封袋密封后立即帶回實驗室,采用組織分離法分離病原菌[9],共獲得13株葡萄灰霉病菌(B. cinerea)見表1,使用PDA培養基和凍干濾紙進行短期和長期保存。
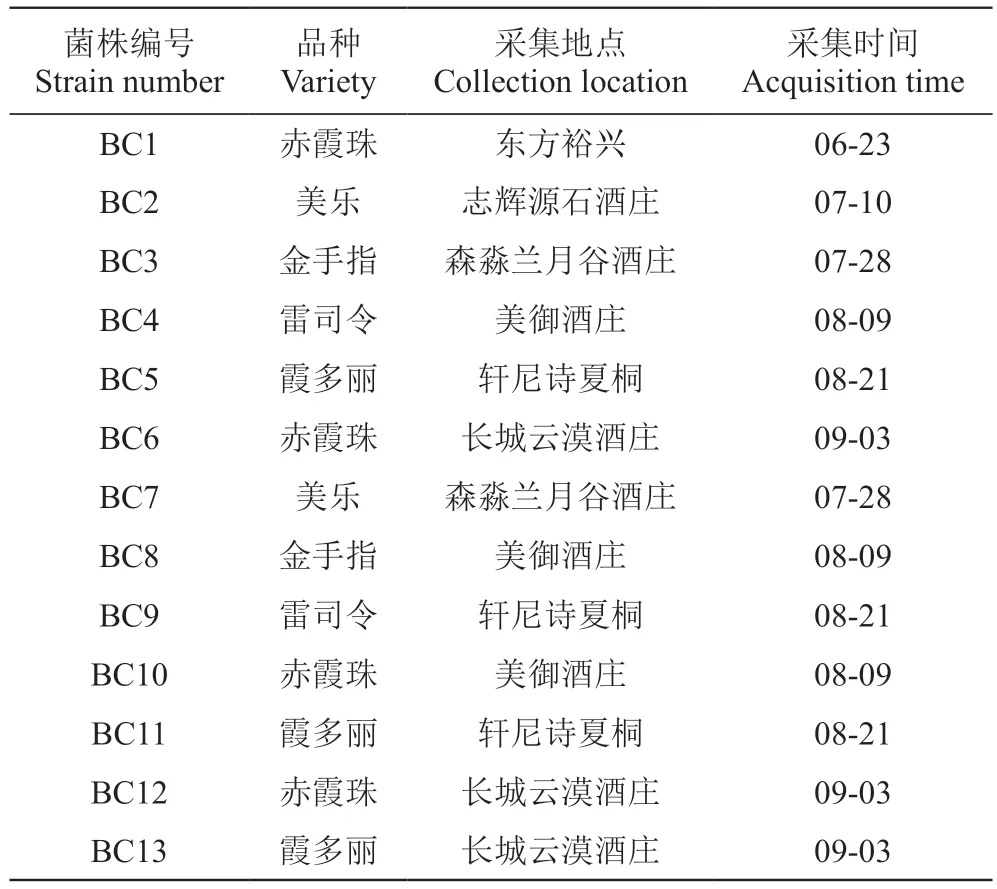
表1 13株葡萄灰霉病菌的采集信息Table 1 Collection information of 13 strains of B. cinerea
2021年8月購買無病斑、果粒大小均等、著色一致的‘紅地球’葡萄果穗,用于致病性測定。
依據13株葡萄灰霉病菌的形態學分析和致病性測定結果,使用SPSS 25進行聚類分析,以歐式距離為10作為聚類分割點,對其進行類群劃分。依據在PDA培養基上的菌落表現及生長速率[14](圖3A)可分為2類,其中菌株BC3、BC6、BC7、BC8、BC9、BC10、BC11和BC12為第一大類,菌株BC1、BC2、BC4、BC5和BC13為第二大類。依據在燕麥培養基上的菌落表現及生長速率(圖3B),可分為3類,其中菌株BC2、BC3、BC5、BC11和BC12為第1類,菌株BC6、BC7、BC8、BC9和BC10為第2類,菌株BC1、BC4和BC13為第3類。依據在改良馬丁培養基上的菌落表現及生長速率(圖3C),可分為3類,其中菌株BC1、BC2、BC3、BC5、BC6、BC8、BC9、BC10、BC11、BC12和BC13為第1類,菌株BC4為第2類,菌株BC7為第3類。依據13株葡萄灰霉病菌產孢量(圖3D),可分為2類,其中菌株BC1、BC2、BC3、BC4、BC5、BC6、BC7、BC8、BC9、BC10、BC11和BC13為第1類,菌株BC12為第2類。依據13株葡萄灰霉病菌產菌核情況(如圖3E),可分為2類,其中菌株BC1、BC2、BC3、BC4、BC5、BC6、BC7、BC9、BC10、BC11、BC12和BC13為第1類,菌株BC8為第2類。依據13株葡萄灰霉病菌致病性(圖3F),可劃分為2類,其中菌株BC1、BC2、BC3、BC4、BC5、BC8和BC13為第1類,菌株BC6、BC7、BC9、BC10、BC11和BC12為第2類。
1.2 供試儀器、試劑、培養基
1.3.3 病菌菌核形體比較
光學顯微鏡(OLYMPUS BX53,Japan);凝膠成像系統(Azure Biosystems c200,USA);Sigma 3K 15通用臺式冷凍離心機(Sigma公司,USA);PCR儀(T100TMThermal Cycler,Singapore)。Biospin?真菌DNA提取試劑盒(BioFlux公司,日本);2×Easy Taq PCR Super Mix、RNase A、DNA marker、Agarose和GelStain染液購自北京全式金生物技術有限公司;50×TAE緩沖液、DNase/RNase-Free Water購自北京索萊寶科技有限公司。
上述區域內的流速場會對物體漂移速度產生影響,因此首先將該區域內所有流場數據提取出來,假設該區域流速場用以下兩個矩陣表示為
1.2.2 供試培養基
馬鈴薯葡萄糖(PDB)培養基:馬鈴薯200 g,葡萄糖20.0 g,定容至1.0 L。馬鈴薯葡萄糖瓊脂(PDA)培養基:馬鈴薯200 g,葡萄糖20.0 g,瓊脂粉15.0 g,定容至1.0 L。改良馬丁瓊脂(GM)培養基:蛋白胨5.0 g,磷酸氫二鉀1.0 g,硫酸鎂0.5 g,酵母浸出粉2.0 g,葡萄糖20.0 g,瓊脂粉15.0 g,定容至1.0 L。燕麥瓊脂(YM)培養基:燕麥片30.0 g制備浸出液,瓊脂粉15.0 g,定容至1.0 L。以上各培養基121℃,20 min高壓蒸汽滅菌后,備用。
丹江口大壩加高工程溢流堰面加高疊梁封堵施工技術……………………………………………… 周利明,丁新中(2.8)
1.3 形態學分析
1.3.1 不同培養基上的培養性狀比較
接種前5 d在PDA培養基上活化培養,使用滅菌打孔器在活化后的菌落邊緣打取直徑為5 mm的葡萄灰霉病菌餅,分別接于PDA培養基、YM培養基和GM培養基中,25 ℃黑暗培養5 d,測定菌落直徑,并記錄其菌落形態,各處理重復3次。
1.3.2 產孢量測定
接種前5 d在PDA培養基上活化培養,使用滅菌打孔器在活化后的菌落邊緣打取直徑為5 mm的葡萄灰霉病菌餅接種于PDB培養基中,180 r·min-1,26±2 ℃下振蕩培養5 d過濾制成孢子懸浮液,血球計數板計數,各處理重復3次。
1.2.1 主要儀器和試劑
回到家里,紫云拿出那個黑色公文包,里面有一張調令文書。所有的手續都辦好了,新的工作單位是華夏出版社。她把房子賣了,帶著兒子,到上海去了。
水平三角形的第(4)浪之后,進入一氣呵成的第(5)浪,至2015年6月7559點結束。當初將2013年6月底的低點劃分作第(4)浪終點,將(4)浪D看作(5)浪1——這是常見的錯誤——因此才有后來的(5)浪5預期。7559點結束第(5)浪,這是循環浪III的頂點,因此后面的調整屬于循環浪IV。其中,第一個浪A可能會于2019年結束,特別是當國證A指跌至上升通道的下軌時,配合波浪理論,將成為循環浪IV第一個買點。
1.4 葡萄灰霉病菌致病性測定
將‘紅地球’果粒用自來水沖洗表面灰塵后,放入75%(Vol)乙醇浸泡3 min消除氣泡,用5%(Vol)次氯酸鈉溶液浸泡5 min,無菌水沖洗4次,置于無菌濾紙晾干果粒表面的水分備用。用無菌接種針刺傷葡萄果粒表面,挑取直徑5 mm的灰霉菌餅接于葡萄果粒傷口處,置于25 ℃、濕度85%的保濕培養箱培養,每菌株處理21粒,無菌水為對照,分別于第5天、第7天觀察并統計發病情況,并計算發病率及病情指數。
病害分級標準[10]如下:
使用Biospin?真菌DNA提取試劑盒提取真菌DNA。PCR反應體系(25 μL):2×Eco Taq?PCR Super Mix 12.5 μL,正、反向引物(10 μmol·L-1)各1 μL(表2),DNA模板1 μL,ddH2O補足。PCR反應條件:94 ℃預變性3 min;94 ℃變性30 s,58~64 ℃退火40 s,72 ℃延伸1 min,35個循環;72 ℃延伸10 min,4 ℃保存。PCR產物經1.5%瓊脂糖凝膠電泳檢測后,凝膠成像系統分析結果。將所得PCR原液送至生工生物工程(上海)有限公司進行測序,并將測序結果在NCBI進行BLAST比對。依據測序結果合并序列相同的菌株,確定菌株的分類地位。
由表5可知,13株葡萄灰霉病菌單孢菌株在接種后第5天發病率均達90.00%以上。其中菌株BC9致病力顯著高于其它菌株,在接種后第5天,發病率為99.04%,病情指數為88.24,產生大量的菌絲將離體果粒迅速包裹,造成果粒腐爛;其次是菌株BC7,在接種第5天時,發病率為99.01%,病情指數為68.63,癥狀表現為產生了少量灰黑色菌絲附著在離體果粒表面,造成果粒部分干枯和果皮凹陷;菌株BC3致病力較弱,在接種后第5天時,發病率為93.07%,病情指數為9.80,發病癥狀為只產生少量菌絲附著于果粒表面,沒有造成任何果粒損傷。
開孔鋼板連接件(perfobond strip connector,也稱為PBL連接件)是1987年,德國學者Leonhardt在解決委內瑞拉的卡羅尼第三大橋組合結構剪力連接件的疲勞問題時提出的一種新型剪力連接件[2].PBL連接件是通過開孔鋼板在混凝土板中形成的一系列混凝土榫、開孔鋼板和貫穿鋼筋共同承擔剪力.
1.5 葡萄灰霉病菌產孢類群的系統發育分析
0級:果實無病;1級:病果(病斑)面積占整個果穗(果粒)面積的5%以下;3級:病果(病斑)面積占整個果穗(果粒)面積6%~15%;5級:病果(病斑)面積占整個果穗(果粒)面積16%~25%;7級:病果(病斑)面積占整個果穗(果粒)面積26%~50%;9級:病果(病斑)面積占整個果穗(果粒)面積50%以上。
1.2.3 觀察指標:術前及術后DASH評分、住院天數、肌電圖、術后當日疼痛VAS評分。DASH評分是一種問卷式的評分方式,旨在調查上肢的癥狀及從事日常活動的能力[17]。VAS疼痛評分法[18](0分~10分):0分:無痛;3分以下:有輕微的疼痛,能忍受;4分~6分:患者疼痛并影響睡眠,尚能忍受;7分~10分:患者有漸強烈的疼痛,疼痛難忍,影響食欲,影響睡眠[19]。剪切波彈性成像,通過發射高頻超聲來激發組織產生剪切波,根據其波速結合彈性測量公式就可以計算出正中神經的實際彈性值從而反映組織的受壓情況[20-21]。

表2 所用引物Table 2 Primers used
1.6 數據分析
數據采用Microsoft Excel 2019對灰霉病樣進行統計分析,使用IBM SPSS Statistics 25進行單因素方差分析及聚類分析。使用MEGA 6.0軟件采用鄰接(Neighbor-Joining)法構建系統發育樹。應用SDTv.1.2軟件分析基因序列間核酸的一致性。
2 結果與分析
2.1 灰葡萄孢在不同培養基上的形態差異
圖1、表3分別為13株葡萄灰霉病菌株在3種培養基上5 d時的菌落形態和平均生長速率。在PDA培養基上,菌株BC1、BC2、BC4、BC5和BC13菌絲生長速率較快,均大于0.53 cm·d-1,菌絲濃密;菌株BC6、BC7、BC8、BC9和BC12菌絲生長速率其次,平均速率為0.36~0.39 cm·d-1;而菌株BC3、BC10和BC11的菌絲生長速率較慢,平均生長速率為0.30~0.33 cm·d-1,菌株BC9的生長速率顯著高于菌株BC3、BC10,菌絲稀疏。在燕麥培養基上,菌株BC7、BC9和BC10菌絲生長速率較快,平均生長速率均大于0.49 cm·d-1,菌絲濃密;菌株BC3、BC5、BC6、BC8、BC11和BC12生長速率較慢,平均生長速率為0.35~0.45 cm·d-1。在改良馬丁培養基上,菌株BC3、BC4、BC5和BC10菌絲生長速率較快,平均生長速率均大于0.6 cm·d-1,菌絲濃密;菌株BC1、BC2、BC9和BC12生長速率較慢,平均生長速率為0.52~0.56 cm·d-1。

圖1 13株葡萄灰霉病菌菌株在不同培養基上的菌落形態Figure 1 Colony morphology of 13 Botrytis cinerea strains on different media
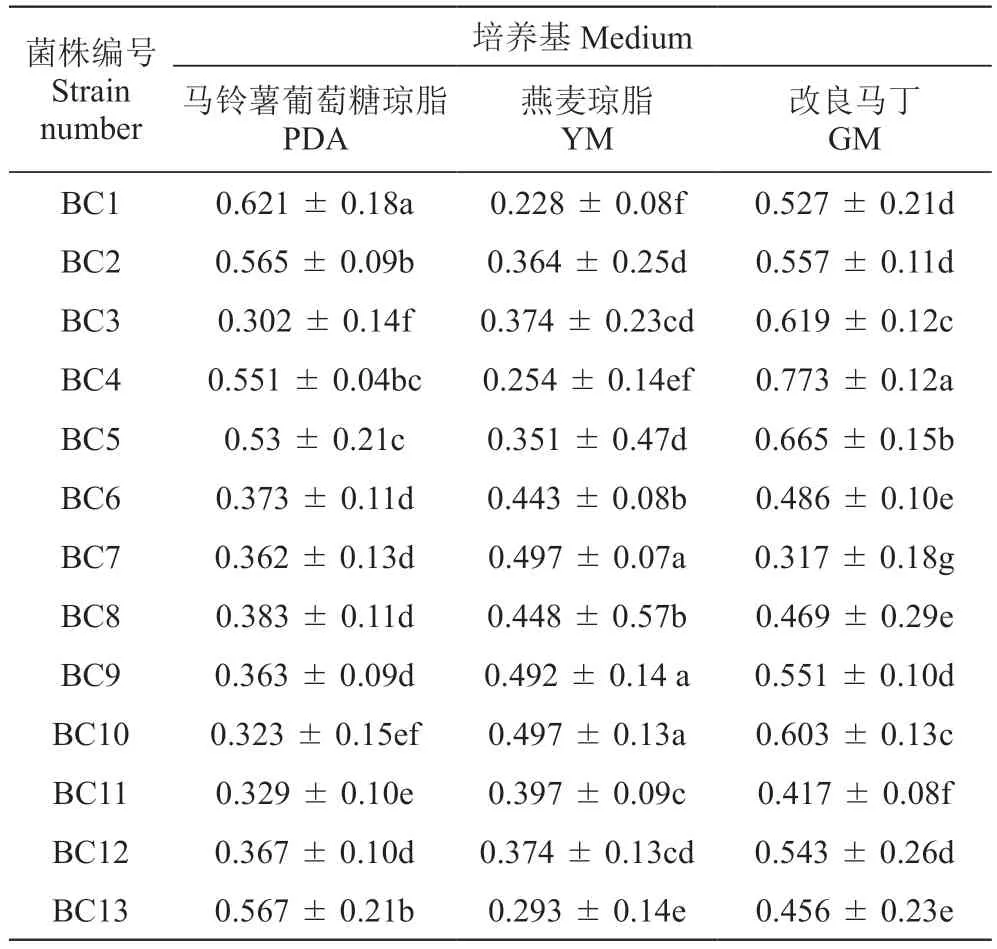
表3 13株葡萄灰霉病菌生長速率Table 3 Growth rate of 13 Botrytis cinerea strains cm·d-1
2.2 葡萄灰霉病的產孢量差異
在表4中,除BC13外,其他12株菌株均能產生分生孢子,其分生孢子形狀為卵圓或橢圓形。其中菌株BC12的產孢量最多,為6.32×107cfu·mL-1。
打取直徑為5 mm的葡萄灰霉病菌餅接種于PDA培養基中,25 ℃黑暗培養30 d后測定菌核數量和大小,各處理重復3次。
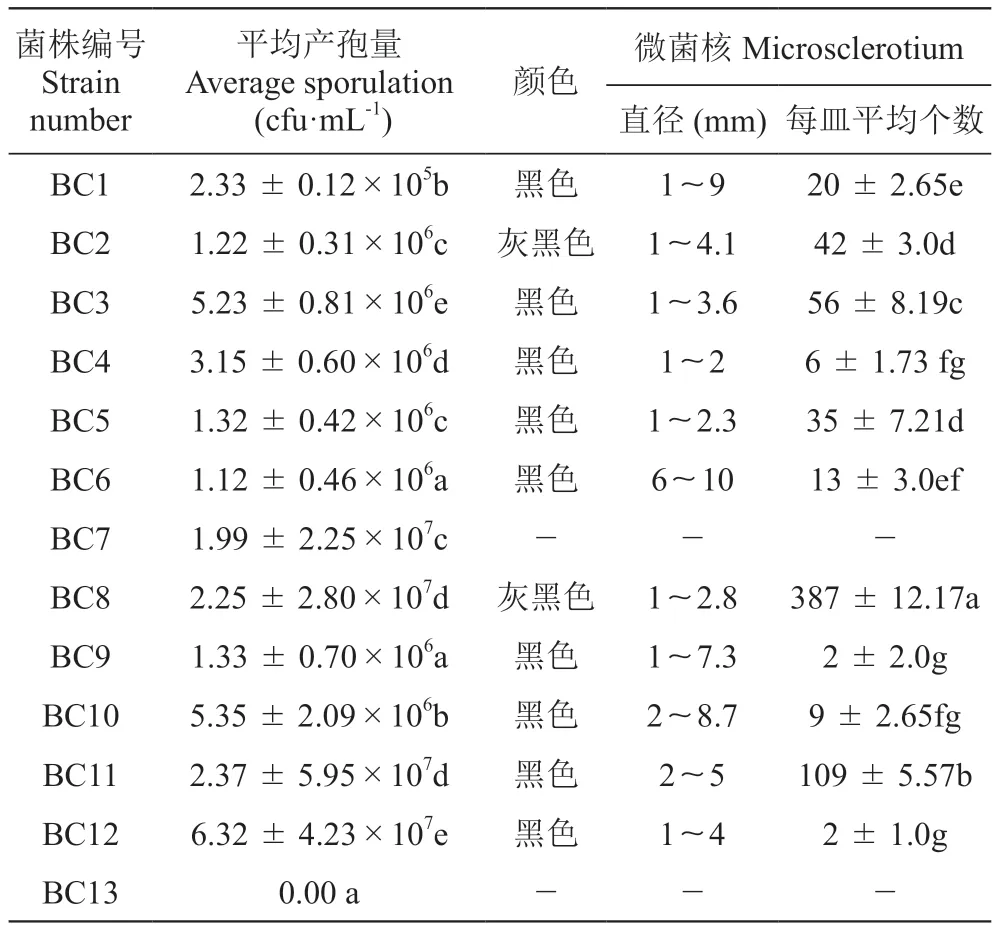
表4 13株葡萄灰霉病菌的產孢量和微菌核Table 4 Spore production of 13 Botrytis cinerea strains
2.3 葡萄灰霉病菌微菌核形態差異
由表4和圖2可知,13株葡萄灰霉病菌單孢菌株中,只有菌株BC7和BC13不產生菌核,其余11株都可產生微菌核。菌株BC8產生的菌核數最多,顏色為灰黑色、每皿菌核數量387個,直徑為1~2.8 mm。其次是菌株BC11產生的菌核顏色為黑色、每皿菌核數量109個,直徑為2~5 mm。菌株BC9和菌株BC12產生的菌核數較少,顏色均為黑色,菌株BC9每皿菌核數量2個,直徑為1~1.7 mm,菌株BC12每皿菌核數量2個,大小為1~4 mm。
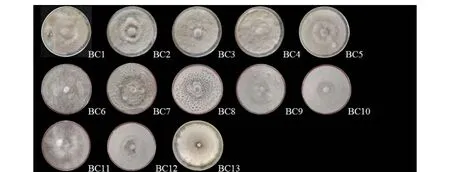
圖2 13種葡萄灰霉病菌微菌核產生情況Figure 2 Microsclerotia production of 13 species of Botrytis cinerea
2.4 不同葡萄灰霉病菌的致病力差異
病情指數=(∑各級病果穗或果粒數×病害級別數)/(調查總果穗或果粒數×9)×100
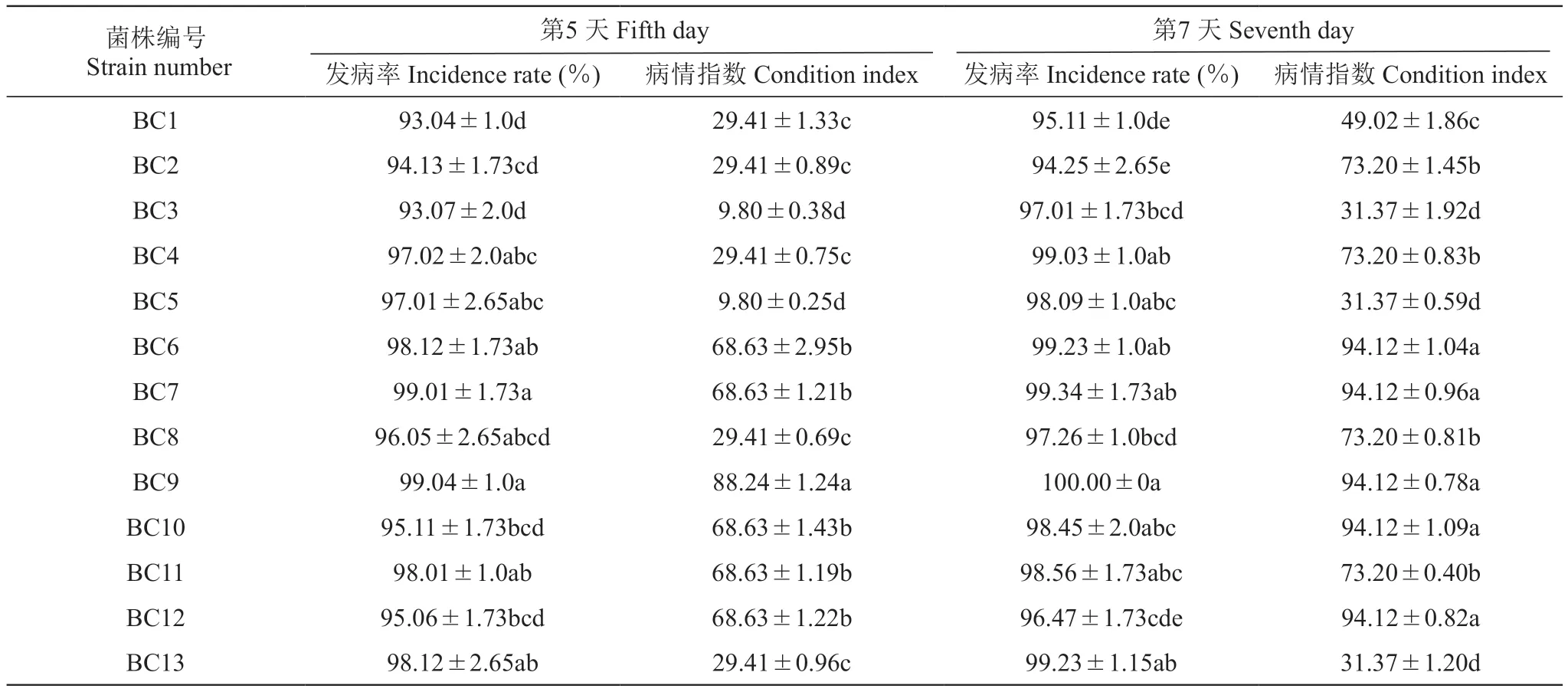
表5 13株葡萄灰霉病菌致病性測定Table 5 Pathogenicity determination of 13 strains of Botrytis cinerea on the 5th day and 7th day
2.5 類群分析
3、棚溫控制:下種后棚溫白天保持在30~35℃之間,夜間保持在15℃以上,注意下種后棚內溫度不能超過38℃以上,因為達到40℃高溫時,種子不利于發芽。根據實踐經驗,幼苗出土后,如果溫度超過26℃時,下胚軸會急速伸長,而形成高腳苗,所以幼苗拱土后,白天棚內溫度降到25~22℃之間,特別要注意的是此時晚間棚內溫度不能超過10℃以上,不能低于5℃,控制夜間苗不生長,棚內高溫、高濕會導致幼苗徒長。第一片真葉見長后白天要適當加大棚內溫度,溫度白天保持在28~33℃之間,夜間5~10℃之間。真葉見長后噴一次“金元寶”液肥,可增加植株葉綠素含量及生根量,使幼苗生長旺盛健壯,提高幼苗抗寒能力。
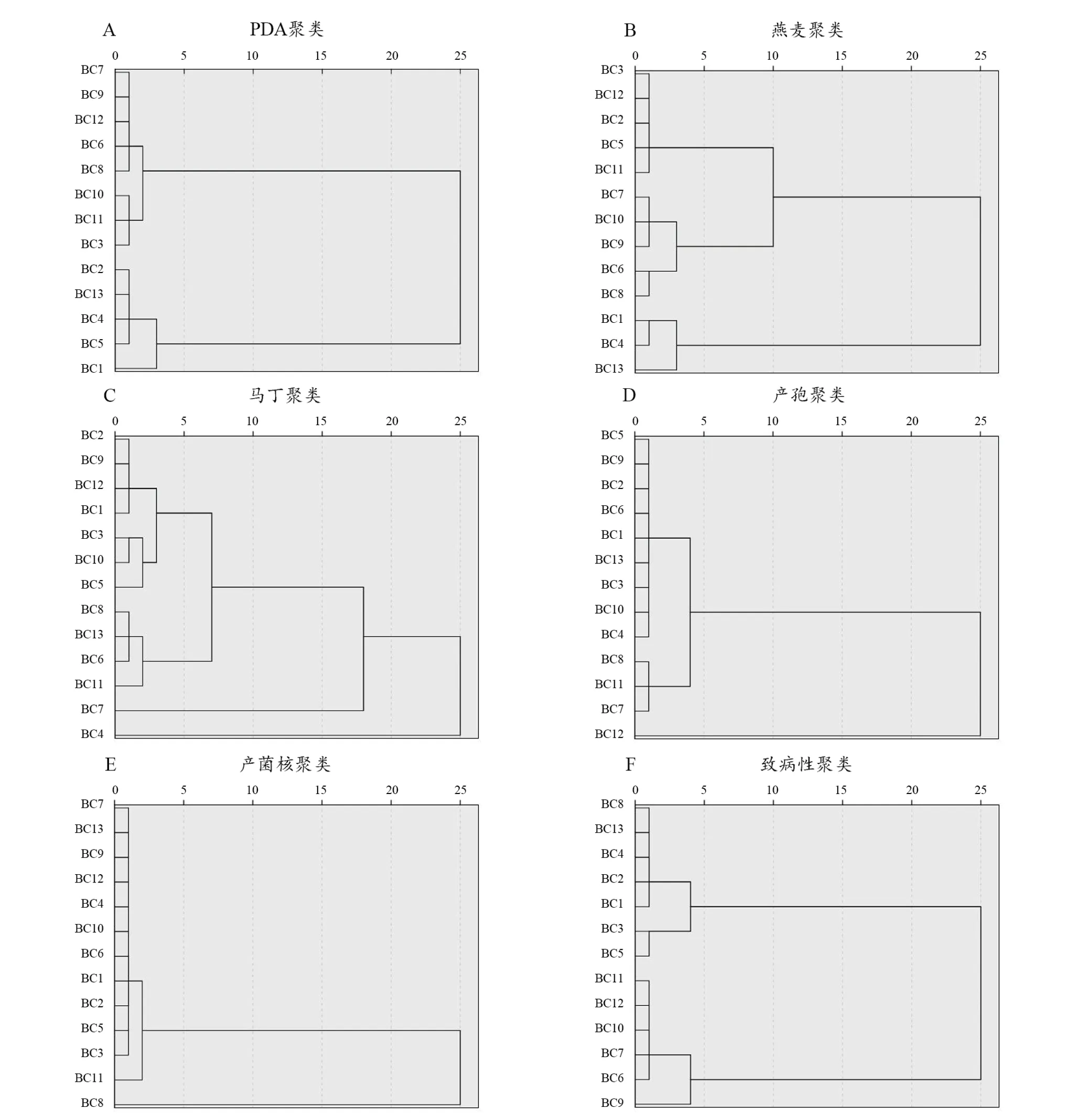
圖3 13株葡萄灰霉菌聚類分析圖Figure 3 Cluster analysis of 13 strains of Botrytis cinerea
依據13株葡萄灰霉病菌在3種培養基上的生長速率和形態、產孢量、產微菌核情況、致病性差異將13株葡萄灰霉病菌單孢菌株劃分為8個類群。其中菌株BC6、BC9、BC10和BC11歸為Ⅰ類群,菌落表現為菌絲生長速率較快,菌絲濃密,既可以產生分生孢子又可產生菌核,其菌絲呈現灰白色的灰霉病菌類群。菌株BC2、BC5歸為Ⅱ類群,菌落表現為菌絲生長速率較快,菌絲呈現灰白色,即產生分生孢子又產生菌核的灰霉病菌類群。菌株BC1和BC13歸為Ⅲ類群,菌落表現為菌絲生長速率中等,菌絲較稀疏,菌絲顏色呈現灰白色的灰霉病菌類群,菌株BC1產生分生孢子量最低,BC13既不產生分生孢子也不產生菌核。菌株BC3歸為Ⅳ類群,菌落表現為菌絲生長速率較慢,菌絲較稀疏,菌絲顏色呈現灰白色的灰霉病菌,即可產生分生孢子也可產生菌核。菌株BC4歸為Ⅴ類群,菌落表現為菌絲生長速率較快,且菌絲稀疏,菌落顏色呈現灰白色的灰霉病菌類群,即可產生分生孢子也可產生菌核。菌株BC7歸為Ⅵ類群。菌株BC8歸為Ⅶ類群。BC7和BC8菌落表現均為菌絲生長速率較慢,且菌絲稀疏。在PDA培養基上培養30 d后BC7不產生菌核,BC4在培養28 d后產生菌核,BC8在培養29 d后產生菌核。菌株BC12歸為Ⅸ類群,菌落表現為菌絲生長速率較快,且菌絲濃密,菌落顏色呈現灰白色的灰霉病菌類群,即可產生分生孢子也可產生菌核,產孢量最高。
巴志鵬認為,中國共產黨的生態文明建設思想是對中國古代樸素的人與自然和諧觀“天人合一”及樸素的自然倫理“仁民愛物”論等傳統生態文化的揚棄和對馬克思恩格斯的人與自然協調發展觀的傳承[19]。而榮開明則認為,生態文明建設思想的主要淵源,包括馬克思主義生態文明理論,黨的三代領導集體探索中國社會主義建設的論述和前期經驗,國外有關生態文明建設的理念和經驗,中華文明中的生態文明智慧[20]。
2.6 葡萄灰霉病菌的分子鑒定
通過對8 株代表性葡萄灰霉病菌的B C7 2 9、BChch、Rpb2和G3pdh進行擴增,結果都能夠擴增出目的基因片段。由圖4可知,分別將8株葡萄灰霉病菌代表菌株依據BC729、BChch、Rpb2和G3pdh四組基因序列構建的系統發育樹。通過4組基因鑒定,證明其均為灰葡萄孢B. cinerea,但是8株葡萄灰霉病菌的基因堿基序列均有差異,與形態學類群劃分結果一致。
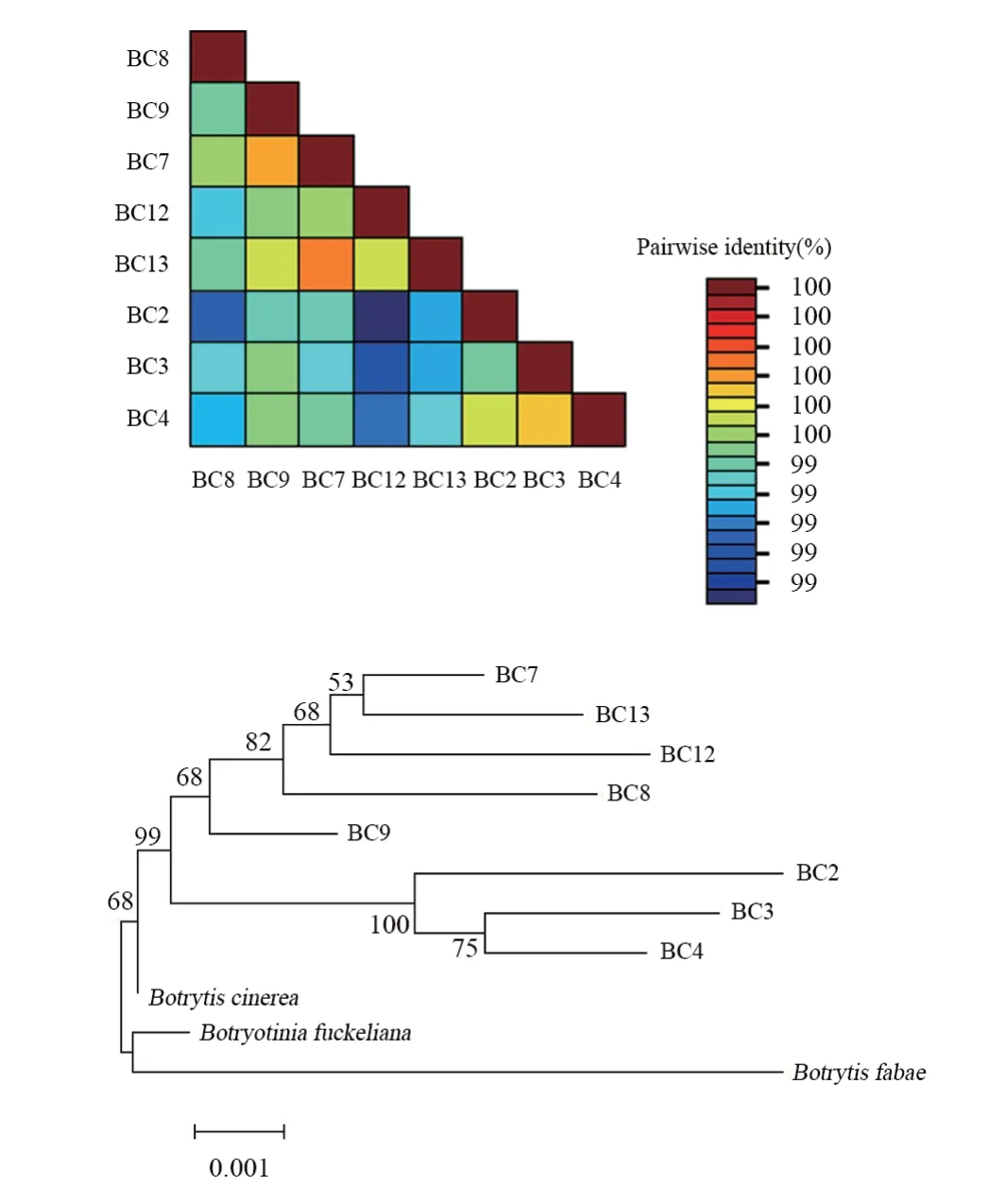
圖4 8株葡萄灰霉病菌菌株基因間的相關性熱圖及構建的系統發育樹Figure 4 Correlation heat map and phylogenetic tree of eight strains of Botrytis cinerea
應用SDTv.1.2軟件分析了8株代表性菌株與灰霉病菌株的BC729、BChch、Rpb2和G3pdh基因拼接序列間核酸的一致性。由圖4可知,菌株BC9與其他灰霉病菌株之間的一致性為99.40%;菌株BC7與其他灰霉病菌株之間的一致性為99.50%~99.70%;菌株BC12與其他灰霉病菌株之間的一致性為99.20%~99.50%;菌株B C 1 3 與其他灰霉病菌株之間的一致性為99.40%~99.70%;菌株BC2與其他灰霉病菌株之間的一致性為98.90%~99.30%;菌株BC3與其他灰霉病菌株之間的一致性為99.0%~99.40%;菌株BC4與其他灰霉病菌株之間的一致性為99.10%~99.60%。
剛走到大街上,就看見村東邊燃放了一個鉆天猴,這個鉆天猴鉆得真叫一個高,搖頭擺尾像火箭一樣直沖上天去了,到了最高處還叭地響了一下。我心里就踏實了。這是我跟劉鐵頭約定的暗號,這說明他已經得手了,他和李金枝已經像鉆天猴一樣鉆上天了。
3 討論
灰葡萄孢(B. cinerea)可通過侵染葡萄植株的葉片、枝干、花和果實導致葡萄灰霉病的發生[15]。其侵染植株后,引起植株體細胞程序性死亡,從而破壞組織正常生長,屬于植物致病的死體營養型病原真菌類型[16]。目前,噴施化學農藥是防治灰霉病的主要方法[17],但是灰葡萄孢為復合種,其種內不同類群之間的致病性和抗藥性都存在很大的差異[18],因此研究其類群劃分對于灰霉病的有效防治非常重要。
目前常利用形態學和致病性鑒定對病原菌進行類群劃分[19]。黃燕等[20]對來源于中國6個不同地區的100株蠶豆灰葡萄孢B. fabae進行形態學鑒定,根據菌落形態可分為菌核型、菌絲型和分生孢子型,其中76個分離物為菌核型。周梓良[21]對湖北省17個地區采集的灰霉病樣品進行離體葉片測定,將942株灰霉病菌的致病力分為強致病力和弱致病力兩類,其中強致病力(病斑直徑>10 mm)灰霉病菌的比例高達93.94%,而弱致病力菌株(病斑直徑≤10 mm)的比例約為6.06%。王賀[22]通過致病性劃分將54個水稻稈腐菌核病菌Helminthosporium sigmoideum劃分為4個致病類群,并發現鹽堿地區水稻稈腐菌核病病原菌致病性高于非鹽堿地區。本研究以從寧夏賀蘭山東麓地區不同葡萄品種葡萄灰霉病病果上分離的13株葡萄灰霉病菌,通過對比其在馬鈴薯葡萄糖瓊脂培養基、燕麥瓊脂培養基、改良馬丁培養基三種不同培養基上的生長速率和形態、產孢量、微菌核和致病性差異劃分為8個類群,比王忠興[23]的劃分更精確和細致。
單基因鑒定對于一些親緣關系較近的種仍不能進行有效區分,因此普遍采用多基因分子鑒定[24]。Yan等[25]從我國6個省的葡萄園中分離得到34個菌株,通過多基因(ACT、ITS、GAPDH、TUB2和CHS)分析結合形態學分析表明,黑刺盤孢(Colletotrichum aenigma)、河北刺盤孢(C. hebeiensis sp. nov.)和炭疽菌屬(C. viniferum)與我國葡萄炭疽病有一定關聯。張佳等[26]根據草莓灰霉病菌(B. cinerea)的RPB2、G3PDH和HSP60的單基因和多基因系統發育分析結果發現,我國草莓灰霉病的病原菌至少包括3個系統發育種。侯圣凡等[27]對來自全國12個草莓產區的109株草莓根腐病樣品通過分子鑒定、多基因序列(ITS-ACT-GAPDH-CALTUB2-CHS)聯合鑒定發現,有27.5%的樣品是由草莓炭疽根腐病病原菌膠孢炭疽菌復合種(Colletotrichum gloeosporioides complex)引起。本研究通過對8株代表性葡萄灰霉病菌的BC729、BChch、Rpb2和G3pdh進行擴增,表明都為灰葡萄孢,但是基因序列略有差異,驗證了形態學劃分的結果。
本研究對分離自寧夏賀蘭山東麓不同葡萄品種的13株葡萄灰霉病菌通過形態學方法對其類群進行劃分為8大類群,并通過分析BC729、BChch、Rbp2和G3pdh4個基因序列,對其進行多基因分子鑒定,顯示這些菌株均為灰葡萄孢,但是堿基序列有差異,與形態學類群劃分結果一致。本研究通過形態學觀察和分子鑒定手法,明確賀蘭山東麓地區灰霉病菌的種群結構,為該地區葡萄灰霉病的有效防治提供理論依據。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
