花生種子在低溫脅迫下的萌發(fā)抑制生理研究
2023-05-19 01:49:54王藝喆蘇良辰
種子 2023年2期
王藝喆, 蘇良辰
(遵義醫(yī)科大學(xué)珠海校區(qū), 廣東 珠海 519041)
花生(ArachishypogaeaL.)是我國重要的油料作物和經(jīng)濟作物之一,種植面積常年保持在450萬hm2以上,僅次于印度(800萬hm2),居世界第2位[1]。萌發(fā)期是花生生產(chǎn)過程中最重要也是最脆弱的一個階段,在此過程中種子遭受環(huán)境脅迫會對花生產(chǎn)量造成嚴重影響[2-3]。其中,低溫對花生種子萌發(fā)和幼苗生長發(fā)育有重要影響,也是影響花生春季和秋季播種的重要因素,極大制約了我國花生生產(chǎn)的種植面積[4-5]。
花生種子萌發(fā)是一個多因素參與的復(fù)雜調(diào)控過程,其中,抗氧化酶能清除種子萌發(fā)過程中產(chǎn)生的自由基及氧化中間產(chǎn)物,維持細胞自由基的低水平狀態(tài)并保護細胞膜不受損傷,為種子正常萌發(fā)提供適宜的條件[6-8]。同時,植物激素互作在種子萌發(fā)過程中發(fā)揮關(guān)鍵調(diào)控作用。赤霉素能增強細胞伸長并促進胚的生長,而脫落酸則抑制種子萌發(fā)、促進種子休眠[9]。細胞分裂素能夠解除種子休眠、促進萌發(fā),增強胚的膜透性及促進赤霉素釋放[10]。赤霉素和脫落酸的拮抗作用調(diào)控著種子萌發(fā)的整個過程,赤霉素能夠直接或者間接抵消脫落酸的抑制作用,而脫落酸又能抑制赤霉素合成[11-12]。因此,花生種子中抗氧化酶活性和植物激素水平變化與低溫條件下種子的萌發(fā)關(guān)系密切。
本研究通過測定低溫條件下的粵油7號花生種子中滲透調(diào)節(jié)物質(zhì)含量、抗氧化酶活性等生理生化指標(biāo)和萌發(fā)相關(guān)植物激素合成基因的相對表達量,探討了花生萌發(fā)時期低溫對于種子生理生化的影響規(guī)律,為深入研究花生的低溫栽培育種提供實驗基礎(chǔ)。
1 材料與方法
1.1 植物材料與培養(yǎng)
供試花生品種為粵油7號,由實驗室保存。將帶殼種子分別置于25 ℃和15 ℃培養(yǎng)箱中7 d以上。選取顆粒飽滿未發(fā)芽的種子洗凈,轉(zhuǎn)移至超純水浸濕濾紙的培養(yǎng)皿上。分別在25 ℃(常溫處理組)和15 ℃(低溫處理組)培養(yǎng)箱(光照0 h,濕度70%)中培養(yǎng),每組50粒花生。分別在0 h,24 h,48 h取花生胚,液氮速凍后于-80 ℃冰箱保存?zhèn)溆?用于后續(xù)生化指標(biāo)和基因相對表達量的測定。
1.2 實驗方法
1.2.1發(fā)芽率的測定
胚芽突破種皮定為發(fā)芽,每個組做3個重復(fù),每個培養(yǎng)皿中50粒花生種子,從第一天開始每天統(tǒng)計種子發(fā)芽數(shù)。
發(fā)芽率/%=(發(fā)芽種子數(shù)/培養(yǎng)皿種子總數(shù))×100%。
1.2.2胚中各項生化指標(biāo)的檢測
超氧化物歧化酶(SOD)、丙二醛(MDA)和蛋白濃度分別按照碧云天公司的S 019、S 0131和P 0012 S試劑盒進行檢測。脯氨酸(Pro)、過氧化物酶(POD)和可溶性糖分別按照Solarbio公司的BC 0295、BC 0095、BC 0030試劑盒進行檢測。
1.2.3基因表達量檢測
利用FastPure Plant Total RNA試劑盒提取總RNA,M-MuLV第一鏈cDNA合成試劑盒進行cDNA合成。采用實時定量PCR技術(shù)檢測胚中AhYUC1、AhGA20OX1、AhIPT5、AhACS1和AhNCED1的表達量,引物見表1,使用儀器為MX 3500 P Real-time PCR system。PCR反應(yīng)條件為:95 ℃預(yù)變性30 s;95 ℃變性10 s;60 ℃退火延伸30 s,共40個循環(huán)。每個樣品3次重復(fù),最后取平均值。
1.2.4數(shù)據(jù)處理
采用SPSS 25.0統(tǒng)計軟件處理數(shù)據(jù),多組間的差異采用單因素方差分析,p<0.05表示有顯著性差異。用Origin 2019軟件作圖。
2 實驗結(jié)果
2.1 不同溫度處理下的花生種子萌發(fā)率
實驗結(jié)果表明,25 ℃培養(yǎng)24 h和48 h時種子萌發(fā)率分別為60.00%和76.67%,種子正常萌發(fā)。而在15 ℃低溫培養(yǎng)48 h內(nèi)沒有種子萌發(fā)(圖1)。說明花生種子萌發(fā)受溫度影響較大,種子在低溫條件下無法正常萌發(fā)。

注:紅色標(biāo)尺表示長度。圖1 不同溫度下花生種子萌發(fā)情況Fig.1 Seed germination at different temperatures
2.2 不同溫度對花生萌發(fā)過程中MDA含量的影響
MDA是膜脂過氧化的主要產(chǎn)物,可以反映細胞或組織中膜脂的過氧化程度。由圖2可知,不同溫度條件下培養(yǎng)的花生種子中MDA含量都是先上升后下降,但低溫條件下的花生種子變化幅度明顯小于常溫組。同0 h相比,24 h處理后,常溫處理組與低溫處理組分別上升了395.79%、121.23%;48 h處理后,常溫處理組上升了37.89%,而低溫處理組下降了10.22%。隨著種子的萌發(fā),種子膜脂過氧化程度提高,MDA含量增加,達到峰值后開始減小。
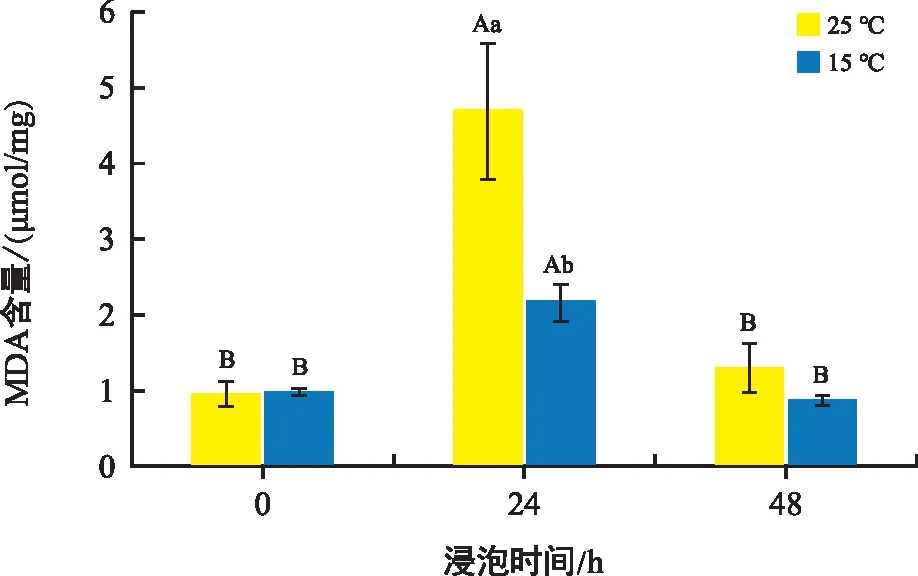
注:不同大寫字母表示相同溫度不同時間下具有顯著性差異,不同小寫字母表示相同時間不同溫度下具有顯著性差異,p<0.05。下同。圖2 不同溫度和時間下花生胚中MDA含量變化Fig.2 Changes in malondialdehyde content in peanut embryos at different temperatures and times
2.3 不同溫度對花生萌發(fā)過程中Pro含量的影響
由圖3可知,常溫處理24 h和48 h的種子Pro含量較0 h分別上升了41.70%和30.89%。低溫處理組的種子Pro含量在24 h和48 h較0 h分別下降40.96%和60.31%,下降趨勢明顯。
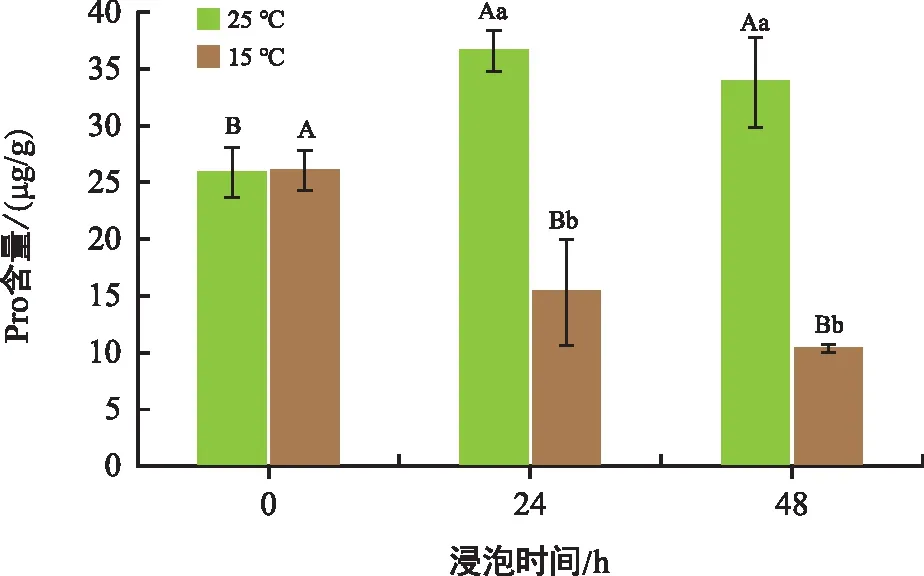
圖3 不同溫度和時間下花生胚中Pro含量變化Fig.3 Proline content in peanut embryos changes at different temperatures and times
2.4 不同溫度對花生萌發(fā)過程中POD活性的影響
由圖4可知,常溫處理組花生種子中的POD含量在各時間點沒有明顯變化。但是在低溫處理組中POD的含量隨著種子培養(yǎng)時間的延長含量不斷上升,且低溫組中的POD含量均高于常溫組。
2.5 不同溫度對花生萌發(fā)過程中SOD活性的影響
SOD是生物體內(nèi)的一種抗氧化金屬酶,它能夠催化超氧陰離子自由基歧化生成氧和過氧化氫,在細胞氧化與抗氧化平衡中起到至關(guān)重要的作用。由圖5可以看出,與對照相比,常溫處理組SOD活性在24 h、48 h分別上升了34.49%、18.24%,而低溫處理組分別上升了8.83%、14.39%。隨著培養(yǎng)時間的延長,SOD活性都有所增加。0 h、24 h、48 h時低溫處理組的SOD活性分別比常溫處理組高42.52%、15.33%、37.89%。
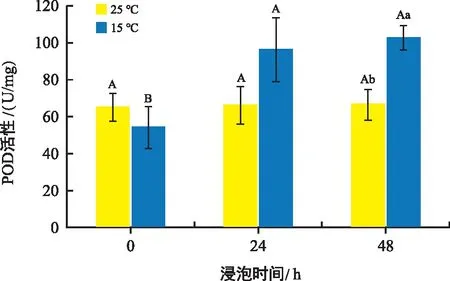
圖4 不同溫度和時間下花生胚中POD活性變化Fig.4 Changes in peroxidase content in peanut embryos at different temperatures and times
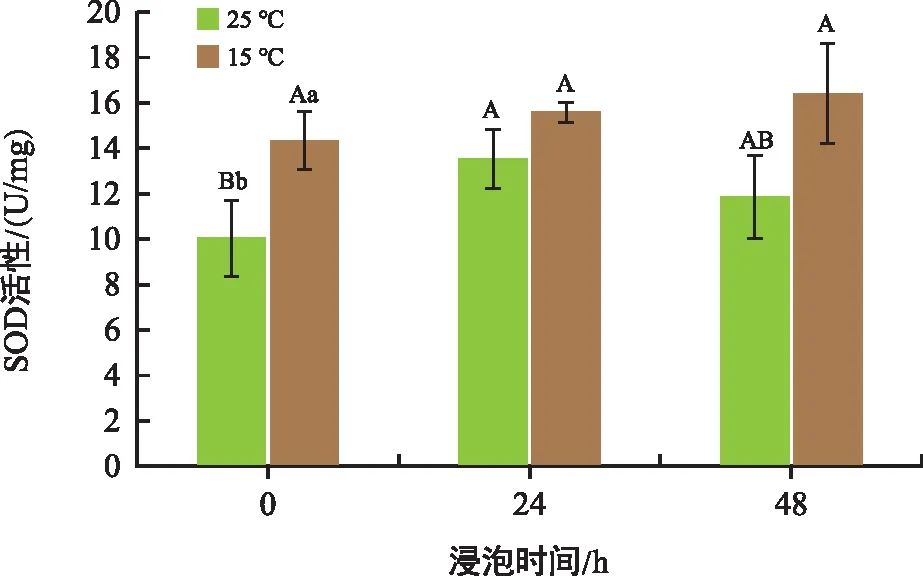
圖5 不同溫度和時間下花生胚中SOD活性變化Fig.5 Changes in superoxide dismutase content in peanut embryos at different temperatures and times
2.6 不同溫度對花生萌發(fā)過程中可溶性蛋白含量的影響
植物可溶性蛋白和可溶性糖都是植物細胞內(nèi)調(diào)節(jié)滲透壓的物質(zhì),還可以作為防凍劑或穩(wěn)定劑,往往作為細胞抗脅迫生理指標(biāo)。由圖6可知,與0 h相比,常溫組24 h、48 h組可溶性蛋白濃度下降了52.58%、65.69%,低溫組24 h、48 h組蛋白濃度分別下降了53.73%、61.78%。兩組隨著培育時間的延長可溶性蛋白濃度都有所下降,但組間無顯著差異。
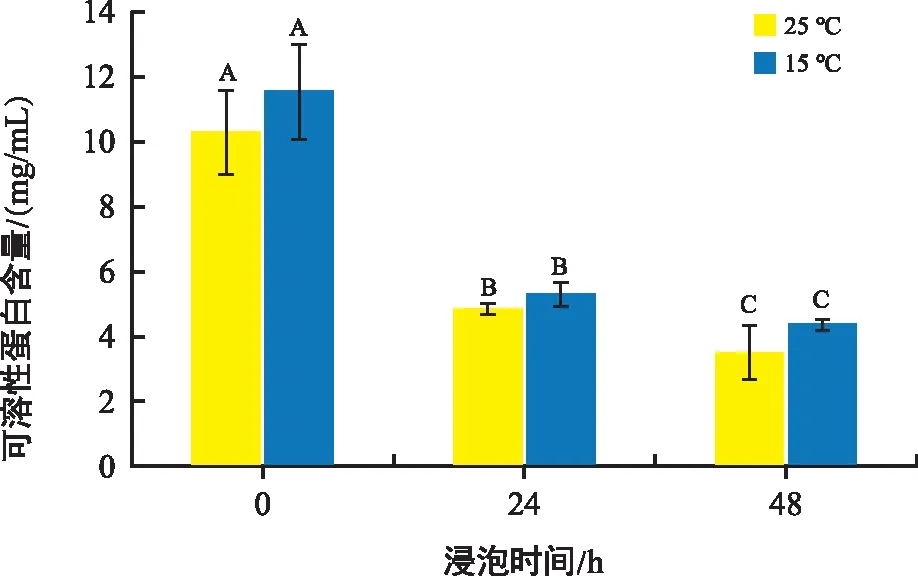
圖6 不同溫度和時間下花生胚中可溶性蛋白含量變化Fig.6 Changes in protein content in peanut embryos at different temperatures and times
2.7 不同溫度對花生萌發(fā)過程中可溶性糖含量的影響
如圖7所示,在低溫脅迫下,低溫組的可溶性糖含量高于常溫組。相比于0 h,常溫處理24 h、48 h組可溶性糖含量分別降低63.67%、58.60%,低溫組24 h、48 h分別降低了29.39%、37.85%。低溫處理組中可溶性糖下降幅度較小。相同處理時間條件下,低溫組的可溶性糖含量明顯高于常溫組。
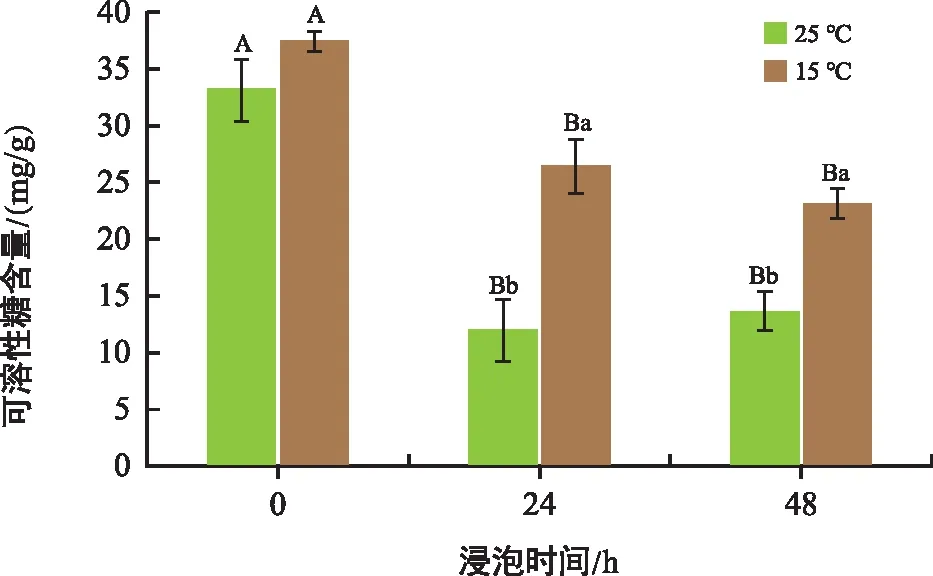
圖7 不同溫度和時間下花生胚中可溶性糖含量變化Fig.7 Changes in soluble sugar content in peanut embryos at different temperatures and times
2.8 低溫對花生萌發(fā)過程中相關(guān)植物激素合成基因表達的影響
為探究低溫脅迫對花生種子萌發(fā)中內(nèi)源激素基因的表達變化,分析了花生AhYUC1(生長素合成相關(guān)基因)、AhGA20OX1(赤霉素合成相關(guān)基因)、AhIPT5(細胞分裂素合成相關(guān)基因)、AhACS1(乙烯合成相關(guān)基因)和AhNCED1(脫落酸合成相關(guān)基因)在不同溫度下的基因相對表達量。由圖8可以看出,AhYUC1、AhGA20OX1、AhIPT5、AhACS1和AhNCED1基因表達水平受低溫脅迫誘導(dǎo)。在低溫脅迫24 h后,花生胚中AhYUC1基因表達量上調(diào)7.19倍,AhGA20OX1基因表達量上調(diào)96.11倍,AhIPT5基因表達量上調(diào)80.08倍,AhACS1基因表達量上調(diào)4.73倍,AhNCED1基因表達量上調(diào)了2.32倍。其中AhYUC1和AhGA20OX1的表達趨勢一致,表達量逐漸升高,AhIPT5、AhACS1和AhNCED1的表達趨勢一致,都是先升高后降低,在24 h達到峰值。常溫處理組,AhYUC1、AhGA20OX1和AhIPT5的變化趨勢一致,表達量先上升后下降,在24 h達到峰值,表達量分別上調(diào)1.29倍、16.19倍和8.07倍,AhACS1和AhNCED1的表達趨勢一致,逐漸下降,在處理48 h后達到最低,表達量分別下調(diào)了90.00%和96.00%。

圖8 低溫對花生胚AhYUC1、AhGA20OX1、AhIPT5、AhACS1、AhNCED1表達的影響Fig.8 Effect of low temperature on the expression of peanut embryos AhYUC1,AhGA20OX1,AhIPT5,AhACS1 and AhNCED1
3 討 論
植物中發(fā)揮關(guān)鍵保護作用的抗氧化酶類主要為SOD和POD。研究表明,種子在低溫下的萌發(fā)率和抗氧化酶含量與植物的抗氧化能力成正比,同時與MDA等過氧化代謝產(chǎn)物含量成反比。Aiju等[13]在玉米萌發(fā)的研究中發(fā)現(xiàn),13 ℃低溫預(yù)處理4 d的玉米種子較常溫培養(yǎng)的種子總抗氧化能力明顯下降。并且在抗性品系中,1~6 d的SOD活性顯著高于對照組,同時POD活性在總體水平上也顯著高于對照組;而MDA含量在抗性品系中表現(xiàn)出較低水平的累積[14]。本研究發(fā)現(xiàn),隨著萌發(fā)的進行,低溫處理組中的POD和SOD酶活性均明顯高于常溫處理組,呈不斷上升的變化趨勢。同時,低溫處理組中的MDA和Pro含量也顯著低于常溫處理組。可溶性蛋白和可溶性糖含量都會下降,但低溫組下降幅度小于常溫組,且低溫處理組的含量都高于常溫處理組。這表明花生種子在萌發(fā)過程中遭受低溫脅迫影響,種子顯著降低了細胞內(nèi)的代謝活動,并在細胞內(nèi)合成大量抗氧化物質(zhì)以降低有害物質(zhì)積累,保護種子活力,從而延緩種子萌發(fā)。
花生種子在不同溫度條件下的萌發(fā)活力受細胞內(nèi)多種植物激素的調(diào)節(jié)控制。據(jù)報道,赤霉素(Gibberellins,GAs)和脫落酸(Abscisic Acid,ABA)之間的植物激素平衡密切調(diào)控種子的萌發(fā)狀態(tài),兩者在多個水平上相互作用[15-16]。本研究中,花生種子無論在常溫條件還是在低溫條件下,AhGA20OX1和AhIPT5基因的表達量均出現(xiàn)不同程度的上調(diào),但在低溫條件下AhYUC1、AhGA20OX1和AhIPT5基因上調(diào)表達的幅度更為明顯。說明在不同的處理溫度下萌發(fā)促進因子均上調(diào)。萌發(fā)抑制因子在不同溫度條件的表現(xiàn)則顯示出明顯的差異,常溫處理組中的AhACS1和AhNCED1基因的表達量出現(xiàn)明顯的下調(diào),而低溫處理組中兩個基因顯著上調(diào)表達。這表明在低溫條件下,花生種子中的萌發(fā)抑制因子的上調(diào)表達影響了種子的萌發(fā)過程。綜合以上結(jié)果認為,低溫條件下種子的萌發(fā)延遲受到萌發(fā)抑制因子的影響最顯著。低溫脅迫下,花生種子達到了生長發(fā)育的新穩(wěn)態(tài),從而抑制了萌發(fā)進程,在這一過程中脫落酸和乙烯的生物合成在其中發(fā)揮了關(guān)鍵的抑制作用。此研究為提高花生種子在低溫脅迫條件下的萌發(fā)率提供前期實驗基礎(chǔ)。
