藍藻生物活性提取物的研究進展
2023-04-29 03:53:36田龍龍張聞定金小霞馮杰李婷婷
國外醫藥抗生素分冊 2023年3期
田龍龍 張聞定 金小霞 馮杰 李婷婷

摘要:藍藻又稱藍綠藻,是可以進行光合作用的原核生物,被認為是新的生物活性物質的重要來源。藍藻的多種天然產物在抗菌、殺蟲、抗病毒以及抗癌等方面展現出不同的生物活性。本文介紹了抗菌、殺蟲、抗病毒、抗癌等藍藻提取物的研究進展,以期為藍藻活性產物的進一步研究與開發提供幫助和新思路。
關鍵詞:藍藻;天然產物;抗菌;殺蟲;抗病毒;抗癌
中圖分類號:R978.1? ? ? ? 文獻標志碼:A? ? ? ? ?文章編號:1001-8751(2023)03-0202-09
Research Progress of Bioactive Extracts from Cyanobacteria
Tian Long-long1,? ?Zhang Wen-ding1,? ?Jin Xiao-xia1, Feng Jie1,2, Li Ting-ting1
(1 School of Basic Medicine, Lanzhou University, Lanzhou? ?730000;
2 State Key Laboratory of Veterinary Etiological Biology,
College of Veterinary Medicine, Lanzhou University,? ?Lanzhou? ?730000)
Abstract: Cyanobacteria, also known as blue-green algae, are photosynthesizing prokaryotes and are considered to be in great potential for providing substances with unique bioactivities. In this paper, the research progress of antibacterial, insecticidal, antiviral and anticancer extracts of cyanobacteria was reviewed, in order to provide help and new ideas for further research and development of cyanobacteria active products.
Key words: cyanobacteria; natural product; antibacterial; disinsection; antiviral; anticancer
藍藻是可以進行光合作用的原核生物,又稱藍綠藻或藍細菌[1]。大多數藍藻的細胞壁外面有膠質衣,因此又名粘藻。藍藻主要包括藍球藻、顫藻、念珠藻和發菜等。藍藻在自然界中分布十分廣泛,可在多種環境中生存,在生態系統中發揮著獨特的作用[2]。藍藻營養需求低,易于培養,某些菌株具有固氮的能力[3]。藍藻富含肽、蛋白質、脂質、維生素、色素、碳水化合物、萜類化合物、多不飽和脂肪酸、類黃酮、酚類化合物等多種化合物,是潛在的新型生物活性化合物的“生產工廠”[4]。藍藻所產的多種活性物質在抗菌、殺蟲、抗病毒、抗癌中都有極大的應用前景[5]。
細菌、寄生蟲、病毒感染以及腫瘤細胞侵襲嚴重危害著人類健康,同時由于耐藥問題,現有的抗感染、抗癌藥物治療效果有不同程度的下降。隨著抗菌藥物的大量使用,細菌耐藥的問題日趨嚴重,2022年世界衛生組織報告顯示,大腸埃希菌對環丙沙星的耐藥率高達92.9%,肺炎克雷伯菌對環丙沙星的耐藥率高達79.4% [6]。寄生蟲感染引起的疾病也在全球范圍蔓延,2019年至2020年期間,瘧疾感染人數達1400萬,死亡人數達6.9萬[7]。然而傳統使用的抗瘧藥物已出現了耐藥問題[8]。病毒也對人類健康造成了巨大的挑戰,2020年全球艾滋病死亡人數約150萬[9],埃博拉病毒(Ebola virus,EBOV)可引起致死性的中風、心肌梗塞、低血容量休克或多發性器官衰竭,患者病死率高達50%至90%[10],嚴重急性呼吸系統綜合征冠狀病毒2型(Severe acute respiratory syndrome coronavirus 2,SARS-CoV-2)導致約660萬人死亡[11]。此外,腫瘤細胞侵襲是人類死亡的主要原因,而且新型癌癥的發病率正在迅速增加,但腫瘤細胞對目前可用的藥物(如長春花生物堿和紫杉烷類)產生耐藥[12],因此也需要開發新的抗癌藥物。綜上所述,隨著細菌、寄生蟲、病毒以及腫瘤細胞對藥物的耐藥問題日趨嚴重,開發新的抗感染、抗癌藥物迫在眉睫,然而活性化合物資源枯竭導致新藥的研發速度越來越慢。目前的研究已從藍藻中分離發現了多種具有抗感染及抗癌活性的化合物[13],藍藻有望成為活性化合物藥物開發的新來源。本綜述將對藍藻天然產物的抗菌、殺蟲、抗病毒以及抗癌作用的最新研究成果進行歸納和總結,以利于進一步研究和開發藍藻活性物質。
1 藍藻提取物的抗菌及抗炎活性研究
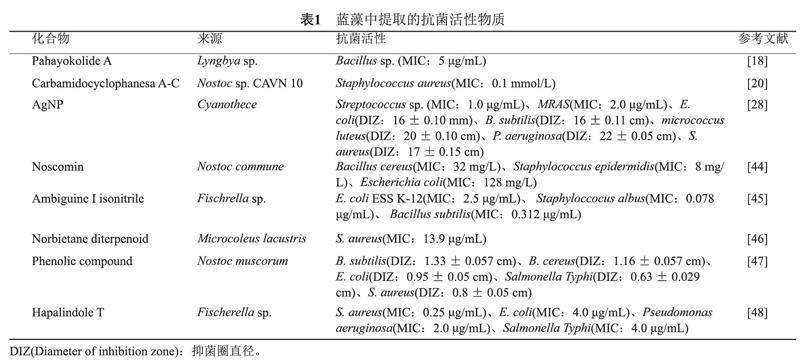
抗菌肽(Antimicrobial peptides , AMPs)是一類具有抗菌活性的多肽。研究表明,它們主要通過破壞細菌細胞膜完整性對細菌起到殺傷作用[14-15]。從淡水菌株鞘絲藻(Lyngbya sp.)中鑒定出的環肽Lyngbyazothrins C和Lyngbyazothrins D對革蘭陰性菌(大腸埃希菌、銅綠假單胞菌和黏質沙雷菌)和革蘭陽性菌枯草芽孢桿菌都具有殺菌作用[16]。研究者還從鞘絲藻屬中發現了Pahayokolide A和Pitipeptolide A-F這七種抗菌肽[17]。Pahayokolide A可以抑制枯草芽孢桿菌、巨大芽孢桿菌、蠟樣芽孢桿菌的生長[18]。Pitipeptolide A-F對結核分枝桿菌有殺菌作用[17],2020年,全球范圍內約有1000萬人罹患結核病,估計有150萬人死于結核病[9],而Pitipeptolide A-F有望進一步開發結核病防治新藥物。相較于化學合成的抗菌肽,藍藻產生的多肽綠色環保,合成成本低。另外,與核糖體多肽相比,藍藻產生的非核糖體多肽常被非核糖體肽合成酶修飾,結構更穩定,不易被蛋白酶降解,不易產生耐藥[15]。且藍藻營養需求低,易于培養,能夠產生的肽類豐富,合成可塑性強 ,因此藍藻有望成為更好的抗菌肽新來源。
我們對多種來源于念珠藻(Nostoc sp.)、費氏藻(Fischrella sp.)和鞘絲藻的抗菌活性物質進行了總結(見表1)。近年來,耐甲氧西林金黃色葡萄球菌(Methicillin-resistant Staphylococcus aureus,MRSA)的感染不斷增多,2020年全國細菌耐藥監測報告顯示MRSA全國平均檢出率為29.4%[19]。而從念珠藻 CAVN 10中分離出來的Carbamidocyclophanes對MRSA表現出抗菌活性,其最小抑制濃度MIC為0.1 mmol/L[20]。此外,研究者利用絲狀藍藻VPG 16-59的天然產物合成了新的抗菌藥物Anaephenes A和B,該藥物對MRSA也具有抗菌活性(MIC:8 μg/mL)[21]。這些化合物可能為MRSA的臨床治療提供新的方案。據報道,從費氏藻屬中分離出的Ambiguine-I isonitrile對枯草芽孢桿菌和白葡萄球菌表現出比鏈霉素更強的抗菌活性,其MIC分別為0.312 μg/mL、0.078 μg/mL[5]。此外,研究人員對藍藻提取物的活性成分進一步分析,發現藍藻產生的脂質α-亞麻酸(α-Linolenic acid)對金黃色葡萄球菌具有抗菌活性(MIC:15.6 mg/L)[22]。
另一方面,目前發現的300多種海洋藍藻生物堿中有很多非核糖體多肽或混合聚酮—非核糖體多肽生物合成途徑的產物[15],通過該途徑合成的天然產物包括許多目前臨床使用的藥物,例如抗菌藥物萬古霉素(糖肽類)、免疫抑制劑環孢霉素(非核糖體多肽)和抗癌劑博來霉素(糖肽類)等[23]。海洋藍藻中這些獨特的天然產物的發現,為此類新藥開發提供了除放線菌和真菌以外的又一重要來源。
不同于傳統的藥物,金屬納米顆粒是一種新型抗菌材料。金屬納米顆粒的主要抗菌機制是破壞細胞膜或細胞壁以及產生活性氧使蛋白質和核酸等生物大分子失活[24]。在金屬和金屬氧化物納米顆粒中,納米銀粒子(Argentum nanoparticles,AgNP)對各種病原體特別是耐藥細菌表現出獨特的抗菌活性[25]。金屬銀離子可以在藍藻細胞內與藻藍蛋白結合形成AgNP,也可以在藍藻細胞外與藻類多糖結合形成AgNP[26]。相比于傳統納米材料,藍藻AgNP體積更大,更易于提取使用[27]。藍藻合成的AgNP對巨大芽孢桿菌、大腸埃希菌、枯草芽孢桿菌、金黃色葡萄球菌、銅綠假單胞菌和MRSA均具有抗菌作用[26,28]。在一項比較研究中,利用海洋藍藻Phormidium formosum合成的AgNP 對金黃色葡萄球菌的抑菌圈值為2.2 cm,而抗菌藥物頭孢丙烯與多黏菌素對金黃色葡萄球菌的抑菌圈值分別為1.5 cm及2.0 cm [29],藍藻AgNP表現出更好的抗菌效果,由此可見藍藻AgNP的抗菌優勢。
廣譜β-內酰胺酶(Extended-spectrum β-lactamases,ESBL)是一類能水解多種青霉素類、頭孢菌素類以及單環β-內酰胺類抗菌藥物的酶。ESBL(+)菌可對上述抗菌藥物產生廣泛的耐藥性,從而導致難以治愈的菌血癥、腹腔內感染、尿路感染和呼吸道感染[30]。5%至25%的呼吸機相關性肺炎患者感染ESBL(+)菌后,會影響治療、預后甚至危及生命[31]。研究人員從海洋藍藻Accillatoria acuminata NTAPC05中提取的生物活性物質MGDG-palmitoyl對ESBL(+)的大腸埃希菌、嗜麥芽窄食單胞菌和阿氏腸桿菌B938的最低殺菌濃度MBC均為100 μg/mL,表現出較高的殺菌活性[32],這為抗ESBL(+)菌的藥物開發提供了新的方向。
此外,具有殺細菌、殺真菌作用的藍藻活性化合物也在開發外用殺菌劑以及農業殺菌劑方面有巨大的優勢和應用前景。一項研究發現魚腥藻屬(Anabaena)藍藻胞外蛋白酶對念珠鐮刀菌等植物病原真菌具有殺菌作用,可以作為植物病原真菌生物防治劑[33]。從藍藻中提取的Cryptophycin是對抗隱球菌的有效殺菌劑,目前已投入使用[34]。藍藻產生的殺菌化合物憑借高活性、易分解的特點,在有效殺滅細菌和真菌的同時能夠避免殺菌劑殘留,防止毒性物質在食物鏈中的富集和擴散。而且由于藍藻營養需求低,無須添加碳源,易于大規模培養,利用藍藻生產天然殺菌劑的方法明顯優于昂貴且不環保的化學合成方法[35]。
藍藻活性物質不僅具有抗菌活性,還可以調節炎癥反應。例如,海洋藍藻產物中的脂多糖分子CyP可以調節細菌感染引起的炎癥。人體細胞中的Toll樣受體4(Toll-like receptor 4,TLR4)和MD-2受體可以通過識別革蘭陰性菌的脂多糖(Lipopolysaccharide,LPS)激活機體免疫應答[36-37],合成促炎因子,誘發機體產生炎癥[38]。CyP可與LPS競爭性結合MD-2受體,進而抑制LPS與TLR4-MD-2受體復合物結合[36],從而選擇性抑制TLR4通路介導的細胞因子表達和一氧化氮(NO)的產生,調節炎癥反應。牙齦卟啉單胞菌的LPS(Pg-LPS)能夠促進牙周的慢性炎癥。研究發現,CyP可以抑制Pg-LPS誘導的炎癥,具體作用機制是阻斷人單核細胞中TLR4-MD-2通路,促進miR-146a(促炎反應的負調節因子)的表達,從而抑制腫瘤壞死因子(Tumor necrosis factor,? TNF)-α、白細胞介素(Interleukin,IL)-1β和IL-8等促炎因子生成[39]。牙齦卟啉單胞菌也在口腔癌的發病中發揮作用[40],CyP的上述抗炎活性可能對牙齦卟啉單胞菌相關的口腔癌的治療具有重大意義。又如,Malyngamides是一類衍生自海洋藍藻鞘絲藻屬巨大鞘絲藻(Lyngbya majuscula)的酰胺類化合物,該類化合物選擇性地抑制TLR4和TNF-α介導的髓樣分化因子依賴性通路中的蛋白質的活化和磷酸化,抑制了IL-1β、IL-6、IL-10、TNF-α和誘導型一氧化氮合酶轉錄,進而抑制巨噬細胞中NO等免疫因子參與的炎癥反應[40-41]。鞘絲藻酰胺的抑制炎癥作用使其成為潛在的免疫調節劑。從海洋藍藻鞘絲藻屬獲得的生物活性物質Grassystatins A可以與外源性抗原(如破傷風毒素)競爭性結合主要組織相容性復合體Ⅱ類分子,阻礙外源性抗原進入輔助性T淋巴細胞,從而抑制T細胞的增殖,抑制炎性細胞因子(IL17和干擾素-γ)的生成,減輕炎癥反應[42-43]。
2 藍藻提取物的殺蟲活性研究
2.1 藍藻提取物的殺傷寄生蟲活性研究
從藍藻菌株中發現的多種生物活性物質如Viridamide A、Venturamides、Dragomabin、Ambigol C可以有效殺傷寄生蟲(見表2)。研究發現Nostocarboline及其二聚體是惡性瘧原蟲的有效抑制劑,其半數抑制濃度(IC50)為194 nmol/L[49],此外,該化合物對布氏錐蟲、克氏錐蟲、杜氏利什曼原蟲均表現出殺蟲作用,且對大鼠L6細胞毒性極弱,有良好的藥物開發的潛力[49-50]。Clark等[50]從海洋藍藻中分離出6種新的酰基脯氨酸衍生物Tumonoic acids D-I均在抗瘧試驗中有效,其中,Tumonoacid I效果最好(IC50:2 μmol/L)。從藍藻束藻屬(Symploca sp.)獲得的Symplocamide A也對瘧原蟲有殺蟲效果(IC50:0.95 μmol/L)[51]。此外,Sanchez等[52]從巨大鞘絲藻中分離并鑒定出了一系列脂肽Almiramids A-C,這些脂肽在體外實驗中殺利什曼原蟲作用與殺錐蟲藥物硝呋替莫相同(IC50:3.0 μmol/L)。2015年,研究者從熱帶藍藻Okeania hirsuta中分離出一種新的化合物Bastimolide A,它對氯喹、甲氟喹、乙胺嘧啶和阿托伐醌耐藥的瘧原蟲有抑制活性[53],有望用于抗耐藥惡性瘧原蟲的藥物開發。
2.2 藍藻提取物殺傷蚊蟲活性的研究
蚊子作為瘧原蟲蟲媒可以傳播瘧疾,2021年世界范圍內通過按蚊傳播的瘧疾導致約61.9萬人死亡[54]。研究發現藍藻活性物質可以對多種蚊子起到強效殺蟲作用,進而減少瘧原蟲等寄生蟲的傳播。研究篩選了多種藍藻分離株及其提取物,發現藍藻Westiellopsis sp.提取物可以殺傷多種蚊子幼蟲,例如埃及伊蚊及致倦庫蚊等[55]。藍藻費氏藻ATCC 43239產生的吲哚生物堿Hapalindole在低劑量下對搖蚊幼蟲表現出殺傷作用[56]。來自鞘絲藻屬的化合物Pahayokolides對蚊子幼蟲也有殺傷作用[35, 57]。此外,從項圈藻(Anabaenopsis)和微囊藻屬(Microcytis sp.)的藍藻中獲得的藍藻毒素(Cyanobacterial toxins)、柱孢藻毒素(Cylindrospermopsin)和微囊藻毒素(Microcystins)等對蚊子幼蟲的殺滅率超過50%[35]。不同于傳統農藥,部分藍藻毒素與微囊藻毒素屬于肽類化合物,這些化合物容易被水解,在環境中不易長期殘留[58],因此該類化合物在殺滅蚊子幼蟲的同時能夠有效避免毒素殘留,屬于環境友好型殺蟲劑。
3 藍藻提取物抗病毒活性的研究
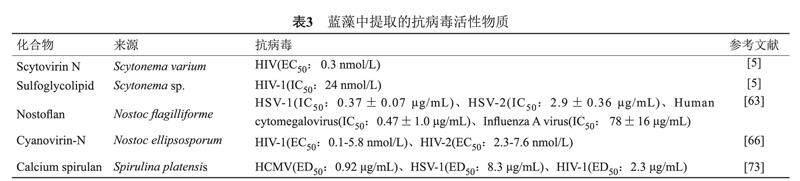
藍藻提取物對人類免疫缺陷病毒(Human immunodeficiency virus,HIV)、單純皰疹病毒(Herpes simplex virus,HSV)、丙型肝炎病毒(Hepatitis C virus,HCV)、EBOV和一系列其他包膜病毒顯示出抗病毒活性[63-66](見表3)。從念珠藻中提取的兩種糖結合蛋白,藍藻抗病毒蛋白N(Cyanovirin-N,CV-N)和Scytovirin N(SVN)有望成為新型的殺病毒藥物。CV-N在體外和體內均表現出抗HIV和其他慢病毒的活性[67-68]。此類糖結合蛋白可以干擾病毒感染細胞過程中的多個步驟,如通過干擾HIV gp120蛋白與CD4+受體、靶細胞趨化因子受體的結合來抑制HIV病毒與CD4+細胞的吸附[67]。CV-N在發揮抗HIV病毒作用的同時對人體毒性小,有望開發成為抑制HIV性傳播的陰道凝膠,從而減少HIV-1的全球傳播[69]。此外,CV-N還被發現可在體外抑制單純皰疹病毒-6與靶細胞的吸附;抑制麻疹病毒的包膜糖蛋白與血凝素的結合;強烈阻斷貓類免疫缺陷病毒感染免疫細胞[67]。CV-N具有SARS-CoV-2的S糖蛋白的聚糖類型表征,利用分子對接和分子動力學模擬研究發現CV-N與SARS-CoV-2 S糖蛋白對接時形成了穩定的分子間氫鍵[70],被認為是SARS-CoV-2的潛在阻斷劑[71]。SVN具有廣泛的抗病毒活性,其主要機制是SVN與許多病毒的包膜糖蛋白上富含甘露糖的寡糖具有高親和力,可以與靶細胞上的受體分子競爭性結合病毒的包膜糖蛋白,從而阻止病毒進入靶細胞[65]。從偽枝藻屬(Scytonema varium)中提取分離出來的Scytovirin N可以與HIV的包膜糖蛋白(gp120、gp160和gp41)結合,在低濃度下就可使HIV病毒失活[69]。研究發現SVN可以與扎伊爾型埃博拉病毒(Zaire Ebola virus,ZEBOV)包膜糖蛋白上的黏蛋白區域特異性結合,抑制ZEBOV復制[65]。來自藍藻偽枝藻屬的天然硫糖脂也可以抑制HIV逆轉錄酶和DNA聚合酶的活性[72],從而起到抗HIV病毒的作用。
從藍藻鈍頂螺旋藻(Spirulina platensis)中分離出的Calcium spirulan(Ca-SP)是一種具有廣譜抗病毒活性的硫酸化多糖,可強烈抑制多種人類病毒如人巨細胞病毒(Human cytomegalovirus,HCMV)、HSV-1、麻疹病毒、腮腺炎病毒、甲型流感病毒和HIV-1的復制[73-74]。Ca-SP可抑制病毒附著并入侵宿主細胞;還可以抑制感染HIV和未感染HIV的CD4 +淋巴細胞之間的融合,從而極大減弱病毒的傳染性[63]。Hayashi等[74-75]評估了Ca-SP對HIV-1和HSV-1的抗病毒潛力,發現使用Ca-SP的艾滋病模型小鼠的血清樣品在24 h后沒有觀察到病毒誘導的合胞體形成。相較于其他硫酸化多糖,Ca-SP抗病毒效果強,抗凝血活性低,藥物動力學半衰期更長,Ca-SP可以作為抗HIV治療藥物的候選藥物。此外,從念珠藻中分離出的抗病毒多糖Nostoflan可以抑制病毒與細胞的相互作用,對HSV-1、HSV-2、人巨細胞病毒和甲型流感病毒顯示出有效的抗病毒活性[63]。
4 藍藻提取物的抗癌及抗肥胖活性研究
有研究顯示,從念珠藻屬GSV224中分離出來的念珠藻素-1可以干擾微管的組裝,導致有絲分裂停滯,阻斷G2或M期的細胞增殖,誘導癌細胞死亡[34]; 現已發現它對KB人鼻咽癌細胞、人結直腸癌LoVo細胞、阿霉素耐藥的M17乳腺癌細胞系和DMS 273肺癌細胞系均表現出有效的抗癌活性[34]。化合物Curacin A首先由Gerwick等從鞘絲藻屬巨大鞘絲藻中分離出來[76],該化合物可導致真核細胞的有絲分裂停滯[77],未來有成為抗癌藥物的潛力。從束藻屬中分離出來的海兔毒素-10是有效的抗細胞增殖劑[78-79],海兔毒素-10的類似物TZT-1027在小鼠癌癥模型中可以有效抑制癌細胞的增殖[80]。目前,海兔毒素類似物ILX-651已成功完成I期臨床試驗[75]。Apratoxin A是從鞘絲藻屬中分離出的環肽,已發現其可誘導人癌細胞的細胞周期停滯和凋亡[81];Somocystinamide A是從海洋藍藻鞘絲藻屬巨大鞘絲藻中分離出來的化合物,可以破壞內皮細胞增殖和內皮細胞小管形成,導致真核細胞的有絲分裂停滯,抑制癌細胞增殖[77]。
研究發現,通過抑制蛋白酶的活性可抑制人癌細胞的生成并誘導細胞凋亡,且對正常細胞的毒性較小[82],因此多種蛋白酶(如胰蛋白酶-3)被認為是癌癥治療的靶點[83],蛋白酶抑制劑在抗癌領域有良好前景。已經有人發現了一些藍藻來源的蛋白酶抑制劑,如圓柱菌素、銅氯菌素[84-85]。來自圓柱菌素的柱胞藻毒素(Cylindrospermopsin,CYN)是由許多淡水藍藻產生的一種聚酮類化合物。CYN可以誘導DNA聚合物形成、DNA鏈斷裂,引起紡錘體破壞,導致染色體非整倍性丟失,進而抑制細胞有絲分裂,抑制癌細胞增殖[85]。胰蛋白酶-3可以介導前列腺癌細胞的增殖和轉移[86],來自銅氯菌素的Suomilide可以抑制胰蛋白酶-3的產生,進而抑制前列腺癌細胞的增殖,同時不影響正常前列腺細胞的增殖[87],可以作為前列腺癌治療的候選藥物進行進一步的開發。
肥胖被認為是一種低度炎癥狀態[88]。鈍頂螺旋藻中的藻藍蛋白和β-胡蘿卜素是有效的抗炎和抗氧化成分,可以通過清除自由基和活性氧來抑制促炎細胞因子(IL-2、TNF-α)的釋放,進而降低血脂、減少脂肪因子的生成,從而有效改善肥胖或超重[89-90]。目前藍藻中的螺旋藻已被開發為膳食補充劑(如螺旋藻片)用于改善肥胖[91-92],未來可能有更多從藍藻中開發的改善肥胖的藥物走向市場。
5 小結與展望
藍藻是天然活性化合物的“合成工廠”和資源寶庫,從藍藻中已發現了豐富的抗菌、殺蟲、抗病毒、抗癌及抗肥胖活性化合物。對藍藻生物活性化合物資源的進一步發掘和研究預期將發現更多的高效、低毒、低成本的新藥物前體,為人類的生命健康作出貢獻。
參 考 文 獻
Singh J S, Kumar A, Rai A N, et al. Cyanobacteria: A precious bio-resource in agriculture, ecosystem, and environmental sustainability[J]. Front Microbiol, 2016, 7: 529.
Lin J Y, Tan S I, Yi Y C, et al. High-level production and extraction of C-phycocyanin from cyanobacteria Synechococcus sp. PCC7002 for antioxidation, antibacterial and lead adsorption[J]. Environ Res, 2022, 206: 112283.
Fay P. Oxygen relations of nitrogen fixation in cyanobacteria[J]. Microbiol Rev, 1992, 56(2): 340-373.
Khalifa S A M, Shedid E S, Saied E M, et al. Cyanobacteria-from the oceans to the potential biotechnological and biomedical applications[J]. Mar Drugs, 2021, 19(5): 241.
Singh R K, Tiwari S P, Rai A K, et al. Cyanobacteria: an emerging source for drug discovery[J]. J Antibiot (Tokyo), 2011, 64(6): 401-412.
Geneva. Antimicrobial resistance fact sheet.[EB/OL]. [2022-5-3] https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance.
World malaria report Geneva: World Health Organization[EB/OL]. [2022-5-3] https://www.who.int/publications/i/item/9789240040496.
Wicht K J, Mok S,Fidock D A. Molecular mechanisms of drug resistance in plasmodium falciparum malaria[J]. Annu Rev Microbiol, 2020, 74: 431-454.
Organization W H. Global tuberculosis report Geneva: World Health Organization [EB/OL]. [2022-5-3] https://apps.who.int/iris/rest/bitstreams/1398397/retrieve.
Jacob S T, Crozier I, Fischer W A, et al. Ebola virus disease[J]. Nat Rev Dis Primers, 2020, 6(1): 13.
Weekly epidemiological update on COVID-19 - 30 November 2022[EB/OL]. [2022-11-30] https://www.who.int/publications/m/item/weekly-epidemiological-update-on-covid-19-30-november-2022.
Haider T, Pandey V, Banjare N, et al. Drug resistance in cancer: mechanisms and tackling strategies[J]. Pharmacol Rep, 2020, 72(5): 1125-1151.
Dixon R A, Al-Nazawi M, Alderson G. Permeabilizing effects of sub-inhibitory concentrations of microcystin on the growth of Escherichia coli[J]. FEMS Microbiol Lett, 2004, 230(2): 167-170.
Lazzaro B P, Zasloff M,Rolff J. Antimicrobial peptides: application informed by evolution[J]. Science, 2020, 368(6490): eaau5480.
Rojas V, Rivas L, Cárdenas C, et al. Cyanobacteria and eukaryotic microalgae as emerging sources of antibacterial peptides[J]. Molecules, 2020, 25(24): 5804.
Zainuddin E N, Jansen R, Nimtz M, et al. Lyngbyazothrins A-D, antimicrobial cyclic undecapeptides from the cultured cyanobacterium Lyngbya sp.[J]. J Nat Prod, 2009, 72(8): 1373-1378.
Xue Y, Zhao P, Quan C, et al. Cyanobacteria-derived peptide antibiotics discovered since 2000[J]. Peptides, 2018, 107: 17-24.
Berry J P, Gantar M, Gawley R E, et al. Pharmacology and toxicology of pahayokolide A, a bioactive metabolite from a freshwater species of Lyngbya isolated from the Florida Everglades[J]. Comp Biochem Physiol C Toxicol Pharmacol, 2004, 139(4): 231-238.
2020年全國細菌耐藥監測報告(簡要版)[EB/OL]. [2021-11-17] http://www.carss.cn/Report/Details/808.
Bui H T, Jansen R, Pham H T, et al. Carbamidocyclophanes A-E, chlorinated paracyclophanes with cytotoxic and antibiotic activity from the vietnamese cyanobacterium Nostoc sp.[J]. J Nat Prod, 2007, 70(4): 499-503.
Kukla D L, Canchola J,Mills J J. Synthesis of the Cyanobacterial Antibiotics Anaephenes A and B[J]. J Nat Prod, 2020, 83(6): 2036-2040.
Cepas V, Gutiérrez-Del-Río I, López Y, et al. Microalgae and cyanobacteria strains as producers of lipids with antibacterial and antibiofilm activity[J]. Mar Drugs, 2021, 19(12): 675.
Schwarzer D, Finking R, Marahiel M A. Nonribosomal peptides: from genes to products[J]. Nat Prod Rep, 2003, 20(3): 275-287.
Karthikeyan C, Tharmalingam N, Varaprasad K, et al. Biocidal and biocompatible hybrid nanomaterials from biomolecule chitosan, alginate and ZnO[J]. Carbohydr Polym, 2021, 274: 118646.
Alavi M. Bacteria and fungi as major bio-sources to fabricate silver nanoparticles with antibacterial activities[J]. Expert Rev Anti Infect Ther, 2022, 20(6): 897-906.
Patel V, Berthold D, Puranik P, et al. Screening of cyanobacteria and microalgae for their ability to synthesize silver nanoparticles with antibacterial activity[J]. Biotechnol Rep (Amst), 2015, 5: 112-119.
Brayner R, Barberousse H, Hemadi M, et al. Cyanobacteria as bioreactors for the synthesis of Au, Ag, Pd, and Pt nanoparticles via an enzyme-mediated route[J]. J Nanosci Nanotechnol, 2007, 7(8): 2696-2708.
El Semary N A,Bakir E M. Multidrug-resistant bacterial pathogens and public Health: The antimicrobial effect of cyanobacterial-biosynthesized silver nanoparticles[J]. Antibiotics (Basel), 2022, 11(8): 1003.
Elkomy R G. Antimicrobial screening of silver nanoparticles synthesized by marine cyanobacterium Phormidium formosum[J]. Iran J Microbiol, 2020, 12(3): 242-248.
Dhillon R H P, Clark J. ESBLs: A Clear and Present Danger?[J]. Critical Care Research and Practice, 2012: 1-11.
Pilmis B,Zahar J R. Ventilator-associated pneumonia related to ESBL-producing gram negative bacilli[J]. Ann Transl Med, 2018, 6(21): 424.
Parveez Ahamed A A, Rasheed M U, Peer Muhamed Noorani K, et al. In vitro antibacterial activity of MGDG-palmitoyl from Oscillatoria acuminata NTAPC05 against extended-spectrum β-lactamase producers[J]. J Antibiot (Tokyo), 2017, 70(6): 754-762.
Prasanna R, Nain L, Tripathi R, et al. Evaluation of fungicidal activity of extracellular filtrates of cyanobacteria--possible role of hydrolytic enzymes[J]. J Basic Microbiol, 2008, 48(3): 186-194.
Patterson G M L, Baldwin C L, Bolis C M, et al. Antineoplastic activity of cultured blue-green algae (cyanophyta)1[J]. J Phycology, 1991, 27(4): 530-536.
Berry J P, Gantar M, Perez M H, et al. Cyanobacterial toxins as allelochemicals with potential applications as algaecides, herbicides and insecticides[J]. Mar Drugs, 2008, 6(2): 117-146.
Macagno A, Molteni M, Rinaldi A, et al. A cyanobacterial LPS antagonist prevents endotoxin shock and blocks sustained TLR4 stimulation required for cytokine expression[J]. J Exp Med, 2006, 203(6): 1481-1492.
Medzhitov R, Preston-Hurlburt P, Janeway C A. A human homologue of the drosophila Toll protein signals activation of adaptive immunity[J]. Nature, 1997, 388(6640): 394-397.
Roy A, Srivastava M, Saqib U, et al. Potential therapeutic targets for inflammation in toll-like receptor 4 (TLR4)-mediated signaling pathways[J]. Int Immunopharmacol, 2016, 40: 79-89.
Molteni M, Bosi A, Rossetti C. The effect of cyanobacterial LPS antagonist (CyP) on cytokines and micro-RNA expression induced by porphyromonas gingivalis LPS[J]. Toxins (Basel), 2018, 10(7): 290.
Tuominen H,Rautava J. Oral microbiota and cancer development[J]. Pathobiology, 2021, 88(2): 116-126.
Villa F A, Lieske K,Gerwick L. Selective MyD88-dependent pathway inhibition by the cyanobacterial natural product malyngamide F acetate[J]. Eur J Pharmacol, 2010, 629(1-3): 140-146.
Kwan J C, Eksioglu E A, Liu C, et al. Grassystatins A-C from marine cyanobacteria, potent cathepsin E inhibitors that reduce antigen presentation[J]. J Med Chem, 2009, 52(18): 5732-5747.
Cunningham C, Appleman L J, Kirvan-Visovatti M, et al. Phase I and pharmacokinetic study of the dolastatin-15 analogue tasidotin (ILX651) administered intravenously on days 1, 3, and 5 every 3 weeks in patients with advanced solid tumors[J]. Clin Cancer Res, 2005, 11(21): 7825-7833.
Jaki B, Orjala J,Sticher O. A novel extracellular diterpenoid with antibacterial activity from the cyanobacterium Nostoc commune[J]. J Nat Prod, 1999, 62(3): 502-503.
Raveh A,Carmeli S. Antimicrobial ambiguines from the cyanobacterium Fischerella sp. collected in Israel[J]. J Nat Prod, 2007, 70(2): 196-201.
Pérez Gutiérrez R M, Martínez Flores A, Vargas Solís R, et al. Two new antibacterial norabietane diterpenoids from cyanobacteria, Microcoleous lacustris[J]. J Nat Med, 2008, 62(3): 328-331.
El-Sheekh M M, Osman M E, Dyab M A, et al. Production and characterization of antimicrobial active substance from the cyanobacterium Nostoc muscorum[J]. Environ Toxicol Pharmacol, 2006, 21(1): 42-50.
Asthana R K, Srivastava A, Singh A P, et al. Identification of an antimicrobial entity from the cyanobacterium Fischerella sp. isolated from bark of Azadirachta indica (Neem) tree[J]. Journal of Applied Phycology, 2006, 18(1): 33-39.
Barbaras D, Kaiser M, Brun R, et al. Potent and selective antiplasmodial activity of the cyanobacterial alkaloid nostocarboline and its dimers[J]. Bioorg Med Chem Lett, 2008, 18(15): 4413-4415.
Clark B R, Engene N, Teasdale M E, et al. Natural products chemistry and taxonomy of the marine cyanobacterium Blennothrix cantharidosmum[J]. J Nat Prod, 2008, 71(9): 1530-1537.
Linington R G, Edwards D J, Shuman C F, et al. Symplocamide A, a potent cytotoxin and chymotrypsin inhibitor from the marine cyanobacterium Symploca sp[J]. J Nat Prod, 2008, 71(1): 22-27.
Sanchez L M, Lopez D, Vesely B A, et al. Almiramides A-C: discovery and development of a new class of leishmaniasis lead compounds[J]. J Med Chem, 2010, 53(10): 4187-4197.
Shao C L, Linington R G, Balunas M J, et al. Bastimolide A, a potent antimalarial polyhydroxy macrolide from the marine cyanobacterium Okeania hirsuta[J]. J Org Chem, 2015, 80(16): 7849-7855.
Despite continued impact of COVID-19, malaria cases and deaths remained stable in 2021[EB/OL]. [2022-12-8] https://www.who.int/news/item/08-12-2022-despite-continued-impact-of-covid-19--malaria-cases-and-deaths-remained-stable-in-2021.
Rao D R, Thangavel C, Kabilan L, et al. Larvicidal properties of the cyanobacterium Westiellopsis sp. (blue-green algae) against mosquito vectors[J]. Trans R Soc Trop Med Hyg, 1999, 93(3): 232.
Becher P G, Keller S, Jung G, et al. Insecticidal activity of 12-epi-hapalindole J isonitrile[J]. Phytochemistry, 2007, 68(19): 2493-2497.
Gantar M, Berry J P, Thomas S, et al. Allelopathic activity among cyanobacteria and microalgae isolated from florida freshwater habitats[J]. FEMS Microbiol Ecol, 2008, 64(1): 55-64.
Gerwick W H, Tan L T,Sitachitta N. Nitrogen-containing metabolites from marine cyanobacteria[J]. Alkaloids Chem Biol, 2001, 57: 75-184.
Simmons T L, Engene N, Ure?a L D, et al. Viridamides A and B, lipodepsipeptides with antiprotozoal activity from the marine cyanobacterium oscillatoria nigro-viridis[J]. J Nat Prod, 2008, 71(9): 1544-1550.
Linington R G, Gonzalez J, Ure?a L D, et al. Venturamides A and B: antimalarial constituents of the panamanian marine cyanobacterium Oscillatoria sp.[J]. J Nat Prod, 2007, 70(3): 397-401.
Mc Phail K L, Correa J, Linington R G, et al. Antimalarial linear lipopeptides from a panamanian strain of the marine cyanobacterium Lyngbya majuscula[J]. J Nat Prod, 2007, 70(6): 984-988.
Wright A D, Papendorf O,K?nig G M. Ambigol C and 2,4-dichlorobenzoic acid, natural products produced by the terrestrial cyanobacterium Fischerella ambigua[J]. J Nat Prod, 2005, 68(3): 459-461.
Kanekiyo K, Lee J B, Hayashi K, et al. Isolation of an antiviral polysaccharide, nostoflan, from a terrestrial cyanobacterium, Nostoc flagelliforme[J]. J Nat Prod, 2005, 68(7): 1037-1041.
Takebe Y, Saucedo C J, Lund G, et al. Antiviral lectins from red and blue-green algae show potent in vitro and in vivo activity against hepatitis C virus[J]. PLoS One, 2013, 8(5): e64449.
Garrison A R, Giomarelli B G, Lear-Rooney C M, et al. The cyanobacterial lectin scytovirin displays potent in vitro and in vivo activity against zaire ebola virus[J]. Antiviral Res, 2014, 112: 1-7.
Boyd M R, Gustafson K R, McMahon J B, et al. Discovery of cyanovirin-N, a novel human immunodeficiency virus-inactivating protein that binds viral surface envelope glycoprotein gp120: potential applications to microbicide development[J]. Antimicrob Agents Chemother, 1997, 41(7): 1521-1530.
Dey B, Lerner D L, Lusso P, et al. Multiple antiviral activities of cyanovirin-N: blocking of human immunodeficiency virus type 1 gp120 interaction with CD4 and coreceptor and inhibition of diverse enveloped viruses[J]. J Virol, 2000, 74(10): 4562-4569.
Klasse P J, Shattock R,Moore J P. Antiretroviral drug-based microbicides to prevent HIV-1 sexual transmission[J]. Annu Rev Med, 2008, 59: 455-471.
Xiong C, OKeefe B R, Byrd R A, et al. Potent anti-HIV activity of scytovirin domain 1 peptide[J]. Peptides, 2006, 27(7): 1668-1675.
Lokhande K B, Apte G R, Shrivastava A, et al. Sensing the interactions between carbohydrate-binding agents and N-linked glycans of SARS-CoV-2 spike glycoprotein using molecular docking and simulation studies[J]. J Biomol Struct Dyn, 2022, 40(9): 3880-3898.
Barre A, Van Damme E J M, Simplicien M, et al. Man-specific lectins from plants, fungi, algae and cyanobacteria, as potential blockers for SARS-CoV, MERS-CoV and SARS-CoV-2 (COVID-19) coronaviruses: biomedical perspectives[J]. Cells, 2021, 10(7): 1619.
Loya S, Reshef V, Mizrachi E, et al. The inhibition of the reverse transcriptase of HIV-1 by the natural sulfoglycolipids from cyanobacteria: contribution of different moieties to their high potency[J]. J Nat Prod, 1998, 61(7): 891-895.
Hayashi T, Hayashi K, Maeda M, et al. Calcium spirulan, an inhibitor of enveloped virus replication, from a blue-green alga Spirulina platensis[J]. J Nat Prod, 1996, 59(1): 83-87.
Hayashi K, Hayashi T,Kojima I. A natural sulfated polysaccharide, calcium spirulan, isolated from Spirulina platensis: in vitro and ex vivo evaluation of anti-herpes simplex virus and anti-human immunodeficiency virus activities[J]. AIDS Res Hum Retroviruses, 1996, 12(15): 1463-1471.
Rechter S, K?nig T, Auerochs S, et al. Antiviral activity of arthrospira-derived spirulan-like substances[J]. Antiviral Res, 2006, 72(3): 197-206.
Nagle D G, Geralds R S, Yoo H D, et al. Absolute configuration of curacin A, a novel antimitotic agent from the tropical marine cyanobacterium Lyngbya majuscula[J]. Tetrahedron Letters, 1995, 36(8): 1189-1192.
Iwasaki S,Shirai R. Natural organic compounds that affect to microtubule functions: syntheses and structure-activity relationships of combretastatins, curacin A and their analogs as the colchicine-site ligands on tubulin[J]. Yakugaku Zasshi, 2000, 120(10): 875-889.
Vaishampayan U, Glode M, Du W, et al. Phase Ⅱ study of dolastatin-10 in patients with hormone-refractory metastatic prostate adenocarcinoma[J]. Clin Cancer Res, 2000, 6(11): 4205-4208.
Luesch H, Moore R E, Paul V J, et al. Isolation of dolastatin 10 from the marine cyanobacterium symploca species VP642 and total stereochemistry and biological evaluation of its analogue symplostatin 1[J]. J Nat Prod, 2001, 64(7): 907-910.
Kobayashi M, Natsume T, Tamaoki S, et al. Antitumor activity of TZT-1027, a novel dolastatin 10 derivative[J]. Jpn J Cancer Res, 1997, 88(3): 316-327.
Luesch H, Chanda S K, Raya R M, et al. A functional genomics approach to the mode of action of apratoxin A[J]. Nat Chem Biol, 2006, 2(3): 158-167.
Daniel K G, Kuhn D J, Kazi A, et al. Anti-angiogenic and anti-tumor properties of proteasome inhibitors[J]. Curr Cancer Drug Targets, 2005, 5(7): 529-541.
Cohen I, Kayode O, Hockla A, et al. Combinatorial protein engineering of proteolytically resistant mesotrypsin inhibitors as candidates for cancer therapy[J]. Biochem J, 2016, 473(10): 1329-1341.
Jaspars M,Lawton L A. Cyanobacteria - a novel source of pharmaceuticals[J]. Curr Opin Drug Discov Devel, 1998, 1(1): 77-84.
Shen X, Lam P K, Shaw G R, et al. Genotoxicity investigation of a cyanobacterial toxin, cylindrospermopsin[J]. Toxicon, 2002, 40(10): 1499-1501.
Diederichs S, Bulk E, Steffen B, et al. S100 family members and trypsinogens are predictors of distant metastasis and survival in early-stage non-small cell lung cancer[J]. Cancer Res, 2004, 64(16): 5564-5569.
Ahmed M N, Wahlsten M, Jokela J, et al. Potent inhibitor of human trypsins from the aeruginosin family of natural products[J]. ACS Chem Biol, 2021, 16(11): 2537-2546.
Bulló M, Casas-Agustench P, Amigó-Correig P, et al. Inflammation, obesity and comorbidities: the role of diet[J]. Public Health Nutrition, 2007, 10(10A): 1164-1172.
Deng R,Chow T J. Hypolipidemic, antioxidant, and antiinflammatory activities of microalgae Spirulina[J]. Cardiovasc Ther, 2010, 28(4): e33-45.
Riss J, Décordé K, Sutra T, et al. Phycobiliprotein C-phycocyanin from spirulina platensis is powerfully responsible for reducing oxidative stress and NADPH oxidase expression induced by an atherogenic diet in hamsters[J]. J Agricultural Food Chemistry, 2007, 55(19): 7962-7967.
Grosshagauer S, Kraemer K,Somoza V. The True Value of Spirulina[J]. J Agric Food Chem, 2020, 68(14): 4109-4115.
Lafarga T, Fernández-Sevilla J M, González-López C, et al. Spirulina for the food and functional food industries[J]. Food Res Int, 2020, 137: 109356.
收稿日期:2022-12-13
作者簡介:田龍龍,本科生,主要從事病原生物學研究。
*通訊作者:李婷婷,博士,研究員,碩士研究生導師,主要從事病原微生物與宿主互作和免疫機制工作。
