2021—2022 年山東榮成海帶產區大規模潰爛災害綜合調查分析
2023-04-18 16:14:54李曉東蘇麗李曉捷李杰徐英江常麗榮于仁成楊德周逄少軍
中國農業科技導報 2023年1期
李曉東 蘇麗 李曉捷 李杰 徐英江 常麗榮 于仁成 楊德周 逄少軍
摘要:2021年11月至2022年4月,海帶(Saccharina japonica)主產區山東榮成發生了海帶潰爛災害,受災面積超9 300 hm2,直接經濟損失20億元。通過現場調查、室內試驗、資料分析等手段進行了研究,結果顯示,2021年10月至2022年12月,由多紋膝溝藻(Gonyaulax polygramma)和紅色赤潮藻(Akashiwo sanguinea)構成的赤潮,順自西向東沿岸流進入榮成海帶養殖區,自北至南蔓延,暴發海域可溶解磷酸鹽含量為3.68 μg·L-1,凸顯磷限制;室內培養試驗顯示問題海域海水正常;浮標觀測顯示,養殖區2021年11月鹽度30.92 PSU,顯著偏低;養殖區海水透明度顯著高于往年同期。綜上,此次災害歸因于海水透明度季節性偏高、赤潮藻枯竭了海帶早期生長賴以需要的營養元素,潰爛海帶促進了褐藻酸降解菌繁殖,加速潰爛過程,帶來次生災害。
關鍵詞:海帶;赤潮;營養鹽;水產養殖;災害doi:10.13304/j.nykjdb.2022.0728
中圖分類號:F326.4 文獻標志碼:A 文章編號:10080864(2023)01020617
海帶(Saccharina japonica)作為重要的養殖經濟海藻,是我國人民菜籃子中的重要水產品。我國人工養殖海帶歷史逾70年,主產區為福建、山東和遼寧3省。山東榮成是海帶養殖的傳統主產區,素有“海帶之鄉”的美譽,每年生產海帶鮮品170 萬t,占全國產量近30%[1]。自榮成北部雞鳴島起,繞過山東半島最東端然后向南到西南端人和鎮都廣泛分布著海帶養殖區,總面積約1 萬hm2[1]。海帶種苗生產、養殖、加工以及依賴海帶為食的鮑、海膽、海參養殖等行業是當地國民經濟發展的重要支柱。榮成地區分布著大小海帶養殖企業72個,中、大型海帶種苗生產企業8個,每年生產海帶種苗59億株[1]。
海帶養殖生產主要包括種苗培育(8 至10月)、苗暫養(10月中旬至11月中旬)和海上養成(11月中旬至次年5、6月)3個階段。種苗培育主要是采取夏苗技術,種海帶來自海上養殖種群或者室內人工蓄養的種群。經過2.5個月室內低溫培育,2~3 cm 的海帶苗在10月10—15日期間出庫下海開始暫養。苗暫養采用水平掛養或者垂養方式,持續1 個月,至11 月中旬,海帶苗達到20 cm左右時開始分苗養殖。分苗養殖是指將單棵海帶苗按照一定間距夾到養殖繩上,然后掛到養殖筏架上進行養殖。夾完苗后的海帶在筏架系統中可以水平或垂直懸掛。目前生產中普遍采用水平懸掛方式。1根養殖繩兩端分別有1根吊繩,一定程度上可以控制水平掛繩深度。水平掛養的海帶實際上是倒著生長,假根部分在上,藻體部分垂直向下。因為海帶假根和生長部相連,這個部分對光照需求不強,過強光照對海帶生長不利,因此需要進行相應水層調節。一般認為,海區溶解無機氮(dissolved inorganic nitrogen, DIN)含量≥0.2 mg·L-1 屬于肥區,含量 0.1~0.2 mg·L-1 屬于中肥區,含量<0.05 mg·L-1 屬于瘦區;海區溶解無機磷(dissolved inorganic phosphorus, DIP) 含量<0.004 mg·L-1 時,海帶生長受到影響;海區氮磷比(N∶P)>16∶1 時,海帶生長主要受活性磷酸鹽限制。海帶筏式養殖以流速大(最大流速25~40 m·min-1)、透明度穩定(養殖周期內變化幅度1~3 m)的海區最為適宜[2]。
2021年11月中下旬,在榮成雞鳴島、榮成灣一帶先后暴發了肉眼可辨的赤潮,海水呈現暗紅色,持續數日,之后分布在赤潮暴發區的剛剛完成夾苗開始生長的海帶出現發黃和潰爛。隨時間推移,這種海帶潰爛現象逐漸從北部榮成灣向南部養殖區擴散,最終蔓延到最南部海帶養殖區人和鎮。
據統計,2022年度榮成市海帶幾乎絕產,預計直接經濟損失近20億元人民幣,同時造成下游加工企業損失超15億元人民幣。造成的間接損失也不容忽視,災情發生后,養殖企業和養殖戶投入的抗災成本合計4.7億元人民幣。由于海帶絕收,當地企業和養殖戶停工,造成勞動力喪失收入預計3.2億元人民幣。此次海帶災害嚴重程度和受災面積均系歷史首次。
災情暴發后,國家藻類產業技術體系組織力量開展了實地調研、海水樣品采集分析、致病微生物分析、病害海帶生化組成分析、室內海帶培養試驗、赤潮藻暴發生物分析、海洋環境數據分析等工作。本文系統地將上述結果進行了整理,以期為防范相關自然災害提供參考。
1 材料與方法
1.1 海帶潰爛形態特征和發展規律調研
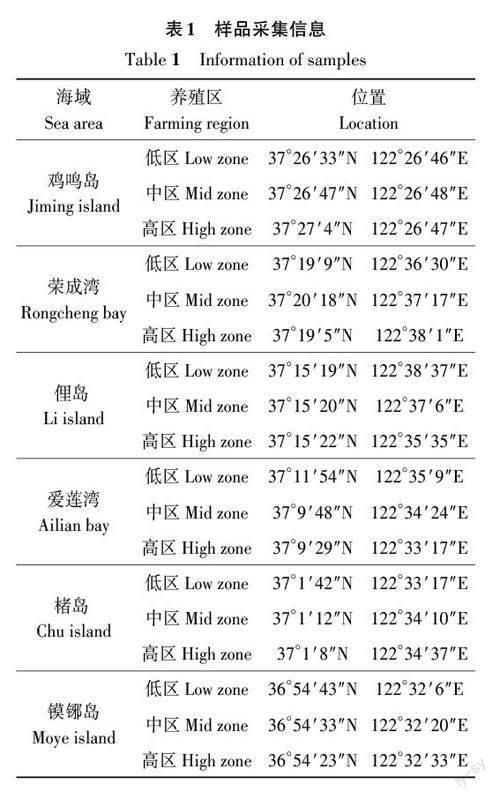
根據榮成近海地理特征及海帶養殖區分布情況,在榮成主要海帶養殖區選擇6個代表性區域,分別對其海帶養殖區低、中、高區(按照離岸距離從近至遠劃分)海帶分苗時間、生長表現、災害發生時間和程度等進行現場調查和連續跟蹤,同時向海帶養殖戶了解其周邊海區災害情況。樣品采集地點見表1。
1.2 海水營養鹽及污染物檢測
1.2.1 養殖海域水樣分析和污染物檢測 調查范圍覆蓋了榮成北部、南部整個養殖海域,共設置了22個站位,每個站位的水樣均采自表層水(水深≤10 m)。檢測指標包括pH、硝酸鹽氮(NO-3-N)、亞硝酸氮(NO-2-N)、氨氮(NH3-N)、溶解無機氮(DIN)、溶解磷酸鹽(DIP)、溶解硅酸鹽(dissolved inorganicsilicon, DISi)、化學需氧量(chemical oxygen demand,COD)、9種重金屬離子、16種三嗪類、酰胺類、二硝基苯胺類除草劑、13種擬除蟲菊酯類殺蟲劑。
各指標的采樣、貯存及后續分析方法依據GB 17378.3—2007《海洋監測規范 第3部分:樣品采集、貯存與運輸》[3]、GB 17378.4—2007《海洋監測規范 第4部分:海水分析》[4]、HJ 700—2014《水質65種元素的測定電感耦合等離子體質譜法》[5]、DB37/T 4013—2020《養殖水體中三嗪類、酰胺類、二硝基甲苯胺類除草劑的測定氣相色譜-質譜法》[6]和張華威等[7]的方法進行測定。
1.2.2 水質標準評價 水質標準判定采用單因子指數(Si)評價法[8],評價標準依據GB 3097—1997《海水水質標準》[ 9],當指數Si ≤1時,表示符合該類水質評價標準,Si越大表示水質污染越嚴重。
1.2.3 海水營養鹽分析 本研究運用相對限制法[10]和絕對限制法[11]評估營養鹽對海帶生長的影響。
1.2.4 海水富營養化程度評價 海水富營養化采用富營養化指數(eutrophication index,E)進行評價,若E>1,表示水體呈現富營養化狀態;若E≤1,表示水體處于貧營養狀態[8]。
1.3 海帶幼苗室內培養
2021年12月23日于榮成愛蓮灣(RC)和青島第二海水浴場(QD)分別采集海水樣品,分別添加1/2 PES[12]作為培養基進行海帶幼苗培育試驗。海帶幼苗來源于中國科學院海藻種質庫(www.caslivealgae.com)室內培育的健康海帶幼苗,長度5 mm,每組20株。培養條件:光照恒溫培養箱,光照/黑暗為12 h/12 h,溫度15 ℃。
1.4 海帶生化組成分析
2022年2月10日采集了雞鳴島(JM)、鏌铘島(MY)受災海帶樣品,2022年2月24日自榮成南部大魚島(DY)采集了尚未發生病變的海帶樣品,2022年2月22日自遼寧省大連市旅順口區采集健康海帶國審品種“中寶1號”(E25)[13]作為對照。每個取樣地點隨機取5棵海帶,烘干后粉碎并混合進行測量,每組測量重復3次。
1.4.1 元素含量測定 樣品中的金屬元素(鐵、鋅、鎂、銅、鈣、磷)檢測方法參照GB 5009.268—2016《食品安全國家標準 食品中多元素的測定》[14],使用電感耦合等離子體質譜法(賽默飛ICAP-RQ)完成測定;碘元素的檢測方法參照GB 5009.267—2016《食品安全國家標準 食品中碘的測定》[15],使用氧化還原滴定法完成測定。
1.4.2 重要糖類含量測定 使用水提取海帶中巖藻糖及褐藻酸,提取液在酸性條件下水解,用1-苯基-3-甲基-5-吡唑啉酮進行衍生,液相色譜紫外檢測器測定;使用水提取海帶中甘露醇,乙醇沉淀去雜后用液相色譜蒸發光散射檢測器進行測定。
1.5 養殖海域赤潮暴發情況分析
1.5.1 赤潮過程的遙感觀測 分析使用的葉綠素數據來自美國國家海洋和大氣管理局(NationalOceanic and Atmospheric Administration, NOAA)極地軌道衛星SNPP(SUOMI national polar-orbitingpartnership)和JPSS-1(joint polar satellite system-1)搭載的可見光與紅外成像輻射儀(visibleinfrared imaging radiometer, VIIRS),從NOAA數據平臺(https://coastwatch.noaa.gov/cw_html/cwViewer.html)獲取葉綠素a數據,分辨率4 km,應用CoastWatch數據分析平臺作圖。
1.5.2 赤潮藻鑒定和分離培養 2021 年12 月11 日在山東半島東北部海域(37.5°~37.8°N、122.4°~122.6°E)采集浮游植物樣品,碘液固定后帶回實驗室,采用Utermühl方法濃縮樣品,在顯微鏡下觀察,對赤潮藻進行鑒定和計數。通過毛細管挑取主要赤潮藻種,室內進行分離培養,獲得純種培養后,通過形態特征進行初步鑒定。同時,收集部分培養藻細胞提取DNA,以28S rRNA 基因D1~D2區序列為靶區,采用1對真核生物通用引物進行擴增[1617],獲得產物后,通過Blast查詢目標藻種,并從GenBank中獲取相關藻種序列信息,對藻種進行輔助鑒定。
1.6 海水鹽度及海表流場模擬分析
海水鹽度和風速風向數據觀測資料來自中國科學院近海海洋觀測研究網絡—— 黃海站16 號浮標觀測系統,浮標站點的經緯度為37°2'43.33″N、122°35'36.28″E,使用的觀測設備為日本ALEC公司生產的ATCW 直讀式溫鹽儀。
觀測時間為2019年11月1—30日和2021年11月1—30日,觀測頻率為1次·h-1。對于觀測得到的原始鹽度數據進行初步處理,剔除因為儀器設備故障導致的異常值以及時間序列(±2) STD以外數據點。對時間序列進行36 h滑動平均,以濾除潮汐信號的主要部分,得到最終數據。
基于Regional Ocean Model System(ROMS)與四維變分同化方法,建成了東中國海海洋動力環境四維變分同化預報系統,給出了2021 年11 月到2022年3月的月平均海表流場;模式配置、驅動場、邊界條件和模擬結果驗證方法參考He等[18]的研究。
1.7 病原微生物調查方法和過程
1.7.1 災區海水菌群和海帶附生菌高通量調查 選擇榮成沿海北、中、南3個區域,分別在雞鳴島(低區和高區)、愛蓮灣(低區、中區和高區)和鏌铘島(低區和高區)海區采集了海水和沉積物樣品,在愛蓮灣(低區、中區和高區)采集潰爛海帶樣品(樣品及代號詳見表2)。海水用0.2 μm孔徑的聚碳酸酯濾膜過濾以收集浮游菌群;沉積物取10 g;海帶樣品切取5 cm× 5 cm表面潰爛部位。水體、沉積物和海帶樣品均按照E.Z.N.A. ? Soil DNAKit (OMEGA, 美國)的要求提取菌群DNA。水體和沉積物菌群DNA用引物對515F 和926R[19] 擴增16S rRNA 的V4~V5 區,海帶菌群DNA 用引物對799F 和1193R[20]擴增16S rRNA基因的V5~V7區。
不同樣品擴增子膠回收目的片段,等量混合后用TruSeq DNA LT sample Prep Kit (Illumina, 美國)建庫,并用Illumina NovaSeq 平臺進行2×250的雙端測序。在2019年1和2月,采用相同的方法對桑溝灣海區的微生物進行測序和分析,相關數據用于本次調查的縱向比較。
下機序列經過拼接、質控后聚類OTU(operational taxonomic unit),計算Alpha 多樣性、Beta多樣性、菌群組成,并對微生物多樣性及群落進行統計分析。
1.7.2 海帶附生菌和海水褐藻酸降解菌數量調查 2021年12月在俚島海區采集潰爛海帶,切取0.5 g潰爛部位,勻漿并稀釋涂布于2216E和褐藻酸培養基。菌落長成后統計單菌落形態特征及菌落數,并進行純化培養。分離得到的菌株用細菌通用引物27F (5-AGAGTTTGATCCTGGCTCAG-3)和1492R (5-TACGGCTACCTTGTTACGACTT-3)對16S rRNA 進行擴增、測序,并在NCBI 網站進行Blast比對確定其分類信息。2021年12月和2022年2月,分別在愛蓮灣養殖海區采集海水,涂布褐藻酸培養基,計數褐藻酸降解菌數量。
2 結果與分析
2.1 海帶潰爛形態特征和發展規律
調查表明,災害最早發生于榮成北部雞鳴島附近海域,于2021年11月底發現分苗后海帶有潰爛發生,12月10日起潰爛加劇,并逐步自西向東、南部海域蔓延。2022年1月10日左右中部愛蓮灣海域海帶開始出現潰爛,隨后向南蔓延至寧津、鏌铘島、石島、人和鎮近岸海帶養殖區。災害發生期間,各區域海帶潰爛癥狀基本相同,主要表現為生長緩慢、藻體發黃、質地脆,起初梢部白爛,藻體表面出現白斑、小泡或腐爛孔洞,隨后白爛部位自梢部逐漸向基部、由葉緣向中帶部蔓延,并伴有假根和柄部色澤加深、生長部變形、葉片邊緣卷曲等現象(圖1A)。總體而言,在單株分苗養殖方式下,同一區域相同時間分苗的不同海帶品種潰爛發生時間、程度無明顯差別;分苗早的海帶(個體相對較大)較分苗晚的海帶(個體較小)抗性強,潰爛程度輕;養殖水層深的海帶較水層淺的海帶潰爛程度輕。南部海域由于災害發生時間較晚,海帶個體較大,受災程度相對輕(圖1C,D)。隨著時間推移,調研區域幾乎所有海帶均發生嚴重潰爛,藻體最終全部脫落,僅余長10~30 cm的基部,甚至只剩假根和柄部殘留在養殖繩上(圖1B)。
2.2 海水營養鹽及污染物檢測結果分析
2.2.1 海水污染物檢測結果分析 檢測了各站位砷、鎘、鋁、鉛、鉻、銅、鋅、汞、硒9種重金屬離子,檢測結果如下:砷為1.50~2.80 μg·L-1,平均值為2.03 μg·L-1;鎘為0.000 52~0.100 00 μg·L-1,平均值為0.051 00 μg·L-1;鋁為1.50~49.54 μg·L-1,平均值為8.27 μg·L-1;硒為0.047~1.660 μg·L-1,平均值為0.620 μg·L-1,其中砷、鎘、硒的Si均小于1,屬于一類水質標準,其他金屬離子均未檢出。
海帶養殖區除草劑檢出3種,分別是阿特拉津(范圍在17.48~30.44 ng·L-1,平均值22.91 ng·L-1)、撲草凈( 范圍在16.90~37.78 ng·L-1,平均值22.14 ng·L-1)、西草凈(范圍在18.56~34.78 ng·L-1,平均值23.45 ng·L-1)。13種擬除蟲菊酯類殺蟲劑均未檢出。
2.2.2 養殖區海水水質標準分析 榮成海帶養殖區海水檢測結果如表3 所示,各站位pH 波動較小,在8.11~8.25 之間,平均值為8.20,Si(pH)≤1,均符合第一類水質標準;DIN 范圍為42.03~675.10 μg·L-1,平均值為192.56 μg·L-1,根據DIN的Si結果,站位4、14的Si(DIN)>1,屬于劣四類海水,站位2、15、16屬于第二類水質,其他站位均屬于一類水質;DIP的范圍在1.33~12.70 μg·L-1,平均值為3.68 μg·L-1,各站位DIP 的Si(DIP)≤1,均符合一類水質標準;檢測站位DISi范圍為86.70~328.00 μg·L-1,平均值為168.27 μg·L-1;檢測站位的COD 范圍在1 154.42~1 571.77 μg·L-1,平均值為1 346.66 μg·L-1,檢測站位COD的Si(COD)≤1,均符合一類水質標準。
2.2.3 養殖區海水營養鹽限制因子分析 海帶養殖區N/P、Si/P、Si/N 見表3,N/P 范圍為15.41~241.97,平均值為62.41;Si/P范圍為0.48~3.64,平均值為1.39;Si/N范圍為10.20~126.82,平均值為60.96。根據相對限制法,檢測的19個站位中(站位4、11、12未檢測硅酸鹽含量),17個站位出現磷限制,均未出現氮限制、硅限制。絕對限制法表明22個站位中有10個站位存在磷限制,未出現氮限制、硅限制。
2.2.4 海水富營養化情況分析 調查區域各站位富營養化指數結果顯示(表3),檢測的16個站位中E 值均小于1,部分站位(站位5、6)小于0.01,表明海帶養殖區處于嚴重貧營養狀態。
2.3 海帶幼苗室內培養
經過4 d 室內培養后,4 組海帶苗均正常生長,海帶苗平均長度分別為(1.29±0.17)(RC)、(1.30± 0.06)(RC+1/2PES)、(1.26±0.13)(QD)、(1.29±0.06)(圖2A);繼續經過4 d培養,RC、RC+1/2PES、QD、QD+1/2PES 海帶苗平均長度分別達到(1.81±0.32)、(3.30±0.41)、(1.66±0.20)、(2.81±0.34) cm,海帶苗均未發生潰爛,說明問題海域海水不存在海帶致死因子(圖2B)。2次測量中,使用RC+1/2PES 作為培養基的海帶幼苗平均長度均為4組中最長;添加營養鹽后海帶苗生長速度加快,顏色更深。
2.4 海帶生化組成
本次試驗采集樣品的生化組成分析結果見圖3。元素分析結果顯示,榮成雞鳴島(JM)、鏌铘島(MY)和大魚島(DY)的海帶樣品在鐵、錳、磷元素的含量上顯著低于旅順地區采集的對照樣品(E25)。巖藻糖、藻酸鹽和甘露醇是海帶重要的光合作用碳代謝產物,液相色譜分析結果顯示,健康海帶與受災海帶樣品中上述3種干物質含量水平接近;MY 的含量顯著高于其他樣品。
2.5 養殖海域赤潮暴發情況分析
2.5.1 赤潮發生過程遙感監測 根據對山東周邊海域葉綠素a含量的遙感分析,自2021年11月上旬起,萊州灣東側海域葉綠素a含量明顯上升。隨著時間推移,葉綠素a 高值區從萊州灣東側逐漸向東擴展,12月上旬煙臺-威海近海葉綠素a含量明顯增加;之后繼續向東擴展,在12月下旬繞過山東半島向南進入榮成近海的海帶養殖區(圖4)。
2.5.2 赤潮藻鑒定 12月11日在威海東北部海域現場調查結果顯示,該海域赤潮優勢種為多紋膝溝藻(Gonyaulax polygramma)和紅色赤潮藻(Akashiwo sanguinea)(圖5),其中,多紋膝溝藻為第一優勢種。現場樣品的計數結果表明,藻細胞密度最高達到3.5×106 cell·L-1,葉綠素a含量最高達到30 μg·L-1,達到赤潮暴發標準。
2.6 海水鹽度及海表流場模擬分析
觀測資料表明, 2021年11月相對于2019年同期的溫度變化不明顯,鹽度存在非常顯著的變化。圖6展示了2019年11月和2021年11月黃海站16號浮標觀測到的2 m深處鹽度變化。通過對比2019和2021年11月平均鹽度可以發現,觀測區域2021年11月的鹽度(平均鹽度30.92 PSU)顯著低于2019年同期(平均鹽度32.35 PSU),表明2021年11月在該區域存在明顯的淡水通量異常,表明該區降水增大或有低鹽水體流入,推測可能與黃河年度輸入淡水增加有關。
通過模擬的海表面流場(圖7)可以看到,海表面流場主要受風場控制,冬季(11 月至次年2月)在北風的控制下,海流主要沿著山東半島岸線以東南向流動為主;進入春季(3月)后,隨著季風轉換,海表流場逐漸轉變方向。
2.7 病原微生物調查結果分析
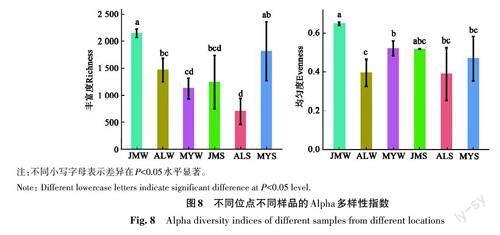
2.7.1 災區海水菌群和海帶附生菌高通量分析 Alpha多樣性結果顯示,水體菌群豐富度呈現自北向南遞減的趨勢,雞鳴島的均勻度最高;從沉積物菌群看,豐富度和均勻度變化趨勢類似,愛蓮灣的指數最低(圖8)。
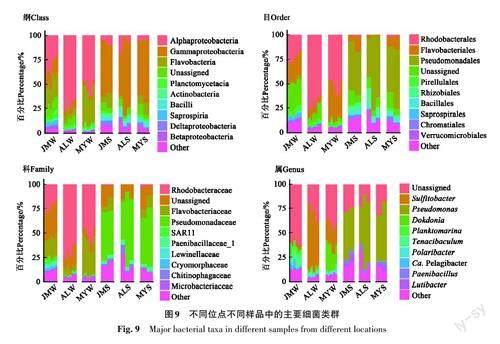
不同海區沉積物有相似細菌組成譜,不同海區水體菌群之間存在較大差別( 圖9)。3個位點水體中主要類群為Alpha變形菌綱(Alphaproteobacteria)(54.69%±18.81%)、黃桿菌綱(Flavobacteriia)(19.74%±12.15%)和Gamma變形菌綱(Gammaproteobacteria)(8.35%±5.33%);在屬水平上,愛蓮灣和鏌铘島有更多的亞硫酸鹽桿菌屬(Sulfitobacter)細菌,其中愛蓮灣最多,鏌铘島次之,雞鳴島最少;鏌铘島還有更多獨島菌屬(Dokdonia)細菌。各海區沉積物在屬水平以假單胞菌屬(Pseudomonas)為主。
對2022 與2019 年桑溝灣菌群豐度進行了比較(圖10)。結果顯示,海洋浮游菌屬(Planktomarina)和Tenacibaculm 屬細菌豐度呈現從北往南遞減趨勢,它們在雞鳴島海區的相對豐度顯著高于其他2個海區及2019年1和2月的相對豐度。假單胞菌屬等褐藻酸降解菌在雞鳴島的相對豐度高于其他2個位點以及2019年2月的相對豐度。愛蓮灣海區亞硫酸鹽桿菌屬和鏌铘島海區獨島菌屬相對豐度顯著高于2019年,假交替單胞菌屬細菌也呈現由北往南遞減趨勢。
2.7.2 海帶附生菌和海水褐藻酸降解菌分析 2216E平板結果顯示,2021年俚島海區海帶潰爛處優勢菌為海神單胞菌屬(Neptunomonas)和假交替單胞菌屬(Pseudoalteromonas)。而海帶表面水泡中的優勢菌為海神單胞菌屬(Neptunomonas),其次為亞硫酸鹽桿菌屬(Sulfitobacter)。2018 年分離的正常海帶上細菌組成相對均勻,存在交替單胞菌屬(Alteromonas)、副球菌屬(Paracoccus)等細菌類群,而潰爛海帶上主要細菌類群則明顯增多,有短桿菌屬(Brevibacterium)、黃桿菌屬(Flavobacterium)、新鞘脂菌屬(Novosphiqobium)、假交替單胞菌屬(Pseudoalteromonas)、假單胞菌屬(Pseudomonas)和希瓦氏菌屬(Shewanella)等(圖11)。
褐藻酸降解菌分析結果顯示,2021年俚島潰爛海帶的優勢菌為噬纖維素菌屬(Cellulophaga),潰爛海帶水泡中未分離到褐藻酸降解菌。2018年潰爛的海帶上以海神單胞菌屬(Neptunomonas)為主,2018 年潰爛的海帶上以假交替單胞菌屬(Pseudoalteromonas)為主,健康海帶表面褐藻酸降解菌以弧菌屬 (Vibrio)為主(圖11)。對海水進行褐藻酸降解菌計數發現,2021年12月愛蓮灣水體中的褐藻酸降解菌數量與2019年2月類似,不超過102·mL-1;在2022年2月,褐藻酸降解菌數量激增,超過104·mL-1。
由圖12 可知,海帶附生菌主要細菌類群有Alpha變形菌綱(Alphaproteobacteria)、Gamma變形菌綱(Gammaproteobacteria) 以及黃小桿菌綱(Flavobacteriia)。將其中主要細菌屬與2019年桑溝灣收集的海帶附生菌群進行比較(圖13),2022年2 月愛蓮灣海帶附生菌中假單胞菌屬(Pseudomonas)、亞硫酸鹽桿菌屬(Sulfitobacter)、極地桿菌屬(Polaribacter)、Tenacibaculm 屬和交替赤細菌屬(Altererythrobacter)等類群相對豐度顯著高于2019年1和2月相對豐度。2019年海帶的優勢類群如青枯菌屬(Ralstonia)、污泥單胞菌屬(Pelomonas)和紅球菌屬(Rhodococcus)等在2022年愛蓮灣海帶中相對豐度較低(<1%)或未檢測到。具有褐藻酸降解能力的細菌顯著高于往年同期水平。
3 討論
3.1 赤潮暴發和海帶潰爛的關系
2021年冬季,山東半島北部海域發生了較大規模甲藻赤潮,這一現象在山東近海非常罕見。通過對2021年9—12月的遙感資料分析,可看出甲藻赤潮在10月份出現在萊州灣東側,11月份赤潮向東擴展至煙臺-威海一帶,12月份繞過山東半島南下進入榮成近海,模擬海表面流場結果與赤潮運動方向一致。對比此次甲藻赤潮的時空分布狀況及海帶病害出現的時空規律可以看出,二者之間高度吻合,赤潮暴發及其對海域環境的改變極有可能是海帶潰爛出現的首要、直接誘導因子。
多紋膝溝藻和紅色赤潮藻是近海常見的2種有害赤潮藻。多紋膝溝藻在我國近海較常見,曾多次形成赤潮。林永水等[21]報告了1991年5月在廣東大鵬灣鹽田附近一次多紋膝溝藻赤潮;潘雪峰等[22]報告了2004年9月在江蘇海州灣連島附近海域一次多紋膝溝藻赤潮。這2次報道均未提及赤潮導致的危害效應,僅對赤潮過程與環境因子相關性進行了分析。紅色赤潮藻在我國近海廣泛分布,煙臺四十里灣曾多次發生紅色赤潮藻赤潮[23],對養殖動物造成了一定損害。在以往報道中均未提及赤潮對大型海藻的不利影響。本團隊的室內培養結果顯示,問題海域海水不存在海帶致死因子,海帶幼苗可以健康存活,另外在問題海域的其他海藻種類比如裙帶菜、蜈蚣藻、龍須菜等都沒有發生類似海帶的潰爛問題,表示這個環境條件的變化只對夾苗養殖的海帶不利。
3.2 海帶生長不良與養殖方式、環境條件異常的關系
海帶正常生長受溫度、光照、營養鹽、pH、鹽度、透明度以及海水流速等因子影響,其中溫度、光照和營養鹽最為重要[24]。海帶適宜生長的水溫在1~13 ℃,災害暴發期間該海域溫度在5 ℃左右,符合海帶生長的條件,但養殖區海水處于嚴重貧營養狀態,養殖區各站位DIN平均含量192.56 μg·L-1,DISi平均含量為168.27 μg·L-1,DIP平均含量僅為3.68 μg·L-1。謝琳萍等[25]研究指出,本區域該季節DIN 含量79.80 μg·L-1,DISi 為188.83 μg·L-1,DIP含量為11.16 μg·L-1,二者對比磷限制問題突出。污染物阿特拉津、撲草凈、西草凈濃度較低,生態風險可以忽略[26]。殺蟲劑均未檢出,重金屬離子符合一類水質標準。海水透明度測定顯示,2022年1和2月楮島海區(泉海水產公司養殖區中區)海水透明度分別為1.3 和2.2 m,顯著高于往年同期(0.5~0.8 m),其他區域海水透明度也明顯高于往年,部分海區甚至高達4 m。2021年11月前后觀測區域明顯的淡水通量異常導致了海水鹽度顯著降低,平均鹽度僅為30.92 PSU。綜合推斷,長時間海水營養貧瘠、高透明度海水引發海帶潰爛的發生。
研究表明,強烈的太陽光(可見光和紫外線)可以引起海帶卷曲和潰爛[27],這也是我國在海帶養殖早期提出養殖過程需要調整水層的根本原因[28]。海帶養殖是倒立進行,單棵海帶的假根固著在養殖繩上,生長部處于整個藻體最頂端,是直接接受太陽光照射的部位。如果海水透明度過高,過強的可見光和紫外線都能夠給海帶生長部帶來直接傷害,使海帶出現卷曲、色素丟失、藻體潰爛等癥狀。在海帶生長期內,具有過高海水透明度的海域通常不適合進行海帶養殖生產,除非將海帶筏架沉入到安全的海水深度。2021—2022年大魚島2月份現場的觀察發現,簇養的海帶和簇養的裙帶菜生長基本正常,雖然也出現了一些由于營養鹽不足帶來的藻體發黃、發脆,但是藻體基本完整,而周圍的單棵夾苗養殖的海帶品種,基本已經潰爛流失。簇養的海帶一般有6~10棵,其生長部可以相互遮光而獲得保護,從而避免了太陽光的直接照射,很大程度上躲過了強光的危害。
海水中磷限制通常被認為是由于浮游植物集中繁殖、營養鹽被大量消耗導致,另外海水中懸浮顆粒物質對其也有吸附作用[10,29]。調查中發現,養殖區海水透明度較高、懸浮顆粒含量極低,因此懸浮顆粒對其吸附可能性不高,基本可以確定營養鹽的問題由赤潮引起。喻龍等[30]報道,紅色赤潮藻赤潮暴發后海水中各類營養鹽含量降低,最終出現磷限制,導致赤潮的最終消退。陳月紅等[31]調查發現,甲藻赤潮暴發后海水中各營養鹽大幅下降,比例失衡,導致赤潮最終消散,這些均證明赤潮導致養殖區營養鹽失衡,出現磷限制,影響了海帶生長,繼而發生潰爛。榮成地區海帶中磷元素含量均小于1 000 mg·kg-1,顯著低于健康海帶(平均1 959 mg·kg-1),進一步佐證了這個看法。
綜上,2021年11月至2022年2月,受災海區經歷了高透明度、低營養鹽、低鹽度等異常海水條件,引發海帶發黃、卷曲和潰爛。
3.3 海帶潰爛和微生物菌群變化的關系
浮游菌群Alpha多樣性指數顯示,雞鳴島水體中的菌群多樣性指數在3 個海區最高。研究表明,赤潮會導致養殖環境中浮游菌群Alpha多樣性指數顯著升高[32]。浮游菌群組成分析結果也反映出赤潮對養殖海區的影響。Planktomarina 和Ca.pelagibacter 是水體出現赤潮時的重要標志類群[3334]。災害發生后假單胞菌屬(Pseudomonas)和Tenacibaculm 屬相對豐度同樣呈現自北向南遞減趨勢且顯著高于它們在2019年同期水平。假單胞菌(Pseudomonas)是CAZY數據庫中已經記錄的褐藻酸降解菌,Tenacibaculm 屬細菌也是赤潮暴發時海水中重要細菌類群,具有瓊脂降解活性及溶藻活性[3536]。因此,二者都是海帶潛在條件致病菌。Sulfitobacter 屬細菌是維持赤潮海區浮游菌群的結構和功能的關鍵類群[37],本研究中該類群在雞鳴島海區相對豐度顯著低于其余2個海區。
分別利用培養和高通量分析方法對潰爛海帶進行了分析,并與正常年份健康海帶樣品菌群組成進行了對比。結果顯示,2018 年(正常年份)2216E和褐藻酸培養基培養的2018年潰爛海帶上的細菌種類明顯多于健康海帶,而2021年俚島海區潰爛海帶雖然在細菌種類上與2018年健康海帶沒有明顯差別,但顯示出明顯不同的群落結構,且存在單一優勢細菌類群,潰爛處和水泡處的海神單胞菌屬(Neptunomonas)(2216E平板)及潰爛處的噬纖維素菌屬(Cellulophaga)。海神單胞菌屬細菌具有有機物降解活性[3839],噬纖維素菌屬細菌除具有較強褐藻酸降解活性外[40],還具有較強的卡拉膠和纖維素降解活性[4142],推測這些細菌可能對海帶形成嚴重威脅,導致富含褐藻酸鈉的海帶發生潰爛。高通量測序結果支持上述推測。通過與2019年數據對比,發現愛蓮灣潰爛海帶上存在的主要細菌類群豐度均顯著高于2019年的健康海帶(除Hella 屬)。研究發現,這些細菌多具有藻類降解活性,具有成為潛在病原的可能。例如假單胞菌(Pseudomonas)具有褐藻酸降解活性,亞硫酸桿菌(Sulfitobacter)具有多糖降解活性[43];Tenacibaculm、極地桿菌(Polaribacter)和交替赤細菌屬(Altererythrobacter)等具有瓊脂、多糖降解及溶藻活性[3536,4445]。各種降解活性細菌類群及其豐度的顯著增加成為養殖海帶潰爛次生原因,海水中的褐藻酸降解菌數量激增也證實了這一推論。
已有研究表明,海藻病害多是環境、微生物以及宿主復雜相互作用結果[4647],環境變化往往起誘導作用。養殖環境異于正常生長環境的顯著變化(例如高溫、高光、缺乏營養、污染物等)使海藻處于環境脅迫條件,導致其生理狀態下降,防御水平降低,海藻附生菌群間接發生顯著變化,條件致病菌大量繁殖,最終促成病害發生。根據這種推斷,認為榮成2021—2022年海帶潰爛過程中,赤潮是導致環境發生顯著變化的直接原因,增加了環境中潛在病原數量,降低了水中營養鹽水平,加之海水透明度過高等環境變化使海帶防御水平降低,附生菌群向著易于發生潰爛的方向轉變,導致潰爛發生。而降解類細菌類群又隨著潰爛、脫落的海帶以及赤潮洋流進行繁殖和傳播,最終導致海區菌群持續失衡,致使海帶在更大的范圍上持續潰爛。
參 考 文 獻
[1] 農業農村部漁業漁政管理局, 全國水產技術推廣總站, 中國
水產學會. 2021中國漁業統計年鑒[M]. 北京: 中國農業出
版社, 2021:1-158.
Fishery Bureau of Ministry of Agriculture and Rural Affairs of
China, National Fisheries Technology Extension Center China
Society of Fisheries. China Fishery Statistical Yearbook [M].
Beijing: China Agriculture Press, 2021:1-158.
[2] 索如瑛. 海帶養殖[M]. 北京: 中國農業出版社, 1988:35-36.
SUO R Y. Kelp Farming [M]. Beijing: China Agriculture Press,
1988:35-36.
[3] 徐恒振,馬永安,于濤,等. 海洋監測規范 第3部分:樣品采
集、貯存與運輸:GB 17378.3—2007[S]. 北京: 中國標準出
版社, 2007.
[4] 馬永安,徐恒振,于濤,等. 海洋監測規范 第4部分:海水分析:
GB 17378.4—2007[S]. 北京: 中國標準出版社, 2007.
[5] 江蘇省環境監測中心,蘇州市環境監測中心站. 水質65種
元素的測定 電感耦合等離子體質譜法:HJ 700—2014[S]. 北
京:中國環境出版社, 2014.
[6] 張秀珍,徐英江,張華威,等. 養殖水體中三嗪類、酰胺類、
二硝基甲苯胺類除草劑的測定 氣相色譜-質譜法:DB37/T
4013—2020[S]. 山東: 山東省市場監督管理局, 2020.
[7] 張華威, 崔艷梅, 王倩, 等. 氣相色譜法同時測定海水中13
種擬除蟲菊酯類殺蟲劑的殘留量[J]. 中國漁業質量與標準,
2019, 9(3): 45-52.
ZHANG H W, CUI Y M, WANG Q, et al .. Determination of 13
kinds of pyrethroid insecticides residues in seawater using gas
chromatography method [J]. Chin. Fish. Quality Standards,
2019, 9(3): 45-52.
[8] 劉愛英, 馬元慶, 李斌, 等. 山東三個貝類增養殖區營養鹽
季節分布及營養鹽限制性分析[J]. 海洋環境科學, 2013,
32(1): 54-57.
LIU A Y, MA Y Q, LI B, et al .. Seasonal distribution of
nutrient and nutrient limitation in shellfish culture area of
Shandong [J]. Marine Environ. Sci., 2013, 32(1): 54-57.
[9] 黃自強,張克,許昆燦,等. 海水水質標準:GB 3097—1997[S].
北京: 中國標準出版社, 1997.
[10] 張鍺, 任宏偉, 牟亮, 等. 春季青島鰲山灣沿岸海水水質狀
況分析與評價[J]. 海洋環境科學, 2021, 40(4): 515-520.
ZHANG Z, REN H W, MOU L, et al .. Assessment on the water
quality of coastal seawater in the oshan bay, Qingdao in spring [J].
Marine Environ. Sci., 2021, 40(4): 515-520.
[11] JUSTI? D, RABALAIS N N, TURNER R E. Stoichiometric
nutrient balance and origin of coastal eutrophication [J].
Marine Poll. Bull., 1995, 30(1): 41-46.
[12] PROVASOLI L. Media and prospects for the cultivation of
marine algae [C]// Cultures and Collections of Algae. Proceedings
of US-Japan Conference, Hakone, September 1966. Japan
Society of Plant Physiology, 1968: 63-15.
[13] SU L, SHAN T F, LI J, et al .. Characterization of the novel
hybrid cultivar E25 of Saccharina japonica in the northern
farming region of China [J]. J. Appl. Phycol., 2021, 33: 4011-
4021.
[14] 中華人民共和國國家衛生和計劃生育委員會,國家食品藥
品監督管理總局. 食品安全國家標準 食品中多元素的測
定:GB 5009.268—2016[S]. 北京: 中國標準出版社, 2016.
[15] 中華人民共和國國家衛生和計劃生育委員會,國家食品藥
品監督管理總局. 食品安全國家標準 食品中碘的測定:GB
5009.267—2016[S]. 北京: 中國標準出版社, 2016.
[16] LENAERS G, MAROTEAUX L, MICHOT B, et al ..
Dinoflagellates in evolution. a molecular phylogenetic analysis
of large subunit ribosomal RNA [J]. J. Mol. Evol., 1989, 29(1):
40-51.
[17] YU R C, ZHANG Q C, LIU Y, et al .. The dinoflagellate
Alexandrium catenella producing only carbamate toxins may
account for the seafood poisonings in Qinhuangdao, China [J/OL].
Harmful Algae, 2021, 103: 101980 [2022-07-30]. https://doi.
org/10.1016/j.hal.2021.101980.
[18] HE Z, YANG D, WANG Y, et al .. Impact of 4D-Var data
assimilation on modelling of the East China Sea dynamics [J/OL].
Ocean Model (Oxf), 2022, 176: 102044 [2022-07-30].https://
doi.org/10.1016/j.ocemod.2022.102044.
[19] WALTERS W, HYDE E R, BERG-LYONS D, et al .. Improved
bacterial 16S rRNA gene (V4 and V4-5) and fungal internal
transcribed spacer marker gene primers for microbial
community surveys [J/OL]. mSystems, 2016, 1(1): e00009-15
[2022-07-30].https://doi.org/10.1128/mSystems.00009-15.
[20] ZHANG J, ZHANG N, LIU Y X, et al .. Root microbiota shift in
rice correlates with resident time in the field and developmental
stage [J]. Sci. China Life Sci., 2018, 61(6): 613-621.
[21] 林永水, 周近明, 黎愛韶. 鹽田水域多紋膝溝藻赤潮發生過
程的生態學研究[J]. 熱帶海洋, 1993(1): 46-50,107.
LIN Y S, ZHOU J M, LI A S. An ecological study on the
occurrence of the tide of Gonyaulax Polygramma in Yantian
waters, Dapeng Bay [J]. J. Phys. Oceanogr., 1993(1): 46-50,107.
[22] 潘雪峰, 張鷹, 劉吉堂. 海州灣多紋膝溝藻赤潮的動態相關
分析[J]. 海洋環境科學, 2007,26(6): 523-526.
PAN X F, ZHANG Y, LIU J T. Dynamic and correlative
analysis of the Gonyaulax Polygramma red tide in Haizhou
Gulf [J]. Mar. Environ. Res., 2007,26(6): 523-526.
[23] 周健, 王瑋, 吳志宏, 等. 山東沿海赤潮災害基本特征及防
控對策建議[J]. 海洋環境科學, 2020, 39(4): 537-543.
ZHOU J, WANG W, WU Z H, et al .. The basic characteristics
and prevention countermeasures of red tide in Shandong coast
waters [J]. Mar. Environ. Res., 2020, 39(4): 537-543.
[24] 蔡霞. 大型藻類對氮的吸收利用模型研究[D]. 舟山:浙江海
洋大學, 2018.
CAI X. Modelling nitrogen absorption and utilization of
Laminaria japonica [D]. Zhoushan:Zhejiang Ocean University,
2018.
[25] 謝琳萍, 蒲新明, 孫霞, 等. 榮成灣營養鹽的時空分布特征
及其影響因素分析[J]. 海洋通報, 2013, 32(1): 19-27.
XIE L P, PU X M, SUN X, et al .. Analysis on the temporal and
spatial distribution of nutrients and the influence factors in
Rongcheng Bay [J]. Marin. Sci. Bull., 2013, 32(1): 19-27.
[26] 徐英江, 劉慧慧, 任傳博, 等. 萊州灣海域表層海水中三嗪
類除草劑的分布特征[J]. 漁業科學進展, 2014, 35(3): 34-39.
XU Y J, LIU H H, REN C B, et al .. Distributions of the tiazine
hrbicides in the srface sawater of Laizhou Bay [J]. Prog. Fish.
Sci., 2014, 35(3): 34-39.
[27] 何培民, 張澤宇, 張學成, 等. 海藻栽培學[M]. 北京: 科學出
版社, 2018:46-49.
HE P M, ZHANG Z Y, ZHANG X C, et al .. Seaweed
Cultivation [M]. Beijing: Science Press, 2018:46-49.
[28] 曾呈奎, 吳超元. 海帶養殖學[M]. 北京: 科學出版社, 1962:
173-174.
ZENG C K, WU C Y. The Kelp Cultivation [M]. Beijing:
Science Press, 1962:173-174.
[29] 劉敏, 侯立軍, 許世遠, 等. 長江河口潮灘表層沉積物對磷
酸鹽的吸附特征[J]. 地理學報, 2002,57(4): 397-406.
LIU M, HOU L J, XU S Y, et al .. Phosphate adsorption
characteristics of tidal flat surface sediments and its
environmental effect from the Yangtz estuary [J]. Acta. Geogr.
Sin., 2002,57(4): 397-406.
[30] 喻龍, 馬兆虎, 郝彥菊, 等. 四十里灣2012年一次血紅哈卡
藻赤潮的調查分析[J]. 海洋科學進展, 2014, 32(1): 84-91.
YU L, MA Z H, HAO Y J, et al .. Analysis of the red tide
caused by Akashiwo sanguinea in Sishili Bay, 2012 [J]. Adv.
Marin. Biol., 2014, 32(1): 84-91.
[31] 陳月紅, 石曉勇, 韓秀榮, 等. 閩浙近岸海域赤潮暴發前后
海水營養鹽特征分析[J]. 海洋通報, 2015, 34(5): 568-576.
CHEN Y H, SHI X Y, HAN X R, et al .. Characteristics of
seawater nutrients during the occurrence of harmful algal
blooms in Fujian and Zhejiang coastal areas [J]. Marin. Sci.
Bull., 2015, 34(5): 568-576.
[32] SUN F, WANG C, WANG Y, et al .. Diatom red tide
significantly drive the changes of microbiome in mariculture
ecosystem [J/OL]. Aquaculture, 2020, 520: 734742[2022-07-
30]. https://doi.org/10.1016/j.aquaculture.2019.734742.
[33] TEELING H, FUCHS B M, BECHER D, et al .. Substratecontrolled
succession of marine bacterioplankton populations
induced by a phytoplankton bloom [J]. Science, 2012,
336(6081): 608-611.
[34] VOGET S, WEMHEUER B, BRINKHOFF T, et al .. Adaptation
of an abundant roseobacter RCA organism to pelagic systems
revealed by genomic and transcriptomic analyses [J]. Isme. J.,
2015, 9(2): 371-384.
[35] LI Y, WEI J, YANG C, et al .. Tenacibaculum xiamenense sp.
nov., an algicidal bacterium isolated from coastal seawater [J].
Int. J. Syst. Evol. Micr., 2013, 63( 9): 3481-3486.
[36] XU Z X, YU P, MU D S, et al .. Tenacibaculum agarivorans sp.
nov., an agar-degrading bacterium isolated from marine alga
Porphyra yezoensis Ueda [J]. Int. J. Syst. Evol. Micr., 2017,
67(12): 5139-5143.
[37] XU S, HE C, SONG S, et al .. Spatiotemporal dynamics of
marine microbial communities following a Phaeocystis bloom:
biogeography and co-occurrence patterns [J]. Environ.
Microbiol. Rep., 2021, 13(3): 294-308.
[38] PU N, WANG M R, LI Z J. Characterization of
polyhydroxyalkanoate synthases from the marine bacterium
Neptunomonas concharum JCM17730 [J]. J. Biotechnol., 2020,
319: 69-73.
[39] DI?GUEZ A L, PICHON P, BALBOA S, et al .. Complete
characterization of new isolates of Neptunomonas phycophila
leads to emend its description and opens possibilities of
biotechnological applications [J/OL]. MicrobiologyOpen, 2017,
6(6): e00519[2022-07-30]. https://doi.org/10.1002/mbo3.519.
[40] MAHAJAN S, RAMYA T N C. Cellulophaga algicola alginate
lyase inhibits biofilm formation of a clinical Pseudomonas
aeruginosa strain MCC 2081 [J]. IUBMB Life, 2021, 73(2):
444-462.
[41] HOWLADER M M, MOLZ J, SACHSE N, et al ..
Optimization of fermentation conditions for carrageenase
production by cellulophaga species: a comparative study
[J/OL]. Biology, 2021, 10(10): 971[2022-07-30]. https://doi.
org/10.3390/biology10100971.
[42] WANG Y, YU W, HAN F. Expression and characterization of a
cold-adapted, thermotolerant and denaturant-stable GH5
endoglucanase Celal_2753 that withstands boiling from the
psychrophilic bacterium Cellulophaga algicola IC166T [J].
Biotechnol. Letters, 2016, 38(2): 285-290.
[43] DOGS M, WEMHEUER B, WOLTER L, et al ..
Rhodobacteraceae on the marine brown alga Fucus spiralis are
abundant and show physiological adaptation to an epiphytic
lifestyle [J]. Syst. Appl. Microbiol., 2017, 40(6): 370-382.
[44] FERN?NDEZ-G?MEZ B, RICHTER M, SCH?LER M, et al ..
Ecology of marine Bacteroidetes: a comparative genomics
approach [J]. Isme. J., 2013, 7(5): 1026-1037.
[45] LEI X, LI Y, CHEN Z, et al .. Altererythrobacter xiamenensis sp.
nov., an algicidal bacterium isolated from red tide seawater [J].
Int. J. Syst. Evol. Micr., 2014, 64( 2): 631-637.
[46] EGAN S, GARDINER M. Microbial dysbiosis: rethinking
disease in marine ecosystems [J/OL]. Front. Microbiol., 2016,
7:991[2022-07-30].https://doi.org/10.3389/fmicb.2016.00991.
[47] EGAN S, FERNANDES N D, KUMAR V, et al .. Bacterial
pathogens, virulence mechanism and host defence in marine
macroalgae [J]. Environ. Microbiol., 2014, 16(4): 925-938.
(責任編輯:胡立霞)
