基于分子與形態證據的傣藥“傣百解”基原考證
2023-03-13 07:47:14李金月譚運洪李海濤巖罕單龔燕雄肖云學郁文彬
廣西植物 2023年1期
李金月, 譚運洪, 李海濤, 巖罕單, 龔燕雄, 肖云學, 郁文彬*
( 1. 中國科學院西雙版納熱帶植物園 綜合保護中心, 云南 勐臘 666303; 2. 中國科學院大學, 北京 100049; 3. 中國科學院東南亞生物多樣性研究中心, 云南 勐臘 666303; 4. 中國醫學科學院、北京協和醫學院藥用植物研究所云南分所, 云南 景洪 666100; 5. 西雙版納州傣醫醫院, 云南 景洪 666100; 6. 云南省熱帶作物科學研究所, 云南 景洪 666100; 7. 中國科學院西雙版納熱帶植物園 園林園藝部, 云南 勐臘 666303 )
“傣百解”為國藥準字品種“雅解片”,傣族稱為“雅解先打”,也俗稱“大百解”,意譯為“解百毒的藥”(云南省食品藥品監督管理局, 2005)。“解藥”是傣族醫藥體系中最為重要的組成部分,也是區別于其他民族藥的特色藥物。“傣百解”是傣醫常用的解藥之一,在中國云南省傣族聚居的西雙版納州、德宏州、新平縣、元江縣等地,以及老撾、緬甸等國家廣泛使用(中國科學院中國植物志編輯委員會,1977)。“傣百解”可用于清熱解毒、止咳平喘,具有抗腫瘤、降血壓、抗炎等功效,還常用于治療藥物或食物中毒等(國家中醫藥管理局, 2005; Wang et al., 2018)。根據已有的文獻報道,從“傣百解”基原植物通光散(Marsdeniatenacissima)中已分離出190余種化學成分,主要包括甾體類化合物、三萜類化合物、有機酸化合物和生物堿化合物等(廖矛川等, 2016),其中固醇類化合物是抗腫瘤和耐化療逆轉作用的特有物質和主要活性成分(Wang et al., 2018)。
“傣百解”用藥歷史悠久,現已被西雙版納州傣醫院用作多個院內制劑的組方藥材,如百解膠囊、雅解嘎罕、版納涼劑等。其中,百解膠囊在傣醫臨床應用最為廣泛,具有良好的臨床療效和開發應用前景(楊蓮等, 2021)。由于“傣百解”具有重要藥用價值、市場前景良好、開發潛力大,因此需要加強對“傣百解”基原植物野生藥材資源的保護,使該藥用植物資源得到可持續利用。研究和開發利用傣藥的首要環節就是對傣藥藥材品種進行整理與考證,這對澄清基原植物和確保傣藥藥材質量以及用藥安全具有重要意義(段寶忠等, 2015)。
基原鑒定是藥材鑒定的核心內容之一。自然條件的限制以及醫著記載不詳等原因,導致各地的傣醫在一些藥材基原的認識上存在分歧,其中對“傣百解”也存在基原不清和錯誤鑒定的問題。《云南省中藥材標準》(2005年版)記載了“傣百解”基原是蘿藦科(Asclepiadaceae)的苦繩(Dregeasinensis)的根(云南省食品藥品監督管理局, 2005)。管艷紅等(2012)采用石蠟切片對“傣百解”藥材和苦繩的根進行橫切面結構比較發現兩者存在明顯差異,認為“傣百解”和苦繩是兩種不同的植物。同年,管志斌等(2012)利用ISSR分子標記分析“傣百解”和幾種牛奶菜屬(Marsdenia)藥用植物的親緣關系,發現“傣百解”與通光散的親緣關系最近。隨后,李海濤(2014)等通過綜合文獻查證、形態學解剖、生藥學等證據對“傣百解”的基原進行了考證,發現“傣百解”基原植物為苦繩是早期的錯誤鑒定,認為“傣百解”的基原植物應該是蘿藦科牛奶菜屬的通光散。值得注意的是,在最新的被子植物分類系統中已將蘿藦科歸并到夾竹桃科(Apocynaceae)作為1個亞科,即蘿藦亞科(Asclepiadoideae) (The Angiosperm Phylogeny Group, 2016)。
目前,由于“傣百解”藥材的研究主要在形態學、生藥學和化學成分等方面且研究比較零散(管艷紅等,2012;李海濤等,2014;廖矛川等,2016),所以整合形態特征與分子證據以開展“傣百解”基原植物考證具有重要意義。由于傳統的藥材鑒定方法具有一定的局限性,尤其是近緣種、易混種在沒有花和果等關鍵特征的情況下分類鑒定難度比較大,因此容易出現錯誤鑒定。隨著DNA測序技術的快速發展,分子鑒定方法因具有方便、快捷和準確等特點而被廣泛應用于物種和藥材鑒定(Yu et al., 2021)。本研究以澄清傣藥“傣百解”基原植物為目的,利用栽培的“傣百解”基原植物材料、通光散以及牛奶菜族的其他植物樣品,采用形態結合分子的方法,通過整合形態性狀比較和系統發育樹分析,探討以下問題:(1) “傣百解”基原植物是否為通光散;(2) “傣百解”的近緣種有哪些;(3) 如何鑒別區分“傣百解”及其近緣種。
1 材料與方法
1.1 材料
新收集了夾竹桃科牛奶菜屬5種共17份樣品,包括栽培于中國醫學科學院藥用植物所云南分所南藥園“傣百解”樣品4份,以及南山藤屬(Dregea) 2種7份樣品和匙羹藤屬(Gymnema)1種1份樣品。具體樣品來源和憑證信息如表1所示。采集植物樣品的新鮮葉片經變色硅膠進行快速干燥,并低溫保存,以備實驗所需。

表 1 研究材料及其憑證信息Table 1 Study materials and voucher information
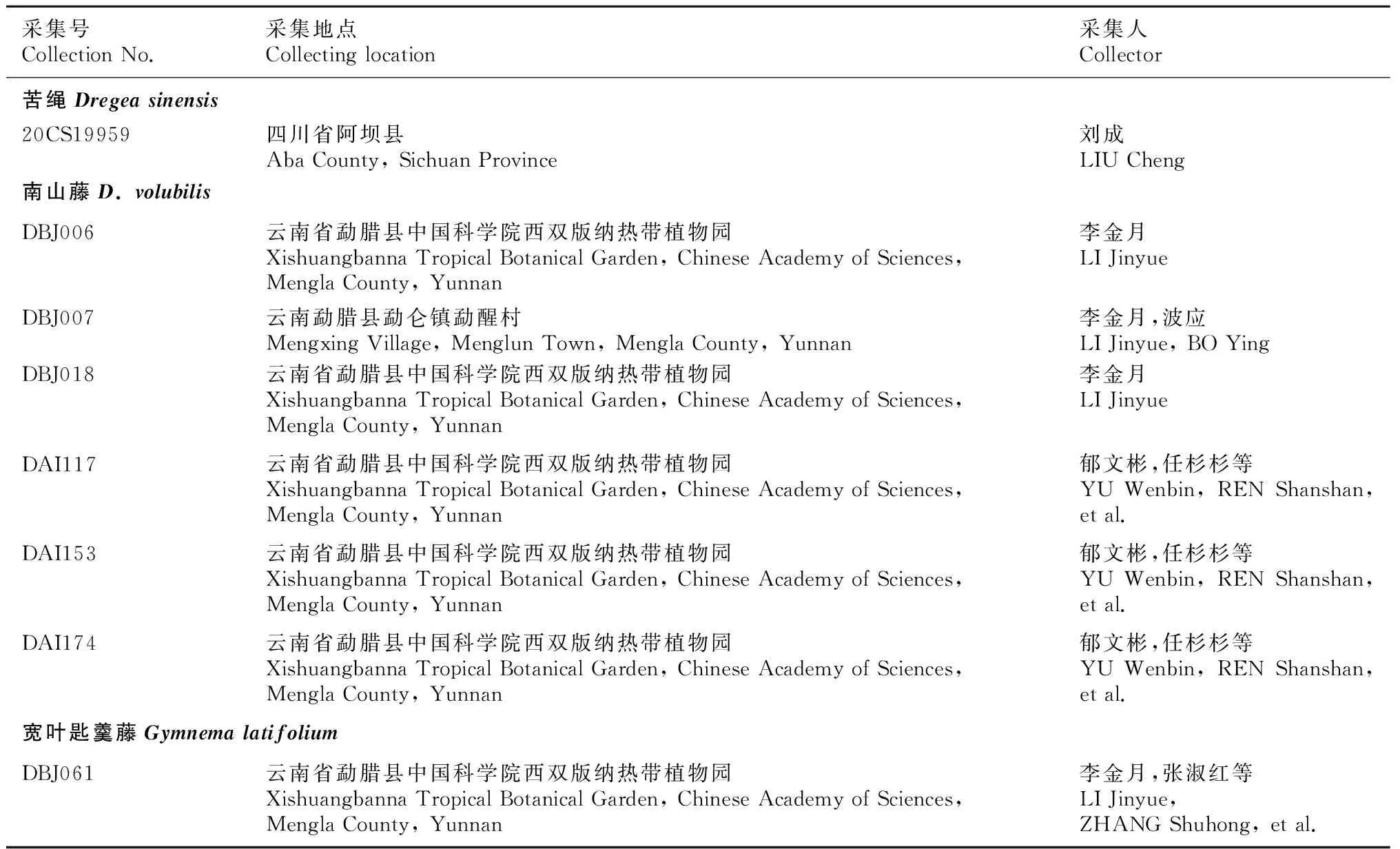
續表 1
1.2 DNA提取、測序和數據處理
對用硅膠干燥后的樣品,先利用改良的十六烷基三甲基溴化銨(CTAB)法提取總DNA (Doyle & Doyle, 1987),再使用瓊脂糖凝膠電泳和Nanodrop分光光度計評估DNA質量。對檢測合格的DNA樣品,使用Covaris 超聲波破碎儀隨機打斷,隨后通過末端修復、加A尾、加測序接頭、純化、PCR擴增、片段篩選等步驟,完成片段長度為350 bp的測序文庫制備。對構建好的測序文庫使用Qubit 2.0對濃度進行檢測,定量范圍為2~1 000 ng,依據溶度進行多樣品混合,并利用Illumina Novaseq 6000進行雙向150 bp讀長測序。對測序原始數據(raw data)直接使用GetOrganelle軟件包 (Jin et al., 2020) 進行葉綠體基因組和核糖體DNA從頭(denovo)組裝。選取長春花(Catharanthusroseus) 的葉綠體基因組(KC561139)和核糖體基因(HQ130657)序列作為參考序列,借助Geneious (Kearse et al., 2012) 對新測物種進行初步注釋,相似性參數設置為70%,并結合ORF (open reading frame)手工調整注釋結果,其中蛋白編碼基因的注釋依據參考序列和開放閱讀框確定起始密碼子和終止子位置。
1.3 系統發育分析
在Geneious軟件中提取新測29份樣品的葉綠體基因片段psbD-trnT和trnL-trnF,以及核糖體基因序列ITS(包括ITS1、5.8S、ITS2三個區),并從GenBank上下載已有的牛奶菜屬42種、南山藤屬1種以及匙羹藤屬2種的psbD-trnT、trnL-trnT以及ITS序列,以吊燈花屬(Ceropegia)植物錐頂吊燈花(Ceropegianilotica)作為外類群(表2)。對3個基因片段的序列分別利用MAFFT 7.450軟件進行多序列自動比對 (Katoh et al., 2013),輸入到Geneious軟件將模糊的區域或空位進行刪除或調整。使用貝葉斯推斷法(Bayesian inference, BI)、最大似然法(maximum likelihood, ML)和最大簡約法(maximum parsimony, MP)對3個片段分別進行單獨和聯合重建。對3個基因片段矩陣分別利用jModeltest 2 2.1.6,選擇貝葉斯信息量準則(Bayesian information criterion, BIC)(Darriba et al., 2012) 篩選出序列矩陣最佳核苷酸替代模型,并采用Mrbayes 3.2.7(Huelsenbeck & Ronquist, 2001)進行貝葉斯系統發生分析。采用最大似然法時選用RAxML-HPC2 on XSEDE 8.2.12工具(Alexandros, 2014) ,選擇GTR+CAT模型,并設置1 000次靴帶值估算分支的支持率(bootstrap support value, BS),使用CIPRES網站在線構建ML樹。采用最大簡約法時使用MEGA 10.2.6軟件(Kumar et al., 2018),使用子樹修剪和嫁接(Subtree-Pruning-Regrafiting, SPR)算法進行分析獲得MP樹(Nei & Kumar, 2000)。構建系統進化樹時使用FigTree 1.4.1 (Price et al., 2009) 進行美化。

表 2 其他取樣物種和序列GenBank登錄號Table 2 List of taxa and GenBank accessions of other sequences used in this study
1.4 形態特征比較
將采集到的“傣百解”樣品與通光散的模式標本進行形態觀察,并進一步比較分析在系統樹上與“傣百解”親緣關系最近的物種,觀察和比較“傣百解”基原植物與其姐妹種之間的形態特征差異。
2 結果與分析
2.1 基因序列特征
核糖體ITS矩陣的長度為768 bp,包含334個變異位點(variable sites),其中202個為位系統發育信息位點(parsimony-informative site)。葉綠體片段psbD-trnT矩陣的長度為1 676 bp, 包含247個變異位點,其中有51個為系統發育信息位點。葉綠體片段trnL-trnF矩陣的長度為1 006 bp,包含142個變異位點,其中有47個為系統發育信息位點。psbD-trnT+trnL-trnF+ ITS序列矩陣全長3 326 bp,包含597個變異位點,其中有305個為系統發育信息位點。
2.2 系統發育分析
同一基因片段矩陣采用貝葉斯法、最大似然法和最大簡約法構建的系統發育樹的拓撲結構基本一致(圖1)。3個片段聯合矩陣得到的系統發育樹比單基因構建的系統發育樹能更好地澄清物種之間的關系(圖1:D)。在單基因系統發育樹中,核糖體ITS矩陣(圖1:C)比葉綠體psbD-trnT矩陣 (圖1:A)和trnL-trnF矩陣(圖1:B)能更好地將物種進行區分和鑒別。
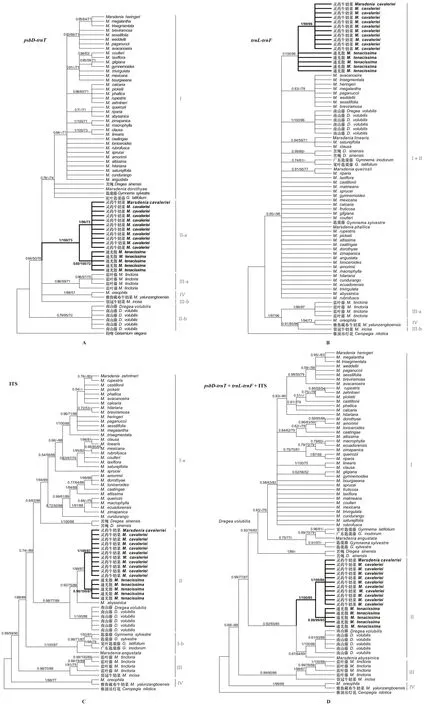
羅馬數字表示主要分支,分支節點上的數值以BIpp/MLBS/MPBS的方式表示貝葉斯后驗概率/最大似然樹自展支持率/最大簡約樹自展支持率,后驗概率不足0.50或支持率不足50的均不標記。Roman numbers indicate main clades, and data upon each node (BIpp/MLBS/MPBS ) indicate Bayesian inference posterior probability / maximum likelihood bootstrap values/ maximum parsimony bootstrap values, and PP below 0.50 and BS below 50 are unlabeled.圖 1 基于葉綠體片段 (psbD-trnT、trnL-trnF) 和核基因ITS構建“傣百解”及其近緣種的系統發育樹Fig. 1 Phylogenetic reconstruction of “Dai-Bai-Jie” and its related species based on two plastids (psbD-trnT and trnL-trnF ) and nuclear ribosome DNA ( ITS ) data
聯合數據矩陣系統發育樹結果表明,牛奶菜屬不是一個單系類群,即南山藤屬和匙羹藤屬物種與牛奶菜屬混合在一起。系統發育分析顯示,牛奶菜族分為4個主要分支,其中雅魯藏布牛奶菜(Marsdeniayarlungzangboensis)與(M.oreophila)構成了分支IV (BIPP/MLBS/MPBS=1.00/99/68),是最早分化出的分支,與其他3個分支構成姐妹關系 (BIPP/MPBS= 0.68/88);隨后是藍葉藤(M.tinctoria)和裂冠牛奶菜(M.inicisa)組成了分支III (BIPP/MLBS/MPBS= 0.99/80/86),并較好支持的是分支I + 分支II的姐妹群(BIPP/MLBS/MPBS= 0.99/77/87)。分支II包括了“傣百解”樣品,以及通光散、靈藥牛奶、南山藤(Dregeavolubilis)和Marsdeniaabyssinica(BIPP/MLBS/MPBS= 0.92/93/85),其中“傣百解”基原植物樣品先與通光散樣品聚在一起形成單系 (BIPP/MLBS/MPBS= 0.92/93/85),再與靈藥牛奶菜(M.cavaleriei)互為姐妹類群,所形成的分支節點支持率較高 (BIPP/MLBS/MPBS= 1.00/100/85),這種關系也得到了單基因片段數據的支持;另外,南山藤與M.abyssinica形成了1個分支(BIPP/MLBS/MPBS= 0.67/64/69)。分支I包括了美洲產的牛奶菜屬物種,以及匙羹藤(Gymnemasylvestre)、寬葉匙羹藤(G.latifolium)、廣東匙羹藤(G.inodorum)和苦繩 ( BIPP/MLBS/MPBS= 0.92/76/82),并且非單系的匙羹藤屬分支包括Marsdeniaabyssinica。
2.3 形態學分析
2.3.1 “傣百解”與通光散模式標本的比較 通過比較“傣百解”基原植物的憑證標本與通光散的模式標本(圖2),發現兩者的葉片和花形態特征基本一致,即莖干和葉片表面密被淡黃色柔毛,葉片在頂端出急驟趨于尖狹,葉基部深心形,近圓形彎缺;花序均為多歧聚散花序,花外部密被絨毛,花冠近鐘狀,花萼5瓣深裂,裂片肉質膨脹,柱頭寬圓筒狀凸起等。因此,進一步證明了“傣百解”的基原為通光散。

圖 2 “傣百解”憑證標本(A)與通光散模式標本(B) 的比較Fig. 2 Comparisons of a voucher specimen of “Dai-Bai-Jie” (A) and the type specimen of Marsdenia tenacissima (B)
2.3.2 通光散與靈藥牛奶菜的形態比較 “傣百解”基原植物與靈藥牛奶菜在系統發育分析中是姐妹類群,但通過比較兩者的形態性狀,發現它們兩者的花冠顏色、葉型、葉面、莖干等相關性狀存在差異(圖3, 圖4)。詳細的特征區要點如表3所示。

圖 3 通光散(A, B) 與靈藥牛奶菜(C, D) 植物圖Fig. 3 Photos of Marsdenia tenacissima (A, B) and Marsdenia cavaleriei (C, D)

A, C, E, G, I. 通光散各部分的形態結構; B, D, F, H, J. 靈藥牛奶菜各部分的形態結構。A, C, E, G, I. Morphological sturcture of each part from M. tenacissima; B, D, F, H, J. Morphological structure of each part from M. cavaleriei.圖 4 通光散與靈藥牛奶菜在葉子、花、莖、根及其橫切面的形態比較Fig. 4 Morphological comparisions of Marsdensia tenacissima and M. cavaleriei about leaf, flower, stem, root and its cross section

表 3 通光散與靈藥牛奶菜形態特征的比較Table 3 Comparison of morphological characters between Marsdenia tenacissima and M. cavaleriei
3 討論與結論
“傣百解”被收錄于2005年版的《云南省中藥材標準》時,其基原植物記載是南山藤屬植物苦繩。基于葉綠體基因psbD-trnT和核糖體基因ITS序列單獨分析和3個片段的聯合分析表明,“傣百解”基原植物樣品和通光散樣品形成1個單系分支,并且與南山藤和M.abyssinica一起屬于分支III。因此,系統發育分析支持“傣百解”的基原植物是通光散,并與靈藥牛奶菜成姐妹關系。在trnL-trnF序列的系統發育樹中“傣百解”基原植物和通光散樣品未聚成1支,而靈藥牛奶菜所有樣品形成1個單系,這可能是由于該基因片段的變異位點較少所造成,因為其他物種的關系也未能得到較好的解析。在系統發育樹上,苦繩與美洲的牛奶菜屬物種和匙羹藤屬物種聚在一起屬于分支I,與“傣百解”基原植物關系較遠。結合《云南省中藥材標準》后面附錄的植物照片和藥材照片,進一步確認“傣百解”的基原就是通光散,這與早期的研究和考證結果一致(李海濤等, 2014)。
值得關注的是,從2012年開始與“傣百解”基原植物相關的考證研究主要基于形態分析和顯微鑒定(管艷紅等, 2012)。由于傳統的鑒定方法會存在局限性,容易受物種本身所處的環境和生長期的影響,因此需要依賴鑒定者的經驗,進而判斷鑒定結果的準確性。已有的研究使用葉綠體基因組序列psbD-trnT、trnL-trnF、核糖體ITS和ETS較好地解析了牛奶菜族的系統發育關系 (Espírito- Santo et al., 2019), 以及利用葉綠體基因rbcL和核糖體ITS2序列能很好地區分一些牛奶菜屬物種與其近緣類群(張忠廉等, 2013),特別是核糖體ITS2序列,在DNA條形碼 (DNA barcoding) 中已被推薦用于物種鑒定的通用條形碼,并已用于藥材市場中通光散及其摻假品的鑒別 (Yu et al., 2018)。當前,植物DNA序列和系統發育的分析方法應用于藥材的分子鑒定已成為植物分類和物種鑒定研究的熱點(陳士林等, 2007)。不過,對于親緣關系接近的物種或姐妹物種,由于單個基因片段因變異度低而使得物種鑒定率低(倪梁紅等, 2014),因此采用變異位點數較多的基因片段或多片段組合可有效提高物種鑒定率(張越等, 2021)。本研究中的葉綠體片段trnL-trnF由于序列變異位點較少,只占所分析序列長度的14.11%,因此系統發育分析顯示許多物種之間的關系未能得到解決。相比而言,核糖體ITS具有較多的變異位點,在系統發育樹中能將大多數物種區分開。因此,本研究推薦使用核糖體ITS,或者使用核糖體ITS與葉綠體片段進行聯合分析,用于“傣百解”基原植物的分子快速鑒定(倪梁紅等, 2014)。
本研究形態特征比較結果顯示,“傣百解”基原植物的憑證標本與通光散的模式標本在葉子形態、花的結構以及植株的被毛等特征基本一致。形態上,“傣百解”基原植物和通光散的莖都密被淡黃色柔毛,葉兩面被絨毛,葉頂端急尖,基部深心形,近似圓形彎缺;莖被毛多,為圓柱形,折斷后有白色乳汁;花序多歧分支,傘形聚傘花序,花冠近鐘狀,裂片肉質膨脹,展開;柱頭凸起,為寬圓筒狀,被花藥附屬物覆蓋 (William et al., 1819; Li et al., 1995)。本研究結果與李海濤等(2014)對“傣百解”基原植物考證結果一致,進一步確定了“傣百解”的基原植物是通光散。從比較通光散與近緣種靈藥牛奶菜的形態結果可以看出,兩者具有明顯的形態差異:(1)靈藥牛奶菜的葉子基部沒有形成圓形彎曲,葉被毛稀疏,表面較為光滑;(2)莖為扁圓柱形,被毛沿著狹窄的兩側生長,花冠顏色與通光散不同,為橙紅色;(3)花冠裂片較淺,張開的幅度較小,直立后稍外展;(4)根表面粗糙,不為粉質,不易剝落。因此,通光散和靈藥牛奶菜可依據根、莖、葉、花的形態性狀差異進行物種鑒別(圖 4)。西雙版納傣藥“傣百解”一直以來備受人們關注,其基原考證首次從分子系統學角度獲得進一步的澄清,將植物的形態特征與其DNA序列相結合可以很好地對物種及其近緣種進行鑒別,并且鑒別效果比單一的傳統鑒定法更佳(金海湘等, 2019; 蘇暢等, 2019; 姚綱和薛彬娥, 2021)。
傣藥,不僅是我國四大民族藥之一,也是我國醫藥文化不可或缺的組成部分。民族醫藥形成于少數民族區域,長久以來,當地的民眾都使用民族藥來治療疾病和保健身體。因其具有良好的效果而得以世代相傳(張夢娜, 2018)。為了使民族藥物資源和民族生物文化多樣性得到保護和持續利用,我們需要對一些常用的傣藥進行系統的整理和物種基原考證,從而能對傣藥的基原植物做準確的鑒定,這是確保臨床上準確、安全用藥的前提(龍春林, 2013)。由于傳統的生藥鑒定法有一定的局限性,主要依賴于經驗,主觀性較強,對于形態學上難以區分的樣品,不同的人依據不用的性狀得到的分類結果也不盡相同,因此結合分子系統學的證據會更有助于物種的鑒定。同時,系統發育分析有助于物種之間親緣關系的梳理,既可能尋找到近緣替代種,又可為“傣百解”基原植物的種質資源綜合評價、系統進化等研究提供有效的科學依據,這對品種整理、鑒定以及保護和開發利用具有重要的意義(段寶忠, 2017)。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
