桑葚幼果的落果與正常果的果柄轉(zhuǎn)錄組分析
2023-03-09 14:53:42陳春兵劉練練查幸福
浙江農(nóng)林大學(xué)學(xué)報(bào) 2023年1期
鄧 璇,陳春兵,鄧 靜,劉練練,李 娟,查幸福
(西南大學(xué) 家蠶基因組生物學(xué)國(guó)家重點(diǎn)實(shí)驗(yàn)室,重慶 400700)
桑Morus是一種重要的木本植物,具有28條染色體[1],不僅可以作為家蠶的重要食物,還具有一定的經(jīng)濟(jì)、藥用及生態(tài)價(jià)值。但是桑葚在成熟的過(guò)程中易脫落,使得桑葚變軟變黑,影響其產(chǎn)業(yè)價(jià)值,因此,如何采用有效的手段對(duì)桑葚脫落進(jìn)行調(diào)控十分重要。
果實(shí)脫落是植物正常發(fā)育過(guò)程中的一種常見(jiàn)現(xiàn)象,但是,在一定程度上會(huì)限制果樹的產(chǎn)量。果實(shí)脫落主要受環(huán)境因素、酶、生理代謝共同調(diào)控[2],其中,環(huán)境因素主要包括生物脅迫(如病蟲害[3])和非生物脅迫(溫度脅迫[4]、水分脅迫[5]、光脅迫[6]等);生理代謝則包括一些激素代謝(如脫落酸[7]、乙烯[8]等植物激素)和糖代謝[9];由于纖維素和果膠是植物細(xì)胞壁的主要組成成分,因此纖維素酶、果膠酶、多聚半乳糖醛酸酶[10]等與果實(shí)脫落相關(guān)[11]。
目前,桑葚脫落的分子機(jī)制還未見(jiàn)報(bào)道。本研究以白桑Morus alba為供試材料,對(duì)其進(jìn)行轉(zhuǎn)錄組測(cè)序分析,探究桑葚在脫落過(guò)程中的生化成分代謝的分子機(jī)制,旨在為進(jìn)一步了解桑葚果實(shí)脫落的分子機(jī)制提供參考。
1 材料與方法
1.1 材料
2021年4月在西南大學(xué)家蠶基因組生物學(xué)國(guó)家重點(diǎn)實(shí)驗(yàn)室桑樹資源種質(zhì)基地,選取無(wú)病害、健康狀況較好的白桑幼果果柄。以落果果柄 (YD) 為實(shí)驗(yàn)組,取6個(gè)果柄為1份,設(shè)3個(gè)生物學(xué)重復(fù),標(biāo)記為YD1、YD2、YD3。正常果果柄 (YN) 為對(duì)照組,同樣取6個(gè)果柄為1份,設(shè)3個(gè)生物學(xué)重復(fù),標(biāo)記為YN1、YN2、YN3。取材后立即在液氮中速凍并儲(chǔ)存于?80 ℃冰箱保存。
1.2 冷凍切片的制作
將正常果和脫落果果柄組織清洗后用質(zhì)量分?jǐn)?shù)為4%的多聚甲醛過(guò)夜固定,用包埋劑包埋材料。待包埋劑徹底凝固后進(jìn)行冷凍切片。切片厚度為5 μm,將切片后的材料吸附于陽(yáng)離子載玻片上,室溫干燥30 min后,用磷酸緩沖鹽溶液(PBS)洗滌樣品,去除包埋劑后置于顯微鏡下觀察。
1.3 RNA 提取
采用TRIzol法提取落果果柄和正常果果柄的總RNA,用分光光度計(jì)檢測(cè)RNA樣品的濃度和純度,以保證是否可以進(jìn)行下一步測(cè)序分析。
1.4 轉(zhuǎn)錄組測(cè)序與分析
樣品由華大基因進(jìn)行測(cè)序及分析,利用華大智造測(cè)序平臺(tái)BGISEQ測(cè)序,所得的原始數(shù)據(jù)為raw reads。過(guò)濾掉低質(zhì)量、接頭污染以及未知堿基N含量過(guò)高的reads,過(guò)濾后的數(shù)據(jù)稱為clean reads。將clean reads比對(duì)到參考基因組上,再進(jìn)行后續(xù)分析。
1.5 測(cè)序原始數(shù)據(jù)的公開(kāi)
將測(cè)序所得原始數(shù)據(jù)提交到美國(guó)國(guó)家生物技術(shù)信息中心(NCBI)的SRA數(shù)據(jù)庫(kù)中,檢索號(hào)為:PRJNA811258。
1.6 差異表達(dá)基因的篩選和富集分析
使用DEseq2方法檢測(cè)樣品之間的差異表達(dá)基因(DEG)[12]。根據(jù)基因本體論數(shù)據(jù)庫(kù)(GO)和京都基因與基金組百科全書(KEGG)注釋結(jié)果以及官方分類,將差異基因進(jìn)行功能分類,同時(shí)使用R軟件中的phyper函數(shù)進(jìn)行富集分析。詳細(xì)說(shuō)明見(jiàn)Wiki網(wǎng)站https://en.wikipedia.org/wiki/Hypergeometric_distribution。校正后的P≤0.05為顯著富集。
1.7 實(shí)時(shí)熒光定量 PCR (RT-qPCR)驗(yàn)證
將桑葚幼果的落果果柄與正常果果柄RNA用 PrimesciptTMRT reagent kit with gDNA Eraser(Takara)反轉(zhuǎn)錄,在 primer premier 5.0 軟件設(shè)計(jì)定量引物 (表 1),內(nèi)參基因?yàn)?Actin,利用 NovoStart?SYBR qPCR SuperMix Plus(novoprotein) 試劑盒進(jìn)行熒光定量 PCR。反應(yīng)程序?yàn)椋?5 ℃ 30 s,95 ℃ 3 s,60 ℃ 30 s,共 40 個(gè)循環(huán)。用 2?ΔΔCt算法處理數(shù)據(jù),利用 GraphPad Prism 8.0.2 軟件作圖。
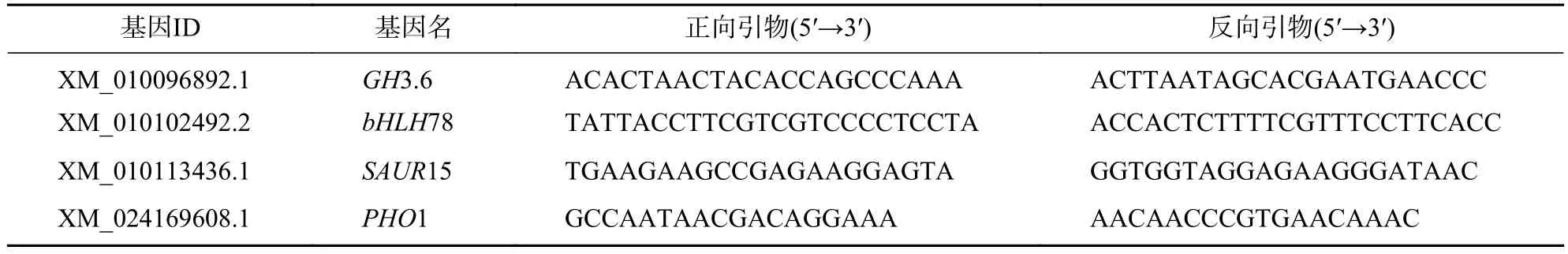
表1 轉(zhuǎn)錄組數(shù)據(jù) RT-qPCR 驗(yàn)證引物序列Table 1 Primer sequences used in RT-qPCR validation of transcriptome data
2 結(jié)果與分析
2.1 果柄離區(qū)切片觀察
桑果是帶果柄脫落,為了研究桑樹正常果柄和脫落果柄的差異,對(duì)其進(jìn)行冷凍切片并觀察顯微結(jié)構(gòu)。從圖1可知:正常幼果果柄與枝干相連的區(qū)域內(nèi)細(xì)胞規(guī)則,形態(tài)一致,而脫落幼果果柄與枝干相連處細(xì)胞小且致密。
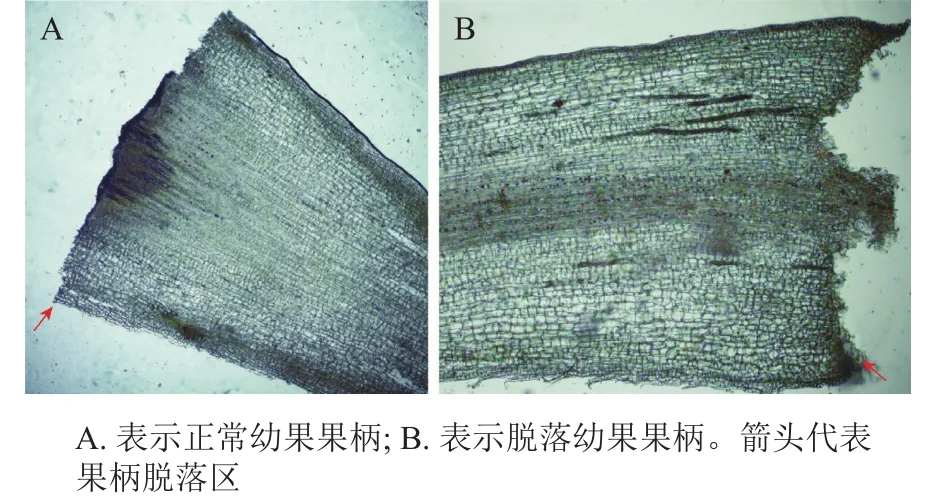
圖1 落果與正常果果柄縱切面顯微結(jié)構(gòu)圖Figure 1 Longitudinal section microscopic structures of abscised and normal surviving young fruit peduncles
2.2 轉(zhuǎn)錄組測(cè)序結(jié)果質(zhì)量分析
基于BGISEQ對(duì)桑葚幼果的落果與正常果果柄離區(qū)進(jìn)行轉(zhuǎn)錄組測(cè)序,6個(gè)樣品獲得262.92 Mb的原始序列。原始測(cè)序數(shù)據(jù)經(jīng)一系列質(zhì)量控制后,每個(gè)樣本獲得超過(guò)6.3 Gb的高質(zhì)量純凈測(cè)序數(shù)據(jù)量,每項(xiàng)堿基質(zhì)量大于20的堿基數(shù)量占總堿基數(shù)量的比例 (Q20) 均大于96%,每項(xiàng)堿基質(zhì)量大于30的堿基數(shù)量占總堿基數(shù)量的比例 (Q30) 均大于91%。將每個(gè)樣本的高質(zhì)量純凈序列與測(cè)序并組裝的桑樹基因組序列進(jìn)行比對(duì),比對(duì)率均高于59%(表2),表明本研究轉(zhuǎn)錄組測(cè)序數(shù)據(jù)質(zhì)量較高,可用于后續(xù)的分析。
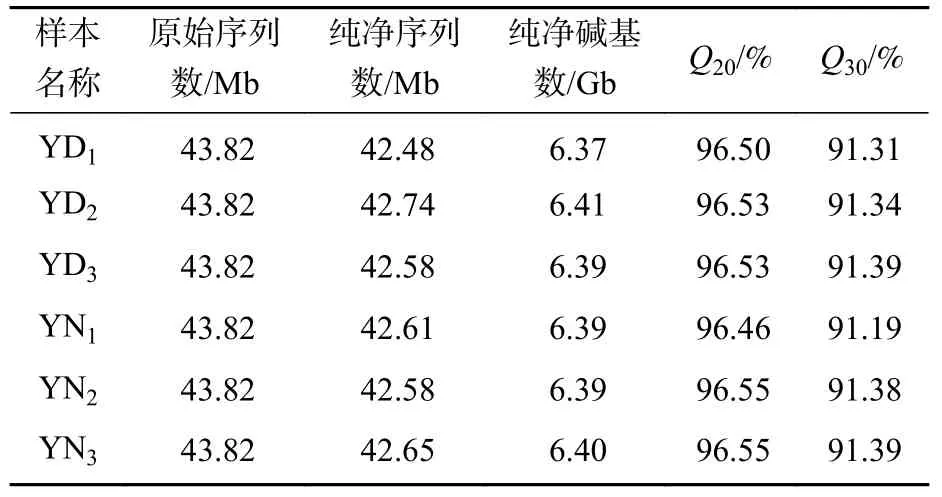
表2 測(cè)序數(shù)據(jù)統(tǒng)計(jì)Table 2 Statistics of sequencing data
2.3 落果與正常果果柄的差異表達(dá)基因
用每2個(gè)樣品之間的Pearson相關(guān)系數(shù)以反映樣本間基因表達(dá)的相關(guān)性(圖2)。結(jié)果發(fā)現(xiàn):Pearson相關(guān)系數(shù)為0.69~1.00,說(shuō)明各樣本間重復(fù)性和相關(guān)性較好。
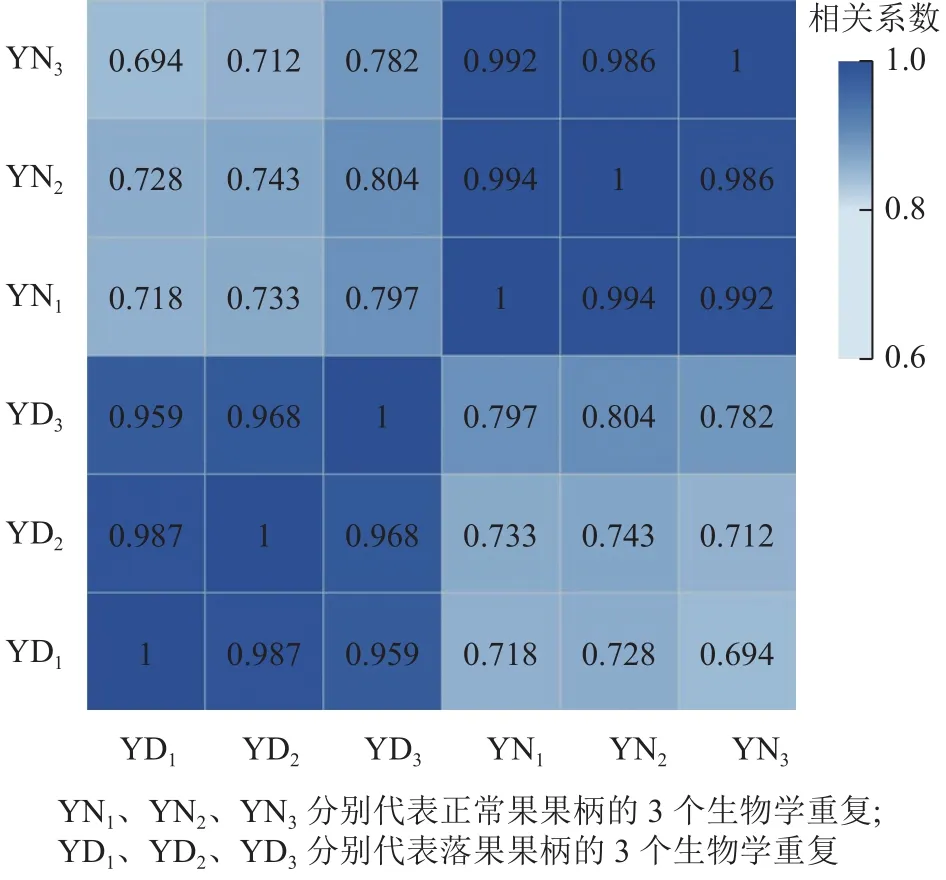
圖2 落果與正常果果柄的相關(guān)性熱圖Figure 2 Correlation heatmap of abscised and normal surviving young fruit peduncles
在桑葚幼果果柄組織中,共鑒定到25 293個(gè)基因(圖3A)。其中,共表達(dá)的基因有23 203個(gè),占91.7%。正常果果柄特有基因?yàn)? 164個(gè),占4.6%,落果果柄組特有基因?yàn)?26個(gè),占3.7%。由此可見(jiàn),落果果柄基因較正常果果柄基因少。根據(jù)差異篩選標(biāo)準(zhǔn),在2組中共篩選到10 481個(gè)差異表達(dá)基因(圖3B),其中,5 239個(gè)差異表達(dá)基因上調(diào)表達(dá),5 242個(gè)差異表達(dá)基因下調(diào)表達(dá)。

圖3 落果與正常果果柄的表達(dá)基因和差異表達(dá)基因Figure 3 Expressed genes and differentially expressed genes of abscised and normal surviving young fruit peduncles
2.4 差異表達(dá)基因的 GO 富集分析
本研究對(duì)差異表達(dá)基因進(jìn)行GO功能富集分析,共發(fā)現(xiàn)37個(gè)顯著性GO條目。生物過(guò)程共富集到7 064個(gè)基因,顯著富集到19個(gè)條目,涉及生物調(diào)節(jié)、細(xì)胞過(guò)程、代謝過(guò)程、對(duì)刺激的反應(yīng)等,其中富集到生物過(guò)程中最多的差異表達(dá)基因?yàn)榧?xì)胞過(guò)程和代謝過(guò)程基因;細(xì)胞組分共富集到6 857個(gè)基因,顯著富集到6個(gè)條目,涉及細(xì)胞解剖實(shí)體、細(xì)胞內(nèi)、其他有機(jī)部分、含蛋白質(zhì)復(fù)合物、病毒粒子等,富集到細(xì)胞組分最多的差異表達(dá)基因?yàn)榧?xì)胞解剖實(shí)體和細(xì)胞內(nèi)基因;分子功能共富集到9 842個(gè)基因,顯著富集到12個(gè)條目,涉及催化活性、轉(zhuǎn)運(yùn)蛋白活性、轉(zhuǎn)錄調(diào)節(jié)活性、分子功能調(diào)節(jié)劑等,富集到分子功能最多的差異表達(dá)基因?yàn)榇呋钚院徒Y(jié)合基因(圖 4)。
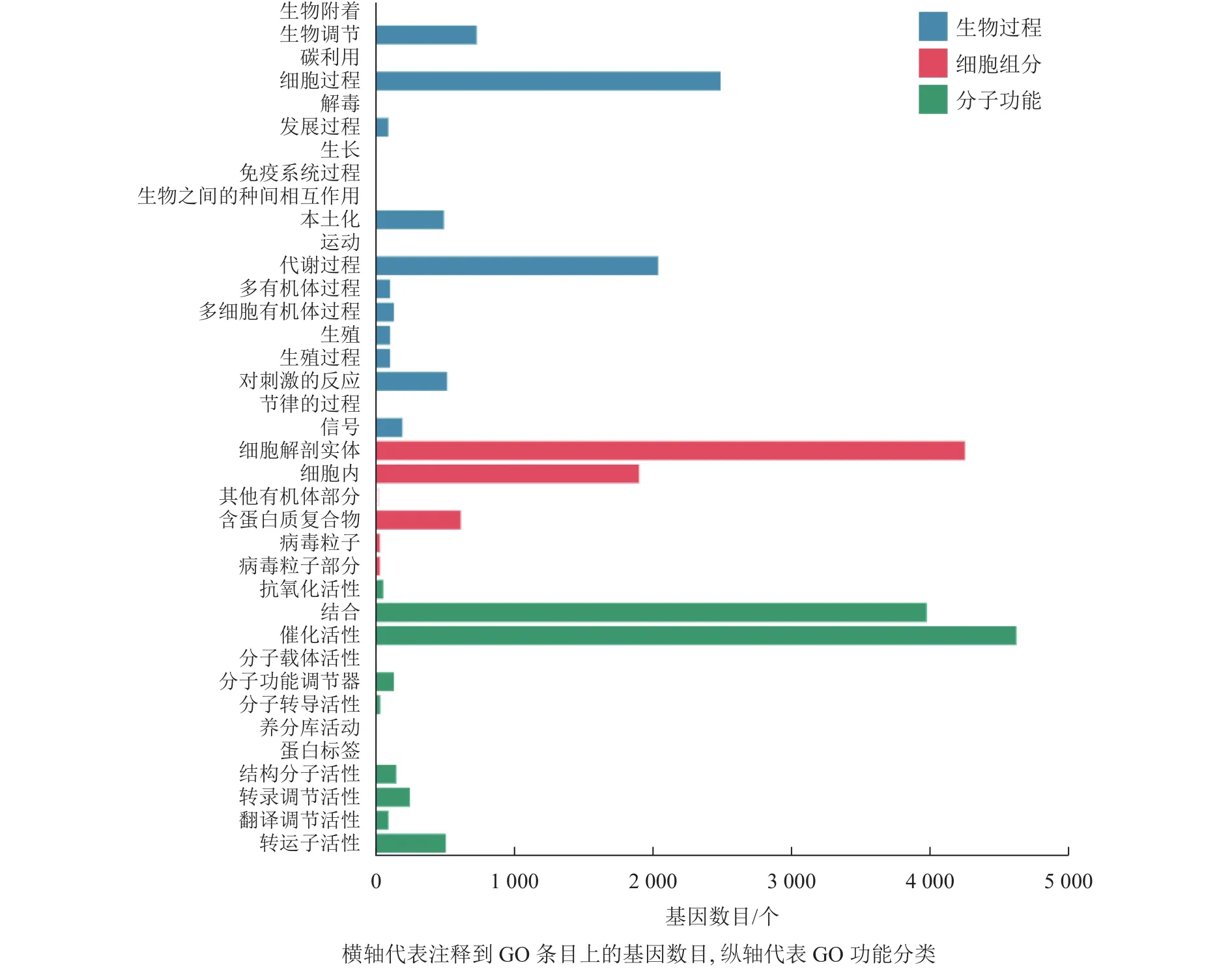
圖4 落果與正常果果柄的差異表達(dá)基因GO分類柱狀圖Figure 4 Histogram of GO classification of differentially expressed genes of abscised and normal surviving young fruit peduncles
GO富集分析結(jié)果顯示:大多數(shù)差異基因集中在催化活性、膜的組成成分、氧化還原酶活性、膜的內(nèi)在成分,分別為 4 623、2 774、777、1 781 個(gè)基因(圖5)。其中,具催化活性的基因最多,說(shuō)明在桑葚的落果過(guò)程中可能有很多重要的酶參與,從而發(fā)生一系列的催化反應(yīng)。
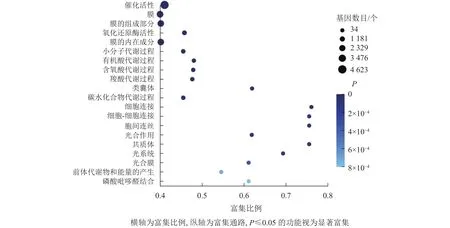
圖5 落果與正常果果柄的差異表達(dá)基因GO富集氣泡圖Figure 5 GO enrichment bubble chart of differentially expressed genes of abscised and normal surviving young fruit peduncles
2.5 差異表達(dá)基因的KEGG代謝通路富集分析
在本研究中,細(xì)胞過(guò)程富集的基因?yàn)?80個(gè),富集到1個(gè)條目,為運(yùn)輸和分解代謝;環(huán)境信息處理共富集到653個(gè)基因,富集到2個(gè)條目,為膜運(yùn)輸和信號(hào)轉(zhuǎn)導(dǎo);遺傳信息處理共富集到1 558個(gè)基因,包含4個(gè)條目,為折疊分類和降解、復(fù)制和修復(fù)、轉(zhuǎn)錄、翻譯;代謝通路富集到5 315個(gè)基因,包含11個(gè)條目,為氨基酸代謝、其他次生代謝物的生物合成、碳水化合物代謝、能量代謝等;有機(jī)系統(tǒng)共富集到427個(gè)基因,含1個(gè)條目(圖6)。分析發(fā)現(xiàn):代謝通路富集的基因最多,說(shuō)明桑葚在脫落過(guò)程中代謝反應(yīng)尤為明顯。

圖6 落果與正常果果柄的差異表達(dá)基因KEGG分類柱狀圖Figure 6 Histogram of KEGG classification of differentially expressed genes of abscised and normal surviving young fruit peduncles
KEGG通路富集分析結(jié)果顯示:大多數(shù)差異表達(dá)基因集中在MAPK信號(hào)通路、黃酮類生物合成、檸檬酸循環(huán)、植物激素信號(hào)轉(zhuǎn)導(dǎo)、氨基酸的生物合成通路,分別含293、102、55、318、251個(gè)基因(圖7)。其中,植物激素信號(hào)轉(zhuǎn)導(dǎo)途徑的差異表達(dá)基因數(shù)量最多,說(shuō)明在桑葚果實(shí)脫落過(guò)程中植物激素起到了十分重要的作用。
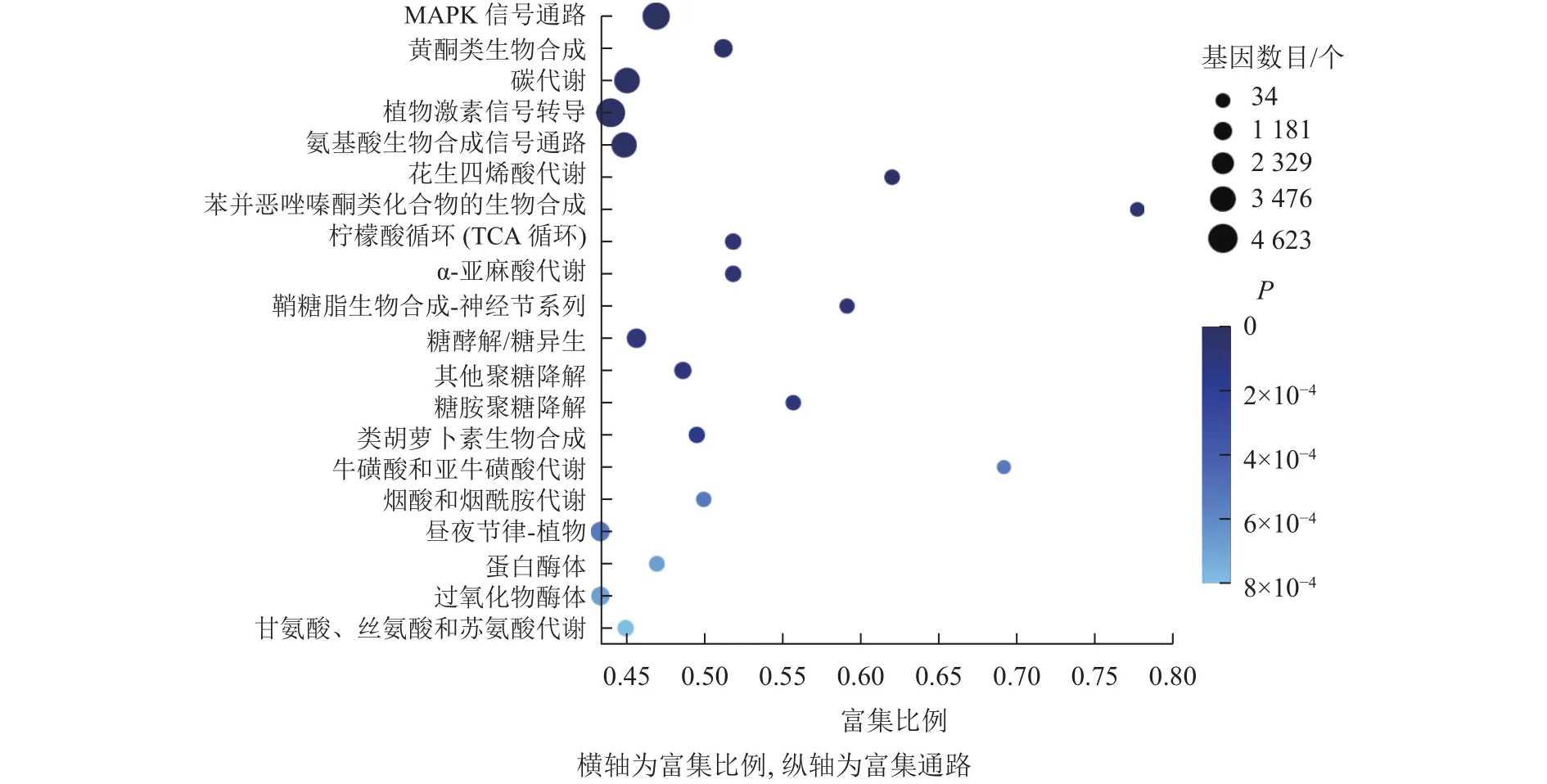
圖7 落果與正常果果柄的差異表達(dá)基因的KEGG富集氣泡圖Figure 7 KEGG enrichment bubble diagram of differentially expressed genes of abscised and normal surviving young fruit peduncles
在桑樹的代謝通路中,植物激素信號(hào)轉(zhuǎn)導(dǎo)途徑的差異表達(dá)基因數(shù)量最多,為318個(gè)基因,其中,落果果柄中有51.9% (156個(gè))的差異表達(dá)基因的表達(dá)量高于正常果果柄(圖8A);在黃酮類生物合成途徑中,有102個(gè)差異基因富集,其中有73.5% (75個(gè))的差異基因在落果果柄中表達(dá)量高(圖8B);在氨基酸生物合成通路中,共有251個(gè)差異表達(dá)基因富集,其中有59.8% (150個(gè))的差異表達(dá)基因在落果果柄中表達(dá)量更高(圖8C);在檸檬酸循環(huán)中,有55個(gè)差異表達(dá)基因富集,其中有74.5% (41個(gè))的差異表達(dá)基因在落果果柄中表達(dá)量更高(圖8D)。不難發(fā)現(xiàn),落果果柄有超過(guò)一半的差異表達(dá)基因表達(dá)量高于正常果果柄。說(shuō)明植物激素、黃酮類等次生代謝物以及檸檬酸等物質(zhì)在桑葚脫落過(guò)程中發(fā)揮了重要的作用。
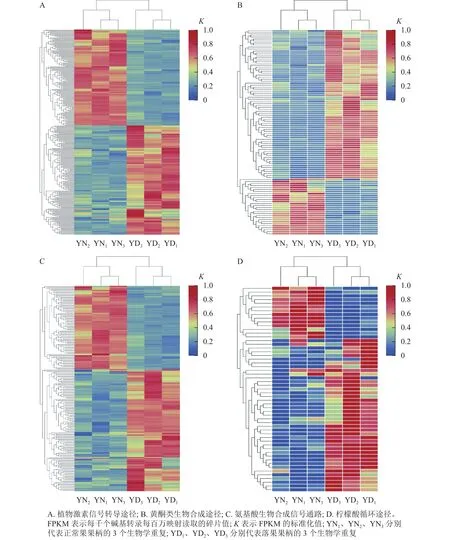
圖8 落果與正常果果柄的代謝途徑的差異表達(dá)基因熱圖Figure 8 Heat map of differentially expressed genes in metabolic pathways of abscised and normal surviving carpopodium
2.6 轉(zhuǎn)錄組測(cè)序數(shù)據(jù)的 RT-qPCR 驗(yàn)證
由于植物激素對(duì)于落花落果以及保花保果都具有十分重要的作用,于是在植物激素信號(hào)轉(zhuǎn)導(dǎo)途徑中篩選4 個(gè)差異十分顯著的基因,分別為GH3.6 (XM_010096892.1)、bHLH78 (XM_010102492.2)、SAUR15 (XM_010113436.1)、PHO1 (XM_024169608.1)。將這4個(gè)基因的每千個(gè)堿基轉(zhuǎn)錄每百萬(wàn)映射讀取的碎片值(FPKM)繪制直方圖,GH3.6在落果組中的表達(dá)水平是正常果組的20多倍,bHLH78在落果組中的表達(dá)水平約為正常果組20倍,PHO1(磷轉(zhuǎn)運(yùn)蛋白)在落果組中的表達(dá)水平是正常果組的30多倍,SAUR15在落果組中的表達(dá)水平是正常果組的100多倍(圖9A)。由此推測(cè),這4個(gè)基因均參與了桑葚的果實(shí)脫落。為了驗(yàn)證轉(zhuǎn)錄組數(shù)據(jù)的準(zhǔn)確性,對(duì)這4個(gè)基因進(jìn)行RT-qPCR驗(yàn)證,結(jié)果發(fā)現(xiàn):4個(gè)差異表達(dá)基因的熒光定量相對(duì)表達(dá)量的變化趨勢(shì)與轉(zhuǎn)錄組表達(dá)趨勢(shì)一致,說(shuō)明該轉(zhuǎn)錄組數(shù)據(jù)可靠(圖9B)。
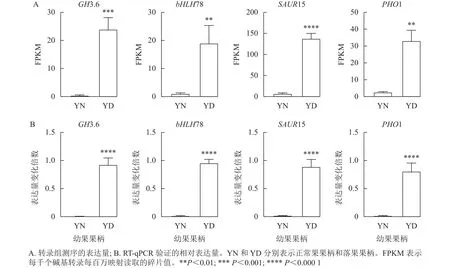
圖9 4 個(gè)差異表達(dá)基因的 RT-qPCR 驗(yàn)證Figure 9 RT-qPCR validation of four differentially expressed genes
3 討論
近年來(lái),為了提高果樹產(chǎn)量,越來(lái)越多果實(shí)脫落的相關(guān)研究被報(bào)道,如番茄Lycopersicon esculentum[13]、荔枝Litchi chinensis[14]、扁桃Amygdalus communis[15]等。桑樹作為一種具有重要經(jīng)濟(jì)價(jià)值的植物,其生理落果引起學(xué)者們的關(guān)注。2021年,有研究者對(duì)不同種類的長(zhǎng)果桑Morus macroura的生理落果進(jìn)行研究,發(fā)現(xiàn)高濃度的脫落酸(ABA)和乙烯(ETH)能夠促進(jìn)落果,而高濃度的赤霉素(GA3)和生長(zhǎng)素(IAA)抑制落果[16]。但是,關(guān)于桑樹果實(shí)脫落的分子機(jī)制還尚不清楚。
植物激素是植物生長(zhǎng)發(fā)育過(guò)程中十分重要的物質(zhì),參與植物的落花落果,與落花落果常見(jiàn)的相關(guān)植物激素有IAA、GA3、細(xì)胞分裂素、ETH、ABA[17]。
IAA主要是促進(jìn)植物的生長(zhǎng)發(fā)育,抑制果實(shí)的脫落,如果生長(zhǎng)素在運(yùn)輸途徑中受到阻礙則會(huì)導(dǎo)致植物果實(shí)的脫落[18?19]。GH3.6是吲哚乙酸酰胺合成酶的基因,能夠催化IAA氨基化,使生長(zhǎng)素失活[20],這與本研究的結(jié)論一致。Small auxin-up RNA(SAUR) 基因是一類生長(zhǎng)素早期響應(yīng)基因,SAUR15能夠調(diào)控植物的生長(zhǎng)發(fā)育并參與環(huán)境脅迫響應(yīng)[21]。在本研究中,SAUR15的表達(dá)水平在落果組高于正常果組100多倍,說(shuō)明SAUR15基因可能參與桑葚果實(shí)脫落;ABA主要是促進(jìn)果實(shí)脫落[7],這是由于ABA能夠增加纖維素酶的活性進(jìn)而促進(jìn)果實(shí)脫落[22]。有學(xué)者通過(guò)研究無(wú)核荔枝ABA合成關(guān)鍵酶LcNCED與生理落果的關(guān)系,驗(yàn)證了ABA對(duì)于落果的作用[14];GA主要是通過(guò)作用于IAA影響果實(shí)脫落[23];ETH主要促進(jìn)果實(shí)的成熟、衰老、脫落,這在多種植物中被報(bào)道,如番茄[13]等;細(xì)胞分裂素能夠促進(jìn)坐果,延遲果實(shí)的脫落[24]。
此外,在對(duì)落果的研究中還發(fā)現(xiàn)了大量的基因和轉(zhuǎn)錄因子的調(diào)控,如JOINTLESS[25]、LATERAL SUPPRESSOR(LS)[26]、MACROCALYX[27]主要調(diào)控離區(qū)的形成,MYB[28]、WRKY[29]、bHLH[30]、bZIP[31]等轉(zhuǎn)錄因子能參與植物的器官脫落。bHLH78屬于bHLH轉(zhuǎn)錄因子,參與植物生長(zhǎng)和代謝[32],調(diào)節(jié)花青素的生物合成[33]。本研究中,bHLH78的表達(dá)水平在落果組較高,說(shuō)明其可能參與桑葚的脫落。此外,還有一些重要的酶及蛋白調(diào)控植物的器官脫落,如纖維素酶、果膠酶、多聚半乳糖醛酸酶、擴(kuò)展蛋白等[34]。在本研究中,磷轉(zhuǎn)運(yùn)蛋白(PHO1)在落果果柄組的表達(dá)水平是正常果柄組的30多倍。此前研究表明:ABA調(diào)控依賴于PHO1的表達(dá)[35],說(shuō)明PHO1基因可能參與桑葚的脫落。
4 結(jié)論
本研究GO富集分析結(jié)果顯示:有4 623個(gè)基因具有催化活性,說(shuō)明在果實(shí)脫落過(guò)程中,有多種重要的酶發(fā)揮催化效應(yīng);KEGG通路富集分析結(jié)果顯示:大多數(shù)差異表達(dá)基因集中在黃酮類生物合成、檸檬酸循環(huán)、植物激素信號(hào)轉(zhuǎn)導(dǎo)、氨基酸的生物合成等通路中,說(shuō)明在果實(shí)脫落過(guò)程中,植物激素、糖類、次生代謝物質(zhì)等發(fā)揮了重要的作用,從而調(diào)控果實(shí)的脫落。本研究篩選了4個(gè)顯著的差異表達(dá)基因,均在桑葚脫落過(guò)程中參與反應(yīng),可為今后進(jìn)一步研究桑樹的果實(shí)脫落提供參考。
猜你喜歡
音樂(lè)探索(2022年2期)2022-05-30 21:01:37
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
小天使·一年級(jí)語(yǔ)數(shù)英綜合(2019年8期)2019-08-27 02:23:00
電子制作(2018年18期)2018-11-14 01:48:24
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學(xué)周刊·兒童版(2015年1期)2015-07-07 04:12:52
