白菜TLP基因家族鑒定及表達分析
2023-03-06 07:09:00王紫雯陳雨吳曉宇趙玉梅任鑫如呂金枝李嘉利段巧紅黃家保
山東農(nóng)業(yè)科學 2023年1期
關鍵詞:分析
王紫雯陳雨吳曉宇趙玉梅任鑫如呂金枝李嘉利段巧紅黃家保
(山東農(nóng)業(yè)大學園藝科學與工程學院/作物生物學國家重點實驗室,山東 泰安 271018)
TLP(Tubby-like protein)最早在肥胖小鼠中發(fā)現(xiàn),并且廣泛存在于真核生物中[1]。植物TLP蛋白C末端通常含有一個由270個氨基酸組成的Tubby保守結構域,該結構域由一個包含12條反向平行鏈和一個中央疏水的α螺旋組成,能與特定的磷脂酰肌醇4,5-二磷酸(PIP2)結合,從而與質膜連接[2,3]。與保守的C末端Tubby結構域相反,TLP的N端在不同物種間存在較大差異,動物TLP蛋白N末端高度分化,大多數(shù)植物TLP蛋白N末端含有一個保守的F-box結構域,該結構域同蛋白泛素化密切相關[4,5],且有研究表明Tubby結構域和F-box結構域是協(xié)同進化的[6]。
動物中TLP基因的生物學功能已經(jīng)得到深入研究,例如,在人體中發(fā)現(xiàn)TLP基因突變可能會導致肥胖、視力喪失和神經(jīng)感覺喪失等[7,8]。在植物中發(fā)現(xiàn)了更多的TLP基因,但是研究較少,許多功能機制仍不清楚[9]。已有的研究更多集中在植物對非生物脅迫的響應,目前已在擬南芥、水稻、番茄、玉米、楊樹中分別鑒定出了11、14、11、15、11個TLP基因家族成員[6,10-12]。通過酵母雙雜交試驗發(fā)現(xiàn),擬南芥TLP家族成員At-TLP3參與了種子萌發(fā)過程中脫落酸(ABA)信號的調(diào)節(jié),AtTLP9在鹽脅迫與干旱脅迫中發(fā)揮作用[10,13]。在鷹嘴豆中,CaTLP1在脫水、高鹽、脫落酸(ABA)處理下表達上調(diào),過表達CaTLP1可以提高轉基因煙草植株對脫水、鹽脅迫和氧化脅迫的耐受性,同時改善植株地上部和根構型,其在細胞核中的優(yōu)先表達可能與抗逆性增強有關[14]。過表達MdTLP7基因可以顯著提高擬南芥幼苗對滲透脅迫、鹽脅迫、冷熱脅迫的耐受性[15]。這都證明了TLP基因參與了植物的非生物脅迫反應,在植物應對非生物脅迫響應的過程中發(fā)揮重要作用。
白菜(Brassica rapa)為十字花科蕓薹屬蔬菜,是中國、韓國和日本等國的主要蔬菜之一,在世界各地廣泛種植。目前,土壤鹽堿化成為制約我國農(nóng)業(yè)可持續(xù)發(fā)展的重要因素之一,我國部分白菜產(chǎn)區(qū)土壤鹽漬化嚴重影響白菜的正常生長發(fā)育過程,因此,亟需培育耐鹽、抗鹽的白菜品種。現(xiàn)有研究表明,TLP家族在植物對非生物脅迫的響應中發(fā)揮一定作用,所以本研究對白菜TLP家族進行了鑒定,并對其基因結構及系統(tǒng)發(fā)育等系列生物信息學進行了分析,同時,對其組織表達模式及鹽脅迫響應情況進行了研究,以期為其后續(xù)功能研究及種質改良奠定基礎。
1 材料與方法
1.1 白菜TLP家族成員鑒定及理化性質分析
根據(jù)擬南芥已經(jīng)鑒定出來的11個TLP成員蛋白序列,通過雙向Blast在白菜基因組搜索候選白菜TLP家族成員,再通過保守結構域分析等方法進一步確定候選基因。應用軟件TBtools[16]和Expasy(https://www.expasy.org/)[17]分析白菜TLP家族的蛋白質分子量(MW)、等電點(pI)等理化性質。
1.2 白菜TLP基因家族染色體定位、共線性分析及進化樹構建
從白菜數(shù)據(jù)庫下載gff3基因組注釋數(shù)據(jù)文件,使用TBtools繪制TLP基因在染色體上的分布圖。利用MCScanX[18]繪制白菜與擬南芥TLP基因家族共線性分析圖。在Ensembl Plants(http://plants.ensembl.org/index.html)獲取番茄及水稻TLP家族氨基酸序列,通過MEGA-X[19]采用最大似然法(Maximum Likelihood Estimate,MLE)構建白菜、擬南芥、番茄及水稻TLP家族系統(tǒng)進化樹。
1.3 白菜TLP基因結構和保守結構域分析
使用TBtools繪制白菜TLP基因結構圖;通過MEME(http://meme-suite.org/)[20]預測白菜TLP蛋白保守基序;利用軟件DNAMAN對白菜TLP序列進行比對,分析其保守氨基酸位點。
1.4 順式作用元件分析
通過Ensembl Plants[21]查找并下載白菜TLP基因家族的起始密碼子上游2 000 bp序列,使用PlantCARE(http:/bioinformatics.psb.ugent.be/webtools/plantcare/htm1/)在線網(wǎng)站[22]對獲取的序列進行分析。
1.5 白菜TLP基因表達分析
試驗材料為山東農(nóng)業(yè)大學蔬菜生長與生殖研究組植物培養(yǎng)室的‘848蛋黃白’,于2022年2月20日定植,分別取定植70 d白菜幼苗的根、莖、葉和當天開放的花、果莢進行RNA提取,用于組織特異性表達研究;使用150 mmol/L NaCl分別處理0、2、4、6、12 h,各部位及各時間段均取3個生物學重復,進行基因表達分析。
通過qPrimerDB-qPCR Primer Database(https:/biodb.swu.edu.cn/qprimerdb/)[23]查找并設計引物(表1),內(nèi)參基因為BrActin7,由青島擎科梓熙生物技術有限公司合成。采用RNA提取試劑盒提取白菜樣品的總RNA,參照HiScriptⅡQ RT SuperMix for qPCR(南京諾唯贊生物科技有限公司)試劑盒產(chǎn)品說明書進行反轉錄得到cDNA,用ChamQ Universal SYBR qPCR Master Mix(南京諾唯贊生物科技有限公司)試劑盒進行定量PCR試驗。反應體系(20μL):上、下游引物各0.4 μL,SYBR 10μL,cDNA 2μL,用滅菌ddH2O補至20μL。PCR反應程序:95℃30 s;95℃10 s,60℃22 s,40個循環(huán)。數(shù)據(jù)使用Microsoft Excel進行整理,并使用TBtools進行熱圖繪制。

表1 白菜TLP基因序列引物
2 結果與分析
2.1 白菜TLP基因家族成員鑒定及理化性質分析
經(jīng)鑒定得到14個白菜TLP基因家族成員,命名為BrTLP1~BrTLP14(表2),這些基因不均勻地分布在白菜10條染色體中的7條,BrTLP1、BrTLP8、BrTLP13未在染色體中發(fā)現(xiàn),根據(jù)亞細胞定位預測其可能分布于葉綠體或線粒體中。

表2 白菜TLP基因家族信息
白菜TLP家族蛋白分子量介于38 735.69~52 747.34 D,等電點介于9.16~9.88。所有TLP家族成員的等電點均大于7.0,說明其均富含堿性氨基酸。
2.2 白菜TLP蛋白保守結構域分析
對白菜的14個TLP蛋白序列進行保守性分析,每個基因均含有Motif 2、Motif 3、Motif 7、Motif 10。除了BrTLP14,其他基因的Motif分布都比較均勻。Motif 9僅在BrTLP3、BrTLP4、BrTLP5上出現(xiàn),BrTLP14中Motif的數(shù)目最少,與其他基因成員差異明顯,推測該基因在功能上與其他基因存在較大不同(圖1)。

圖1 白菜TLP基因家族結構域分析
2.3 白菜TLP基因家族結構分析
在鑒定到的14個BrTLP基因中,BrTLP14含有9個CDS,絕大部分BrTLP基因含有4~5個CDS(圖2)。BrTLP14的CDS數(shù)目與其他基因存在較大差別,推測其可能在白菜器官中發(fā)揮特異性功能。對蛋白的保守功能域進行分析,發(fā)現(xiàn)絕大多數(shù)白菜TLP家族基因都含有F-box結構域,其中Motif 4序列為F-box結構域序列信息。4個保守功能域分別對應Motif 4、Motif 6、Motif 5、Motif 1保守基序(圖3)。

圖2 白菜TLP基因家族結構分析

圖3 白菜TLP家族氨基酸序列比對
2.4 白菜與擬南芥、番茄、水稻TLP家族系統(tǒng)發(fā)育分析
由系統(tǒng)發(fā)育樹(圖4)可知,TLP家族被劃分為3個亞族,其中亞族A包括1個BrTLP、1個AtTLP和1個OsTLP,亞族B包括3個BrTLP、2個AtTLP、4個SlTLP和1個OsTLP,亞族C包括10個BrTLP、8個AtTLP、7個SlTLP和12個Os-TLP。白菜和擬南芥、番茄、水稻的TLP家族成員進化關系較近,推測其直系同源基因之間可能存在相似的功能。

圖4 白菜、擬南芥、番茄、水稻TLP家族進化樹
2.5 白菜TLP基因家族染色體定位及與擬南芥基因的共線性分析
通過TBtools工具對白菜與擬南芥的TLP基因家族成員進行共線性分析(圖5),結果顯示,有10個白菜TLP家族成員在擬南芥TLP家族中找到了對應的直系同源基因,表明TLP基因家族在白菜與擬南芥間有較高的保守性。
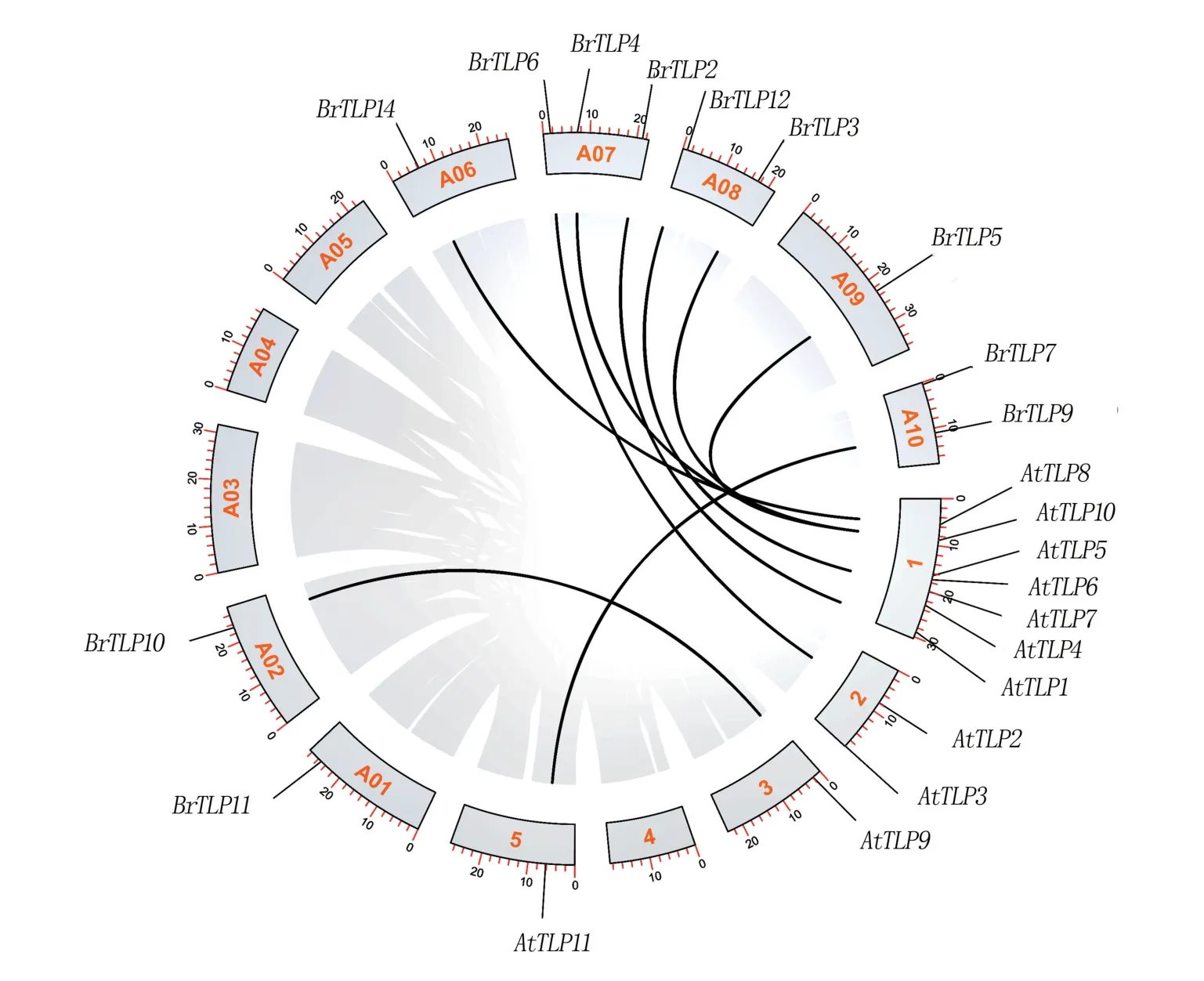
圖5 白菜與擬南芥TLP基因家族共線性分析
根據(jù)基因位置信息對14個BrTLP基因家族染色體進行定位分析(圖6),結果顯示其不均勻分布在大白菜7條染色體上,其中7號染色體上的基因最多,有3個成員;其次是8號、10號染色體上有2個基因成員;1號、2號、6號、9號分別各有1個基因成員。

圖6 白菜TLP基因在染色體上的分布
2.6 白菜TLP基因家族順式作用元件分析
通過對白菜TLP基因家族啟動子上游2 000 bp的序列進行順式作用元件分析(圖7)發(fā)現(xiàn),14條白菜TLP基因中均含有光響應元件,除了BrTLP9、BrTLP11、BrTLP14,其他11個基因均含有脫落酸響應元件,且BrTLP12最多;除了BrTLP4、BrTLP5、BrTLP10、BrTLP12、BrTLP13,其他基因均含有赤霉素響應元件和生長素響應元件;除了BrTLP7,其他基因均含有厭氧響應位點,且BrTLP14含量最多,推測其可能在厭氧逆境脅迫中起重要作用。除了BrTLP4、BrTLP8、BrTLP11、BrTLP14、BrTLP13,其他9個基因均含有茉莉酸甲酯響應元件,6個白菜TLP基因含有水楊酸響應元件和干旱響應元件。

圖7 白菜TLP基因啟動子順式作用元件分析
2.7 白菜TLP基因家族亞細胞定位預測
白菜TLP基因家族主要集中在細胞核、線粒體和細胞質基質(表3),其次為葉綠體、細胞質和細胞骨架。細胞核中所有的基因均有定位,在線粒體中僅有BrTLP2、BrTLP3、BrTLP14沒有定位信號,僅有BrTLP6定位在內(nèi)質網(wǎng)中。每個基因在不同細胞器中的表達位置和表達量各不相同,推測這與基因所執(zhí)行的功能有關。

表3 白菜TLP基因家族亞細胞定位預測
2.8 白菜TLP基因家族表達分析
2.8.1 組織特異性表達分析 以根的表達量為對照,大多數(shù)BrTLPs在白菜花中表達量較高,在莖、葉和果莢中的表達量與根相似。其中BrTLP14在果莢、葉和莖中的表達量最高,BrTLP5在花中的表達量最高,同時在花中表達量較高的基因還有BrTLP8、BrTLP10(圖8)。

圖8 BrTLPs在大白菜不同器官中的表達
2.8.2 鹽脅迫下的基因表達分析 由圖9可知,隨著鹽脅迫時間的延長,BrTLP2、BrTLP6兩個基因的相對表達量呈現(xiàn)升高的趨勢;BrTLP4、BrTLP7、BrTLP10、BrTLP13的相對表達量均在2 h時最高,包括BrTLP5在內(nèi)均呈現(xiàn)先升后降的趨勢;BrTLP1、BrTLP3、BrTLP8、BrTLP9、BrTLP11、BrTLP12相對表達量較低;BrTLP14在鹽脅迫12h時表達量達到最高。

圖9 鹽脅迫下白菜TLP基因家族表達分析
3 討論與結論
在動植物生長發(fā)育過程中,TLP家族在維持細胞正常生理功能及應對脅迫過程中發(fā)揮了重要的調(diào)控作用。本研究通過雙向Blast在白菜基因組中初步完成了白菜TLP家族成員的鑒定,并對其系統(tǒng)進化關系、保守性及表達特性進行了初步分析,為后續(xù)探究白菜中TLP基因功能奠定了基礎。
白菜與擬南芥在進化上具有較近的祖先,而白菜經(jīng)歷的全基因組三倍化(WGT)事件導致了其與擬南芥在基因組層面上的一些差異。根據(jù)WGT理論,白菜TLP基因數(shù)量應為擬南芥的三倍左右[24],而本研究結果表明BrTLP家族數(shù)量并未出現(xiàn)顯著增加。在其他幾種植物中發(fā)現(xiàn)的TLP基因家族成員數(shù)量均為十幾個,表明在不同物種之間具有高度的保守性,白菜與擬南芥TLP基因家族的共線性分析也證明了兩個物種間顯著的共線性關系。在動物中的研究表明盡管Tubby結構域高度保守,但不同TLP基因成員在功能上卻無法相互補償[25],推測在進化過程中TLP家族可能丟棄了部分冗余基因,僅保留了現(xiàn)有的少數(shù)成員高度參與眾多真核生物生長發(fā)育中的調(diào)控過程。
基因結構與蛋白序列的解析為相關基因生物學功能的研究奠定了基礎。對白菜TLP基因家族結構及保守基序進行分析表明,白菜TLP家族在進化上距離較近成員之間的基因結構具有一定相似性,而與距離較遠的成員基因結構存在一定差異,暗示了家族成員在生物學功能上可能存在不同。另外,保守基序分析表明白菜TLP家族成員雖然在基因結構上存在一定差異,但部分序列仍具有很強的保守性。非保守區(qū)段氨基酸序列的變化導致了白菜TLP家族在多物種TLP家族進化樹中的不均勻分布,這也進一步暗示了該家族不同成員間的生物學功能可能存在較大差異,而在物種間進化距離較近的TLP成員在功能上可能高度保守。
以往研究證明TLP基因家族在植物應對非生物脅迫過程中發(fā)揮了重要調(diào)控作用。本研究發(fā)現(xiàn)絕大多數(shù)白菜TLP成員均對鹽脅迫有不同程度的響應,其中BrTLP10在鹽脅迫發(fā)生后的4~12 h表達明顯降低,對鹽脅迫的響應十分顯著。研究表明,擬南芥中的基因AtTLP9在響應鹽脅迫及干旱脅迫過程中具有重要作用[13],多物種TLP進化分析的結果表明BrTLP10與AtTLP9在進化上距離最近,另外,在白菜與擬南芥的共線性分析中也表明這兩個成員存在一定共線性關系[10]。根據(jù)AtTLP10現(xiàn)有的研究推測BrTLP9不僅參與鹽脅迫響應,可能也在白菜應對干旱脅迫過程中具有重要作用,但其在鹽脅迫下的具體調(diào)控機理以及是否在白菜應對干旱脅迫的過程中發(fā)揮作用還有待進一步研究。
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(2016年15期)2016-12-01 05:31:22
當代經(jīng)濟研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫(yī)藥現(xiàn)代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
