綠萼鳳仙花MYB4的克隆及表達(dá)分析
2023-03-06 07:08:58李澤鳳向南星魏春梅李新藝孟丹晨黃海泉黃美娟
山東農(nóng)業(yè)科學(xué) 2023年1期
李澤鳳向南星魏春梅李新藝孟丹晨黃海泉黃美娟
(西南林業(yè)大學(xué)園林園藝學(xué)院/國家林業(yè)和草原局西南風(fēng)景園林工程技術(shù)工程研究中心/云南省功能性花卉資源及產(chǎn)業(yè)化技術(shù)工程研究中心/西南林業(yè)大學(xué)園林園藝花卉研發(fā)中心,云南 昆明 650224)
植物木質(zhì)化是指木質(zhì)素形成并沉積在細(xì)胞壁中的一種生物學(xué)過程[1,2],一般在次生細(xì)胞壁中發(fā)生,且木質(zhì)素與植物的進(jìn)化歷程緊密相關(guān),這可能對植物從水生環(huán)境登陸到陸地環(huán)境生活發(fā)揮著關(guān)鍵作用[3]。較高的木質(zhì)化程度不僅可以提高植物的抗旱性[4],而且能提高植物的觀賞價值。木質(zhì)素是細(xì)胞壁的重要組分之一,主要在細(xì)胞次生壁中沉積,如導(dǎo)管、纖維和表皮等[5],具有硬度強、不溶水性以及不易被降解的化學(xué)特性,在植物生長和發(fā)育過程中起機械支撐作用,也是植物防御的屏障和水分運輸所必需的[6-8]。關(guān)于木質(zhì)素的合成調(diào)控已有研究表明,MYB46可直接對木質(zhì)素途徑進(jìn)行調(diào)控[9];小麥中的TaMYB[10]以及玉米中的ZmMYB46和ZmMYB31等[11]基因參與了調(diào)控木質(zhì)素的生物合成;火炬松(Pinus taedaL.)中的MYB4[12]能與木質(zhì)素合成基因的啟動子區(qū)的AC元件結(jié)合;桉樹(Eucalyptus grandis)中的MYB2會下調(diào)結(jié)構(gòu)基因CCR的表達(dá)[13],推測會抑制木質(zhì)素的生物合成;除此之外,AtWRKY12可與NST2的啟動子區(qū)域結(jié)合,進(jìn)而負(fù)調(diào)控植物細(xì)胞中木質(zhì)素的表達(dá)[14]。MYB、NAC和WRKY類轉(zhuǎn)錄因子都參與了木質(zhì)素的表達(dá)調(diào)控,但尤以MYB轉(zhuǎn)錄因子報道得最多。
按照MYB結(jié)構(gòu)域數(shù)目,MYB類轉(zhuǎn)錄因子被分為R1-MYB、R2R3-MYB、R1R2R3-MYB以及4R的MYB[15]。已有眾多研究表明MYB4的N端具有特定的R2、R3結(jié)構(gòu)域,因此可以推測其屬于R2R3-MYB轉(zhuǎn)錄因子。在木質(zhì)素合成調(diào)控網(wǎng)絡(luò)中,郭亞玉等[16]認(rèn)為SND1S作用于MYB轉(zhuǎn)錄因子,其中MYB轉(zhuǎn)錄因子不僅可以作為轉(zhuǎn)錄激活因子,還可以作為轉(zhuǎn)錄抑制因子來調(diào)控木質(zhì)素的合成。MYB4轉(zhuǎn)錄因子通過參與木質(zhì)素的調(diào)控過程而影響植物木質(zhì)部細(xì)胞壁的加厚,如在轉(zhuǎn)BpMYB4基因擬南芥(Arabidopsis thaliana)植株中木質(zhì)素含量降低,因此BpMYB4可作為木質(zhì)素生物合成的抑制因子[17];Shen等[18]研究表明,在煙草和柳枝稷中過表達(dá)PvMYB4基因,木質(zhì)素含量顯著減少,從而影響其生長發(fā)育質(zhì)量;擬南芥的MYB4轉(zhuǎn)錄因子會抑制木質(zhì)素的合成,其同源基因MYB32和MYB7則負(fù)調(diào)控木質(zhì)素的相關(guān)合成途徑[19,20]。由此可見MYB4轉(zhuǎn)錄因子在植物中的過表達(dá)可能負(fù)調(diào)控植物木質(zhì)素的合成,進(jìn)而影響植物的木質(zhì)化程度及次生細(xì)胞壁加厚。
鳳仙花屬植物具有經(jīng)濟(jì)、藥用、觀賞和園藝價值[21]。綠萼鳳仙花(Impatiens chlorosepala)是鳳仙花科(Balsaminaceae)鳳仙花屬(Impatiens)的一年或多年生植物,生于山谷水旁陰處或疏林溪旁,其花形奇特,花淡黃色或橘黃色,低矮,分枝能力強,莖肉質(zhì)實心。綠萼鳳仙花觀賞價值極高,可以作為園林地被植物予以開發(fā)。木質(zhì)素含量能影響植株木質(zhì)化程度,木質(zhì)化程度高的植物株型挺拔、抗逆性強、觀賞價值高。迄今為止,國內(nèi)外尚未見有關(guān)綠萼鳳仙花木質(zhì)素調(diào)控基因MYB4的相關(guān)報道。
本研究克隆了綠萼鳳仙花MYB4基因,并對其序列進(jìn)行相關(guān)分析;在此基礎(chǔ)上,采用qRTPCR探究其在綠萼鳳仙花不同部位的表達(dá)模式,進(jìn)而探究MYB4基因在木質(zhì)素合成過程中的功能和作用,以期為進(jìn)一步研究綠萼鳳仙花中MYB4基因在木質(zhì)素合成中的分子機制提供理論支持,并為后期增加鳳仙花木質(zhì)化程度及品種改良提供基礎(chǔ)數(shù)據(jù)。
1 材料與方法
1.1 試驗材料
綠萼鳳仙花于2020年7月種植于西南林業(yè)大學(xué)樹木園溫室大棚,2021年10月取其生長健壯的根尖、地上5 cm的莖基部及從上往下第四片成熟葉。
試劑材料:RNA提取試劑盒(OMEGA)、凝膠回收試劑盒(昆明碩陽科技有限公司)、逆轉(zhuǎn)錄試劑盒(昆明云科生物技術(shù)有限公司)、DH5α菌株(TAKARA)、pMD19-T載體(TAKARA)等。引物由生工生物工程(上海)股份有限公司合成。
1.2 試驗方法
1.2.1 總RNA的提取與MYB4基因的克隆 根據(jù)RNA提取試劑盒說明書提取綠萼鳳仙花根、莖和葉的總RNA;采用逆轉(zhuǎn)錄試劑盒將RNA逆轉(zhuǎn)錄成cDNA,-20℃保存?zhèn)溆谩J褂谜n題組已有的轉(zhuǎn)錄組數(shù)據(jù)對綠萼鳳仙花MYB4基因設(shè)計特異性引物,詳見表1。以逆轉(zhuǎn)錄后的cDNA第一鏈為模板進(jìn)行擴增,擴增體系(50μL):10×Easy TaqBuffer(Mg2+)6μL,dNTP 4μL,Easy TaqDNA Polymerase 0.5μL,上下游引物各2.5μL,cDNA 2.5 μL,ddH2O 32μL。擴增程序:95℃5 min;95℃50 s,55℃1 min,72℃1 min,35個循環(huán);72℃10 min。對目的片段進(jìn)行回收并純化后克隆連接到pMD19-T載體上,轉(zhuǎn)化大腸桿菌DH5α感受態(tài)細(xì)胞后,挑選陽性克隆進(jìn)行驗證、測序。
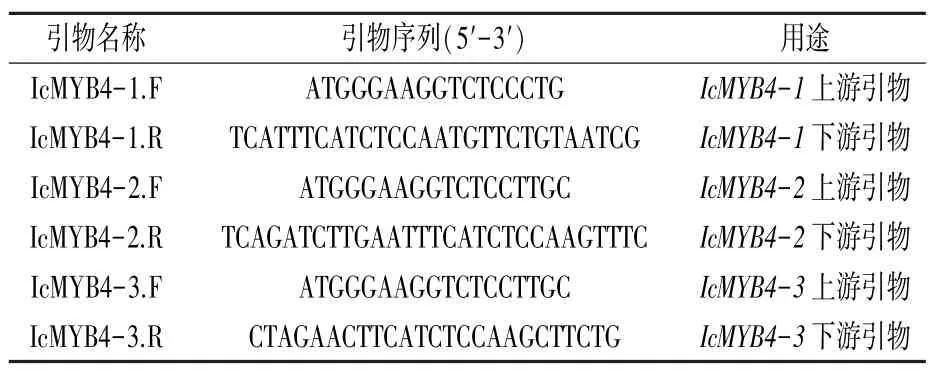
表1 綠萼鳳仙花MYB4基因cDNA序列擴增引物
1.2.2MYB4基因序列分析 運用ExPasy-Prot-Param軟件對MYB4基因編碼的蛋白質(zhì)進(jìn)行理化性質(zhì)分析;利用CDD工具對目的基因的結(jié)構(gòu)域進(jìn)行預(yù)測;使用BLAST功能找出與綠萼鳳仙花MYB4基因同源性相對較高的其他植物的氨基酸序列,通過DNAMAN軟件完成相關(guān)序列的同源性分析,并使用MEGA 7.0構(gòu)建系統(tǒng)進(jìn)化樹。
1.2.3MYB4基因表達(dá)模式分析 將綠萼鳳仙花根、莖和葉3個部位的RNA逆轉(zhuǎn)錄為cDNA并稀釋10倍備用。設(shè)計qRT-PCR引物(表2),qRTPCR擴增體系(20μL):qPCR SYBR Green Master Mix 10μL,ddH2O 8.2μL,上下游引物各0.4μL,cDNA 1μL。擴增程序:95℃5 min;95℃15 s,60℃30 s,72℃1 min,40個循環(huán);95℃15 s,60℃1 min,95℃15 s。內(nèi)參基因為IcActin,每個樣品3個重復(fù),并將根定義為1個單位作為對照,采用2-△△Ct對目的基因在根、莖和葉3個部位的相對表達(dá)量進(jìn)行計算,利用SPSS對其進(jìn)行顯著性分析。

表2 綠萼鳳仙花MYB4基因qRT-PCR擴增引物
2 結(jié)果與分析
2.1 綠萼鳳仙花MYB4基因的克隆
通過PCR擴增,獲得IcMYB4-1、IcMYB4-2和IcMYB4-3基因全長cDNA序列,分別為666、888、771 bp(圖1),編碼221、295、256 aa。將其cDNA序列與基因組gDNA比較發(fā)現(xiàn),IcMYB4-1、IcMYB4-2和IcMYB4-3基因組序列均在ATG下游263 bp處含有長度分別為83、107、117 bp的內(nèi)含子。
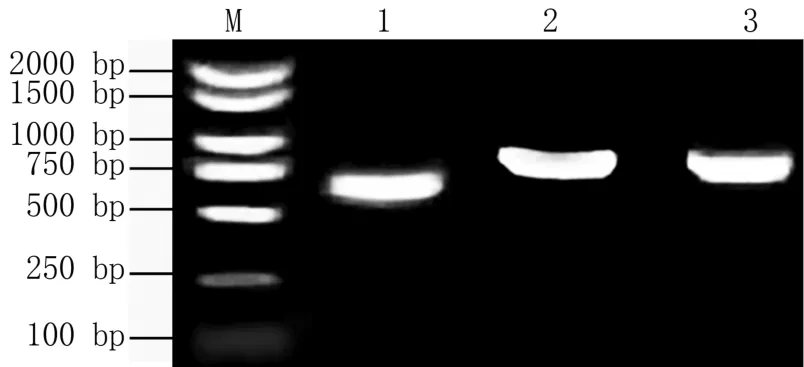
圖1 綠萼鳳仙花MYB4基因PCR擴增結(jié)果
2.2 綠萼鳳仙花MYB4基因的序列結(jié)構(gòu)分析
綠萼鳳仙花MYB4基因編碼蛋白質(zhì)的基本理化性質(zhì)分析結(jié)果(表3)表明,IcMYB4-1、IcMYB4-2和IcMYB4-3的不穩(wěn)定指數(shù)均高于40,為不穩(wěn)定蛋白,平均親水指數(shù)為負(fù)值,屬于親水性蛋白。通過CDD工具分析結(jié)果表明,3個MYB4蛋白均屬于MYB超家族蛋白成員,具特有的SANT結(jié)構(gòu)域和穩(wěn)定的PLN03091結(jié)構(gòu)域(圖2)。
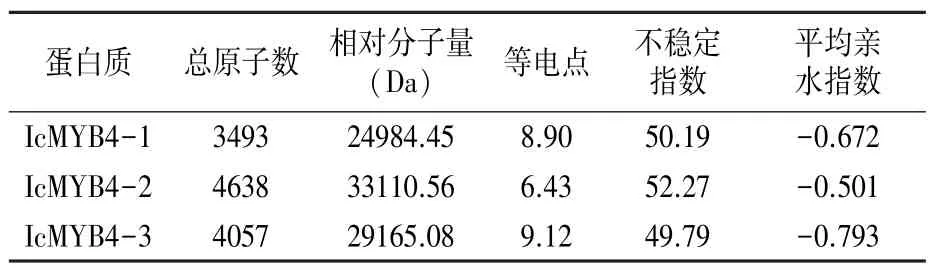
表3 綠萼鳳仙花MYB4蛋白質(zhì)的理化性質(zhì)
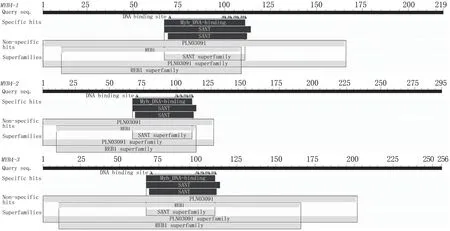
圖2 綠萼鳳仙花MYB4蛋白質(zhì)的保守結(jié)構(gòu)域
2.3 綠萼鳳仙花MYB4基因的同源性及系統(tǒng)進(jìn)化分析
對綠萼鳳仙花及其他物種MYB4基因的氨基酸序列進(jìn)行分析,結(jié)果表明,IcMYB4-1、IcMYB4-2和IcMYB4-3與其他物種的相似性在62%左右(圖3)。N端存在保守的R2和R3 DNA結(jié)合結(jié)構(gòu)域,同源性較高;C端同源性相對較低,且同時具有抑制型的保守基序EAR。系統(tǒng)親緣關(guān)系結(jié)果(圖4)表明,進(jìn)化樹分為兩大分支,IcMYB4-1和IcMYB4-2處于一個分支,推測其可能為直系同源關(guān)系,而IcMYB4-3與前兩者分屬兩個分支,推測其可能為旁系同源關(guān)系。
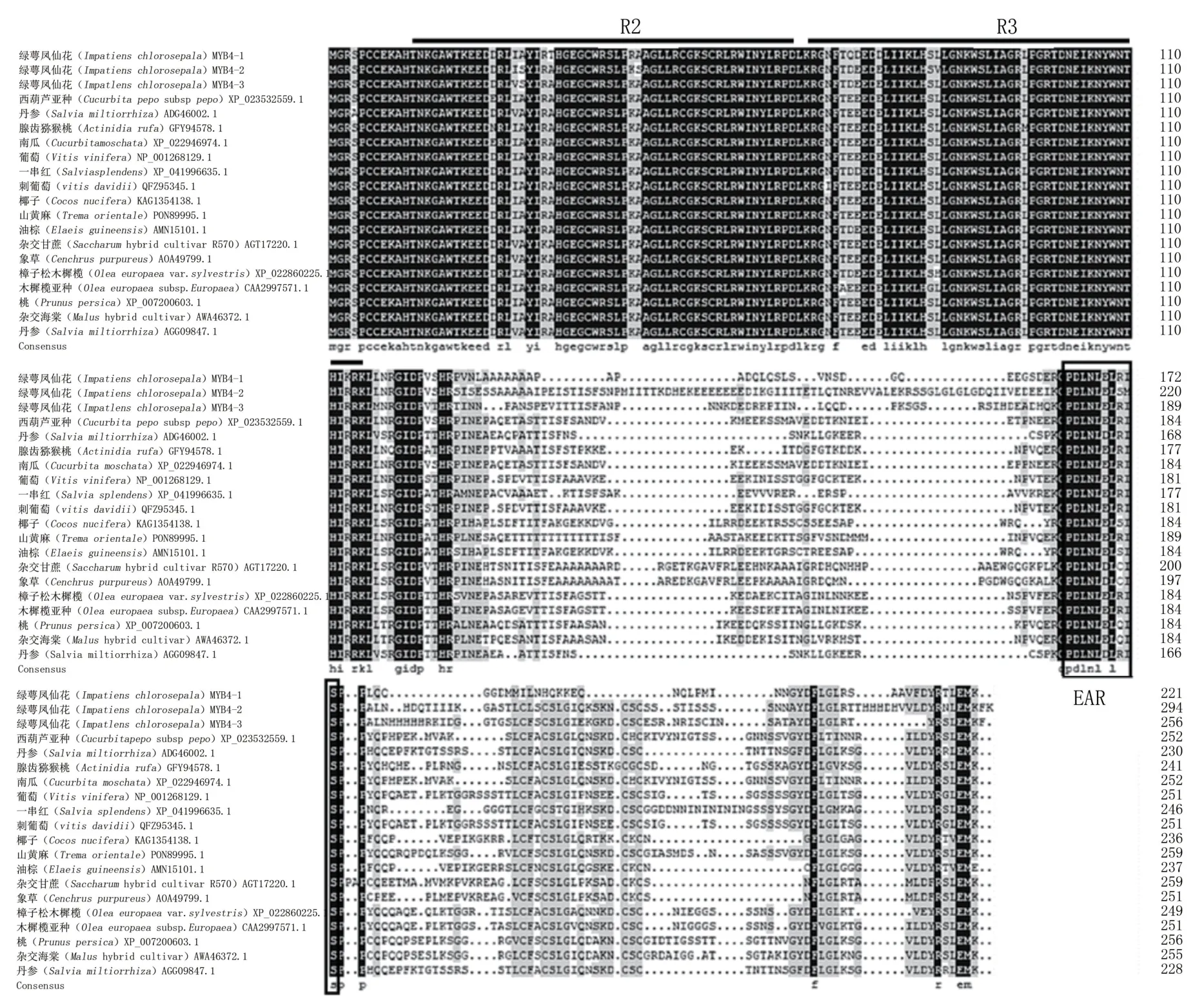
圖3 MYB4基因的同源氨基酸序列比對
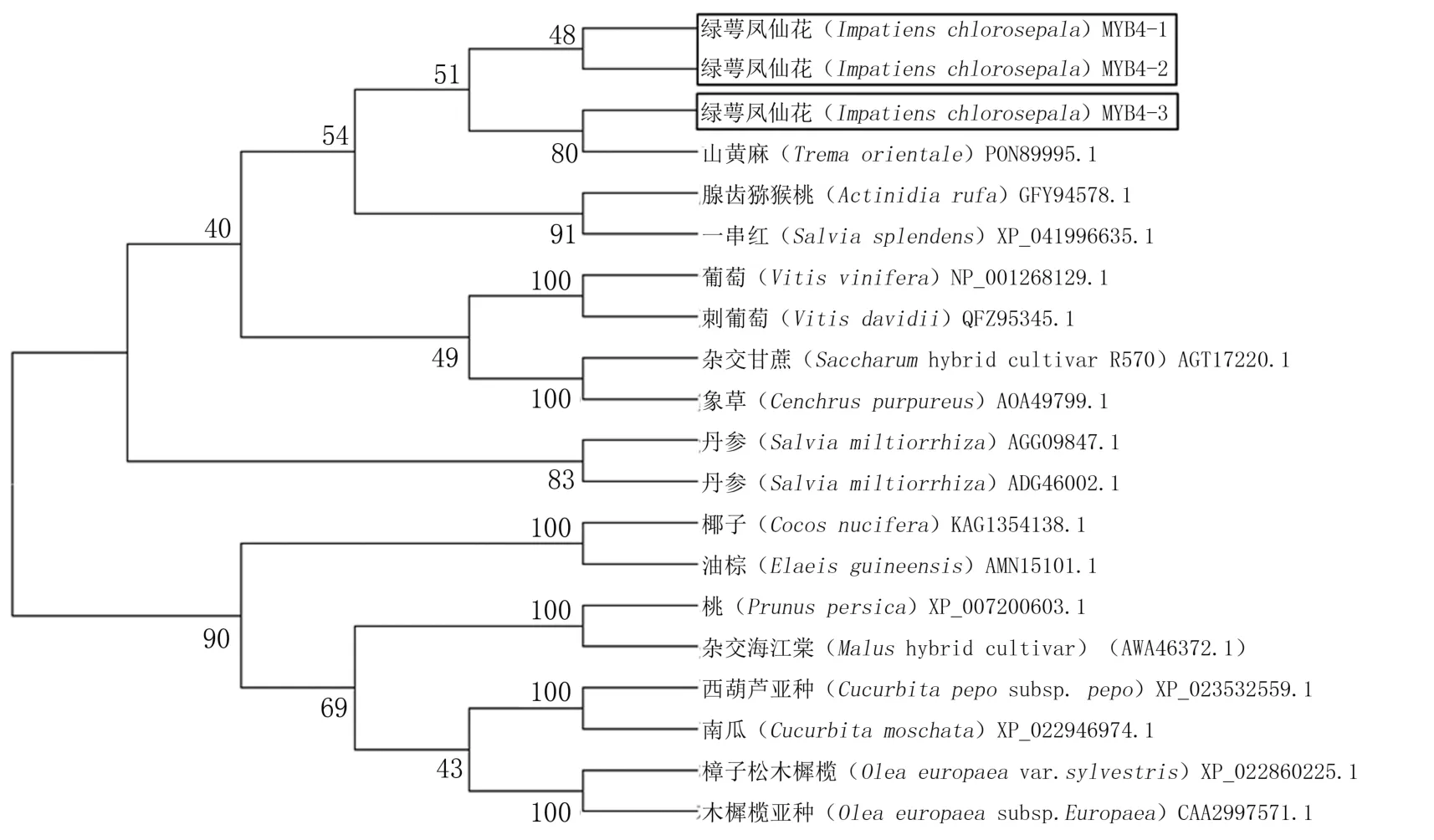
圖4 基于綠萼鳳仙花MYB4基因氨基酸序列構(gòu)建的系統(tǒng)發(fā)育樹
2.4 綠萼鳳仙花MYB4基因的表達(dá)分析
研究發(fā)現(xiàn),3個IcMYB4基因在綠萼鳳仙花的3個不同部位(根、莖和葉)均有表達(dá)(圖5)。以根為參照,IcMYB4-1在綠萼鳳仙花不同部位的表達(dá)均為最低,表現(xiàn)為在根中最多,葉次之,莖最少;IcMYB4-2和IcMYB4-3基因在綠萼鳳仙花不同部位的表達(dá)趨勢一致,表現(xiàn)為在根、莖和葉中依次升高。在綠萼鳳仙花的3個不同部位中,根木質(zhì)化程度相對較高,含木質(zhì)素較多,而葉含木質(zhì)素較少,推測IcMYB4-1基因可能對綠萼鳳仙花木質(zhì)素合成起正調(diào)控作用,而IcMYB4-2和IcMYB4-3對綠萼鳳仙花的木質(zhì)素合成起負(fù)調(diào)控作用。
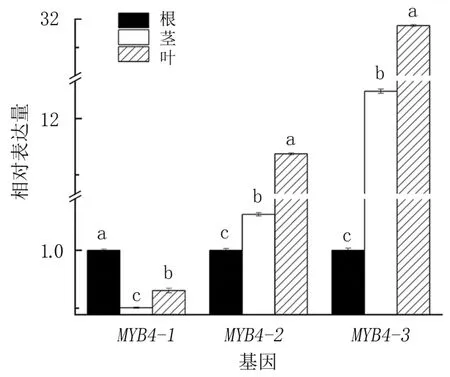
圖5 MYB4基因在綠萼鳳仙花3個不同部位的表達(dá)模式分析
3 討論與結(jié)論
MYB轉(zhuǎn)錄因子參與了植物的整個生長過程及其響應(yīng)生物和非生物脅迫的過程[22]。在MYB家族中,R2R3-MYB蛋白在木質(zhì)素合成及次生壁增厚等方面發(fā)揮著重要作用[23,24]。本研究從綠萼鳳仙花中成功克隆了木質(zhì)素合成相關(guān)轉(zhuǎn)錄因子IcMYB4-1、IcMYB4-2和IcMYB4-3,cDNA全長分別為666、888、771 bp,編碼221、295、256 aa,均屬于不穩(wěn)定親水性蛋白,與李嘉哲等[25]在山新楊中的研究結(jié)果一致;且都含有一個內(nèi)含子,與趙佳等[26]對月季花青素相關(guān)R2R3-MYB蛋白的研究結(jié)果一致。3個MYB4蛋白均屬于MYB超家族蛋白成員,含有MYB家族R2和R3的DNA結(jié)合結(jié)構(gòu)域,在N端同源性較高,C端差異較大,并且存在保守抑制型基序EAR[27],與已有研究結(jié)果一致[28];IcMYB4-1、IcMYB4-2和IcMYB4-3與其他物種MYB4基因的氨基酸序列同源性在62%左右。上述研究均證明在各物種之間MYB轉(zhuǎn)錄因子的DNA結(jié)構(gòu)域是高度保守的[29]。系統(tǒng)進(jìn)化分析表明IcMYB4-1和IcMYB4-2處于一個分支,推測其可能為直系同源關(guān)系;而IcMYB4-3與前兩者分屬不同分支,因此推測可能為旁系同源關(guān)系。
已有研究表明,MYB4基因在不同植物中的拷貝數(shù)不同,且在大多數(shù)植物的各個部位均有表達(dá);即使在同一物種,不同部位的表達(dá)量也具有顯著差異。例如在茶樹兩個MYB轉(zhuǎn)錄因子的克隆及功能驗證中,CsMYB4-5與CsMYB4-6都在根中表達(dá)量較高,在莖中表達(dá)量較低[30]。本研究分析發(fā)現(xiàn),IcMYB4-1、IcMYB4-2和IcMYB4-3在綠萼鳳仙花的3個部位均有表達(dá);其中,IcMYB4-1的表達(dá)量在根中最多,莖和葉的表達(dá)量較少,與李嘉哲等[25]的研究結(jié)果一致;IcMYB4-2和IcMYB4-3的表達(dá)趨勢一致,均為在葉中最多,莖次之,根最少,與對當(dāng)歸MYB4轉(zhuǎn)錄因子的研究結(jié)果一致[31]。以上結(jié)果表明,MYB4轉(zhuǎn)錄因子在不同植物甚至是同一植物的不同部位中可能發(fā)揮著不一樣的功能和作用。
有研究證明小麥的TaMYB4轉(zhuǎn)錄因子在煙草中異源過表達(dá)時,其總木質(zhì)素含量下降[10];象草PpMYB4基因在煙草中過表達(dá)時,轉(zhuǎn)基因煙草的木質(zhì)素含量顯著低于野生型[13]。這些研究表明MYB4基因在木質(zhì)素的合成調(diào)控中起到了抑制作用。在山新楊中PdPapMYB4在木質(zhì)化莖中大量表達(dá),遠(yuǎn)高于幼莖[25],說明MYB4基因可能在不同植物中對木質(zhì)素的調(diào)控作用不同。IcMYB4基因在綠萼鳳仙花的木質(zhì)素合成調(diào)控中起到了重要的作用,但其具體調(diào)控機制和功能還需進(jìn)一步驗證。
猜你喜歡
體育科技文獻(xiàn)通報(2022年3期)2022-05-23 13:46:54
天津外國語大學(xué)學(xué)報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設(shè)計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學(xué)報(2017年2期)2017-07-05 08:13:02
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
