甘薯IbbZIP22基因克隆與表達分析
2023-03-06 07:08:58孟鑫王慶美侯夫云李愛賢董順旭周媛媛秦楨張立明
山東農業科學 2023年1期
關鍵詞:分析
孟鑫王慶美侯夫云李愛賢董順旭周媛媛秦楨張立明
(1.青島農業大學農學院,山東 青島 266109;2.山東省農業科學院作物研究所/農業農村部黃淮海薯類科學觀測實驗站,山東 濟南 250100;3.山東省農業科學院,山東 濟南 250100;4.山東師范大學生命科學學院,山東 濟南 250014)
甘薯(Ipomoea batatasL.)為旋花科甘薯屬的藤本植物,是我國重要的糧食和經濟作物[1]。因具有高產穩產、適應性強、經濟效益好等諸多優勢,甘薯種植遍布世界100多個國家[2]。塊根作為甘薯的重要收獲器官,富含淀粉、維生素和礦質元素等營養物質[3],其中淀粉的含量和組成是甘薯產量和品質的主要決定因素[4]。
堿性亮氨酸拉鏈(bZIP)是家族成員眾多的一類轉錄因子,在真核生物轉錄因子中分布最廣泛、最保守[5]。bZIP轉錄因子家族具有一個bZIP結構域,由堿性區和亮氨酸拉鏈區兩部分組成,堿性區含有識別并結合啟動子上特定序列N-x7-R/K的基序;亮氨酸拉鏈區具有寡聚功能,行使轉錄激活或抑制功能。bZIP蛋白不僅參與植物的生長發育過程,而且參與響應逆境脅迫[6]。
有研究表明,bZIP轉錄因子可調控淀粉代謝途徑。ZmbZIP22基因過表達導致玉米和水稻中糖的含量、淀粉的結構和含量以及淀粉合成基因的表達水平發生變化[7]。bZIP基因參與木薯ABA信號通路且影響塊根淀粉積累[8]。然而bZIP轉錄因子在調節甘薯淀粉代謝途徑中的作用尚不清楚。本研究以濟薯25為材料,克隆得到IbbZIP22的基因序列及其啟動子序列,并對其含有的順式元件及在濟薯25和徐紫薯8號中的表達模式進行了分析,以期為深入研究IbbZIP22基因功能及進一步解析該基因調節甘薯淀粉代謝的機制提供依據。
1 材料與方法
1.1 試驗材料
試驗于2021年11月在山東省農業科學院作物研究所甘薯試驗基地進行。本研究用到的植物材料有甘薯栽培品種濟薯25(淀粉含量為22.6%)和徐紫薯8號(淀粉含量為18.0%)。將兩個甘薯品種的塊根種于大棚中的花盆內,重復3次,30 d后剪取長勢良好且大小一致、長度約為10 cm的甘薯嫩枝若干,在24℃恒溫室用Hoagland營養液進行水培,14 d后分別對塊根、須根、莖、葉取樣,經液氮快速冷凍,置于-80℃超低溫冰箱保存,用于后續表達分析。
1.2 試驗方法
1.2.1IbbZIP22基因克隆 使用Trizol法提取濟薯25塊根的總RNA,使用TaKaRa反轉錄試劑盒合成cDNA第一鏈。以野生甘薯(Ipomoea trifida)的ItZIP22基因CDS序列為模板,用Primer Premier 5軟件設計克隆引物(表1),以濟薯25的cDNA為模板,使用LA酶進行PCR擴增。PCR反應體系為20μL:10×Buffer 2μL,dNTPmix 1μL,正反向引物各0.5μL,cDNA模板1μL,LA酶0.2μL,ddH2O補足到20μL。反應程序為95℃預變性2 min;95℃變性30 s,55℃退火30 s,68℃延伸1 min,共35個循環;68℃延伸5 min。擴增后的PCR產物用1.5%瓊脂糖凝膠電泳檢測,并用DNA膠回收試劑盒(TIANGEN,北京)回收目的片段;將目的片段連接007VS-T載體并轉化到DH5α感受態中,挑選陽性克隆送青島擎科有限公司測序,保存測序正確菌液,完成克隆。
1.2.2IbbZIP22基因的生物信息學分析 使用NCBI ORF finder(http://www.ncbi.nlm.nih.gov/gorf/orfig.cgi)分析基因的ORF序列;利用NCBI的CDD數據庫(http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)分析IbbZIP22蛋白的保守結構域;利用ExPASy(http://web.expasy.org/protparam/)在線工具分析IbbZIP22蛋白的理化性質;利用SOPMA(http://pbil.ibcp.fr/)在線網站對IbbZIP22蛋白的二級結構進行預測;利用SWISS-MODEL(http://swissmodel.expasy.org/)對IbbZIP22蛋白的三級結構進行預測;利用Cell-PLoc 2.0(http://www.csbio.sjtu.edu.cn/bioinf/plantmulti/)對IbbZIP22蛋白進行亞細胞定位預測;在NCBI數據庫中選取不同物種的bZIP22同源蛋白,利用MEGA 7.0軟件構建系統發育進化樹。
1.2.3IbbZIP22基因在兩個甘薯品種不同部位的表達分析 根據獲得的IbbZIP22基因的全長cDNA序列,設計qRT-PCR引物(表1)。使用Trizol法分別提取濟薯25和徐紫薯8號的莖、葉、須根和塊根的總RNA并合成cDNA,以IbActin序列為內參,以兩個甘薯品種莖、葉、須根和塊根的cDNA為模板進行qRT-PCR反應,該反應在羅氏公司的LightCycler?480ⅡPCR儀上進行。每個樣品設置3次重復,基因相對表達量的計算采用2-ΔΔCt法。
1.2.4IbbZIP22基因的啟動子克隆與分析 根據IbbZIP22基因的cDNA序列,在I.trifida數據庫中預測IbbZIP22基因上游2 000 bp的序列作為啟動子擴增模板,利用濟薯25葉片基因組DNA、bZIP-pro-S1/A1引物(表1)擴增啟動子序列,連接純化后的擴增產物至007VS-T克隆載體并轉化大腸桿菌DH5α,陽性克隆送青島擎科生物技術有限公司測序,測序正確的序列利用PlantCARE在線進行順式作用元件預測和分析。

表1 引物序列
2 結果與分析
2.1 IbbZIP22基因的克隆
利用引物bZIP-S1和bZIP-A1,以濟薯25塊根cDNA為模板,通過PCR擴增及電泳檢測,得到1 400 bp左右的目的條帶(圖1)。經測序,得到大小為1 368 bp的IbbZIP22基因CDS序列(圖2)。
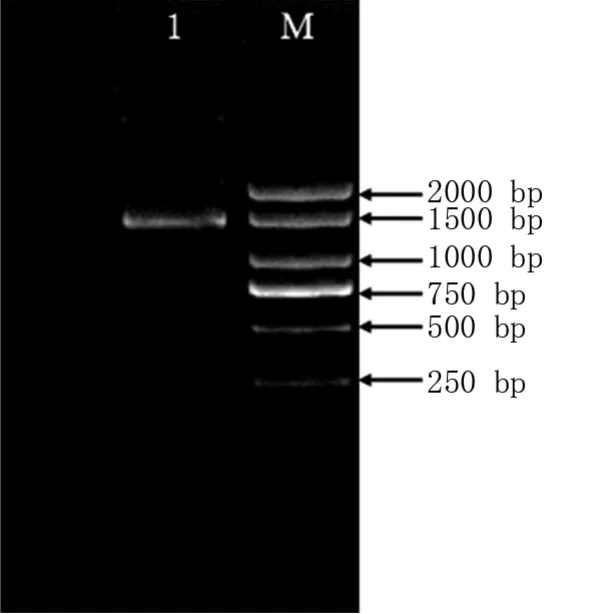
圖1 IbbZIP22基因的克隆
圖2 IbbZIP22基因cDNA序列和編碼氨基酸序列
2.2 IbbZIP22基因的生物信息學分析
經ORF finder分析,IbbZIP22基因開放閱讀框全長1 368 bp,編碼455個氨基酸(圖2)。利用NCBI分析保守結構域,發現該基因在第313~376位氨基酸殘基上有bZIP_plant_RF2保守結構域(圖3),推測其屬于RF2類bZIP轉錄因子。
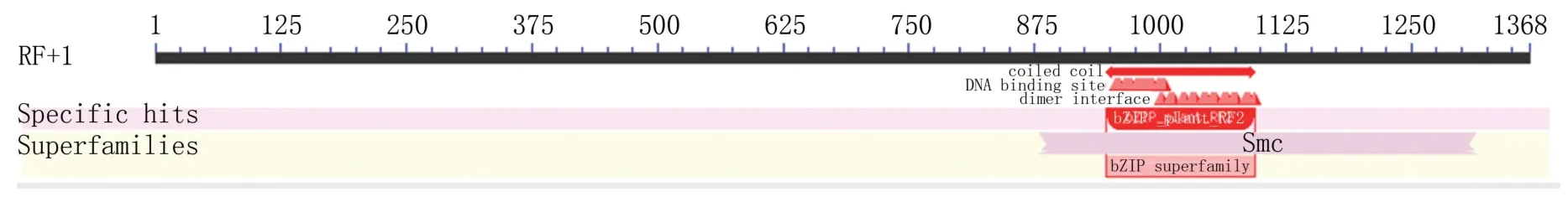
圖3 IbbZIP22基因的保守結構域分析
使用ExPASy在線分析IbbZIP22表達蛋白,推測該蛋白分子式為C2116H3402N632O702S24,相對分子量為49.6 kDa,其理論等電點為6.10,負電荷殘基總數和正電荷殘基總數分別為58和52;推測其半衰期為30 h,不穩定指數為56.65,脂肪族指數為65.21,親水性總平均值為-0.649,推測IbbZIP22蛋白為親水、不穩定蛋白。
通過SOPMA在線網站對IbbZIP22蛋白的二級結構進行預測,結果表明該蛋白含有α螺旋37.80%,β轉角1.54%,折疊延伸鏈3.30%和無規卷曲57.36%(圖4);通過SWISS-MODEL預測并構建IbbZIP22蛋白的三級結構模型(圖5);通過Cell-PLoc2.0對IbbZIP22蛋白進行亞細胞定位預測,預測該蛋白位于細胞核中。

圖5 IbbZIP22蛋白的三級結構預測
利用MEGA 7.0軟件構建甘薯(Ipomoea batatasL.)和煙草(Nicotianatabacum,XP_016451524)、可 可(Theobromacacao,XP_017971734)、木 薯(Manihot esculenta,XP_021610187)、三裂葉薯(Ipomoea triloba,XP_031125140)、牽牛(Ipomoea nil,XP_01916537)等16種植物基于bZIP同源蛋白的系統進化樹,結果(圖6)表明,甘薯的bZIP22蛋白與三裂葉薯的RF2a-like蛋白親緣關系最近,其次是煙草的bZIP19-like蛋白,與月季、山梨獼猴桃的bZIP蛋白親緣關系較遠,符合系統分類學特征。
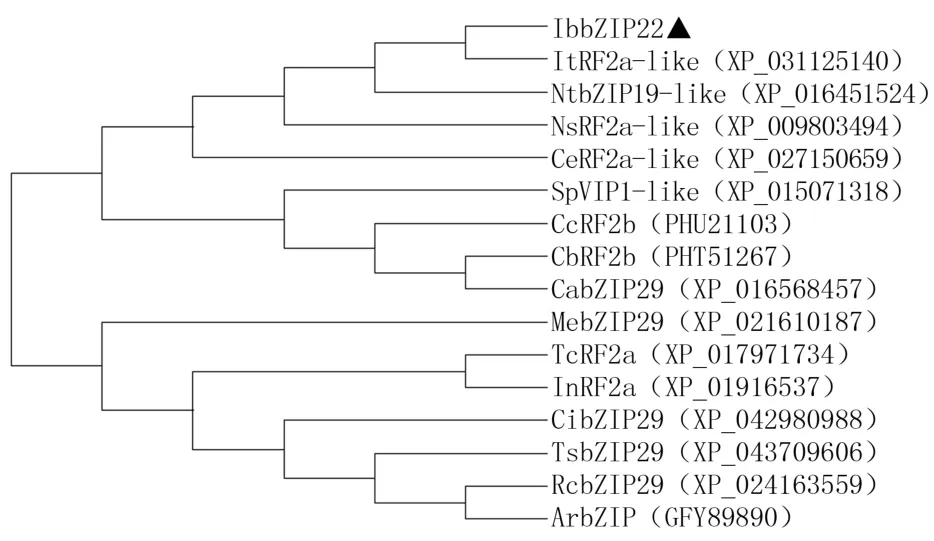
圖6 IbbZIP22蛋白與其他植物同源蛋白進化分析
2.3 IbbZIP22基因在兩個甘薯品種不同部位的表達分析
以IbActin作為內參基因,對濟薯25和徐紫薯8號不同部位中IbbZIP22基因的表達進行分析,結果(圖7)表明,IbbZIP22基因在兩個品種的葉、莖、須根和塊根中均有不同程度的表達,且均以塊根中的表達量最高,顯著高于在其他部位中的表達量。IbbZIP22基因在濟薯25須根中、在徐紫薯8號葉中的表達量最低,顯著低于其他部位。比較IbbZIP22在兩個品種塊根中的表達量(圖8)發現,IbbZIP22基因在濟薯25塊根中的表達量顯著高于徐紫薯8號,是徐紫薯8號的1.39倍。
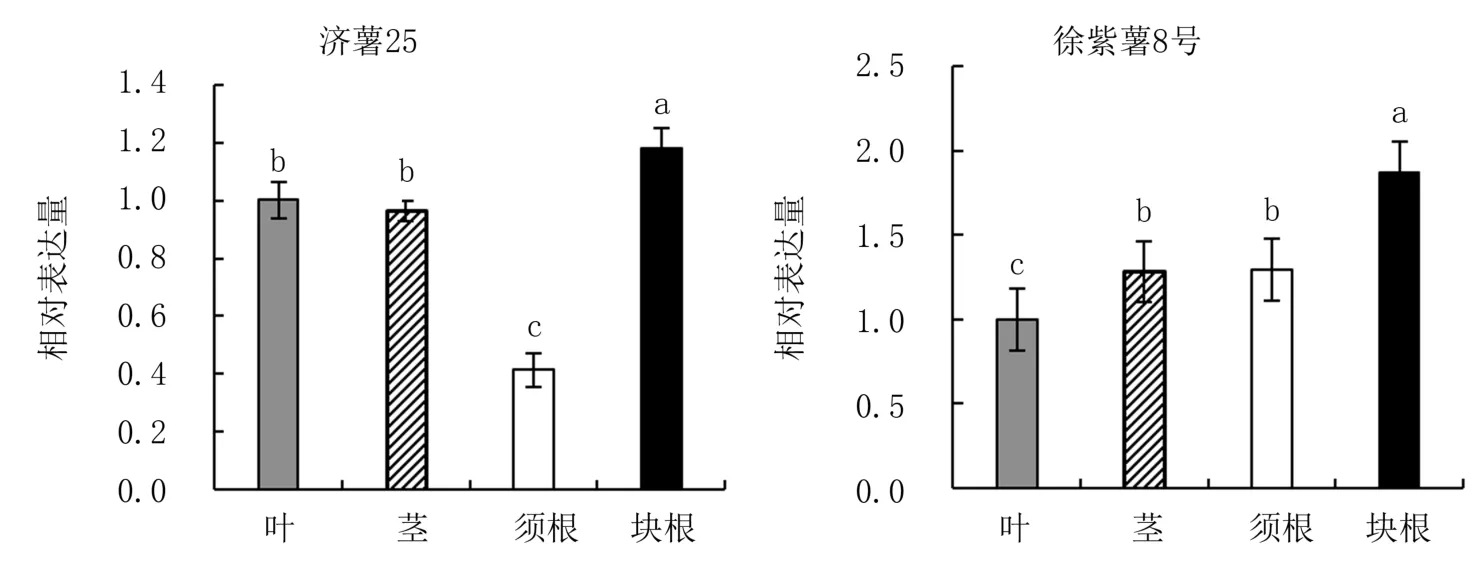
圖7 IbbZIP22基因在兩個甘薯品種中的表達模式分析
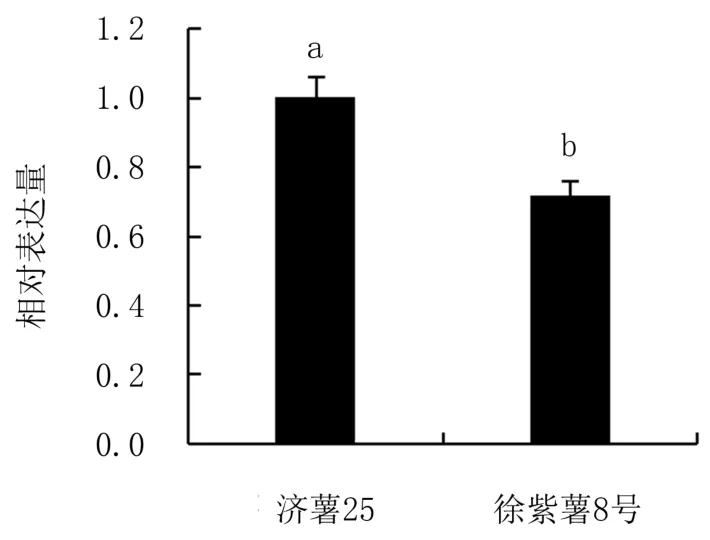
圖8 IbbZIP22基因在兩個甘薯品種塊根中的相對表達量
2.4 IbbZIP22基因的啟動子克隆與分析
以濟薯25基因組DNA為模板,通過PCR反應克隆得到長度為1 688 bp的啟動子片段(圖9)。利用PlantCARE在線軟件對IbbZIP22基因的啟動子進行預測,發現序列中除了含有TATAbox、CAAT-box等啟動子核心元件,還包括光響應元件(GT1-motif,TCT-motif,MRE,ACE)、干旱脅迫的MYB響應元件(MBS)、參與低溫響應的順式作用元件(LTR)、玉米醇溶蛋白代謝調控中的順式調控元件(O2-site)、厭氧誘導元件(ARE)及茉莉酸甲酯(TGACG-motif,CGTCA-motif)、脫落酸(ABRE)等激素相關的響應元件,以及一些功能未知的元件如MYB、MYC和STRE(表2)。
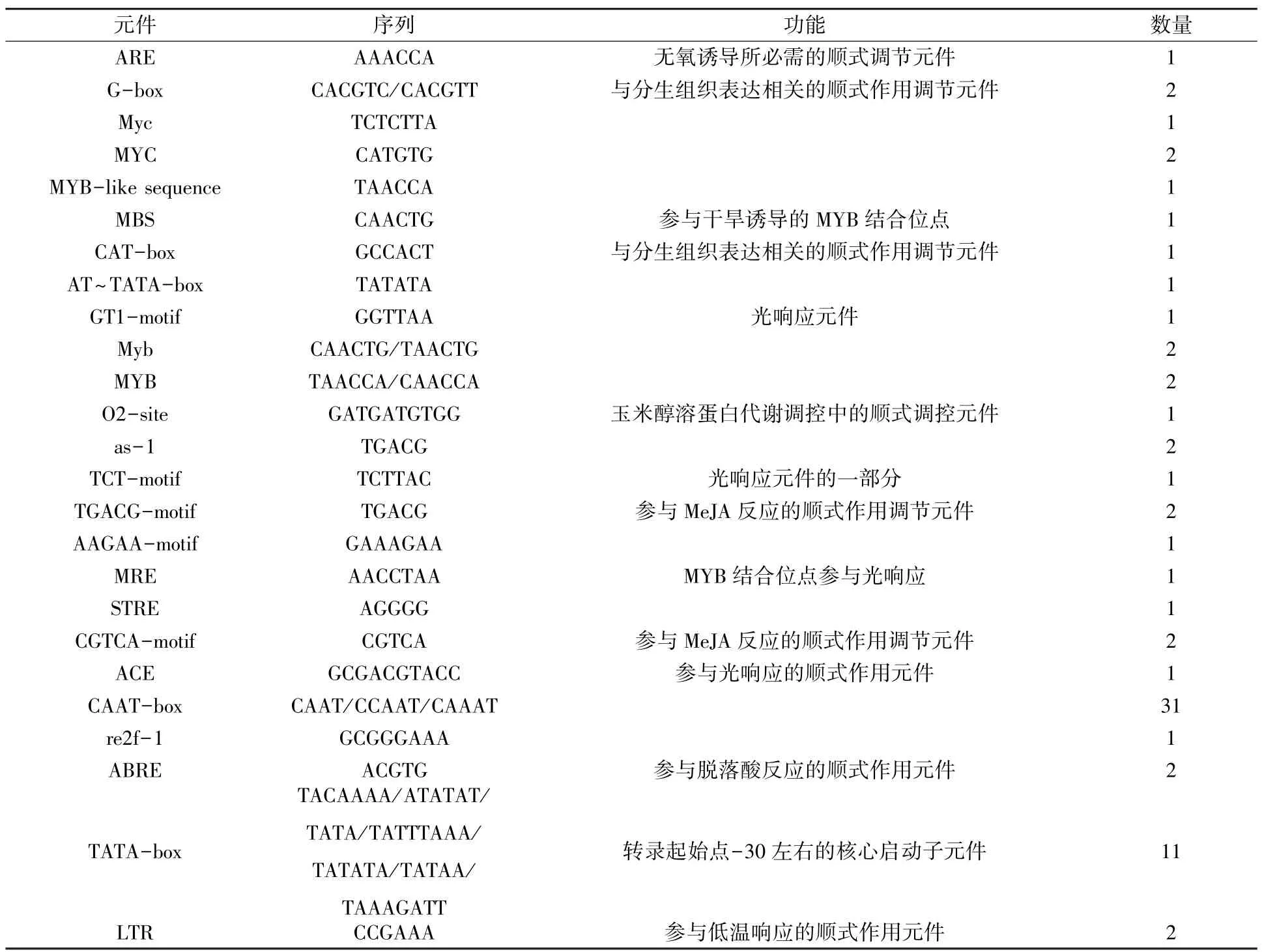
表2 IbbZIP22基因啟動子作用元件分析
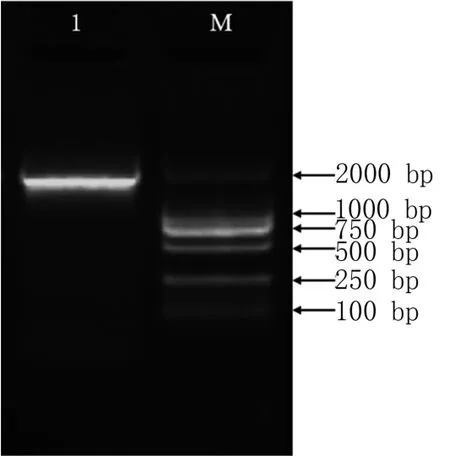
圖9 IbbZIP22基因啟動子的克隆
3 討論與結論
堿性亮氨酸拉鏈(bZIP)家族的轉錄因子是真核生物許多器官發育和生理過程的主要調節因子,包括形態發生、種子形成、非生物和生物脅迫反應[9,10]。本研究從濟薯25中克隆出IbbZIP22基因,根據保守結構域分析IbbZIP22蛋白含有bZIP_plant_RF2保守結構域,推測其屬于RF2類bZIP轉錄因子。系統進化樹分析結果表明,IbbZIP22蛋白與三裂葉薯的RF2a-like蛋白具有較高的同源性。
植物bZIP轉錄因子的研究進展大多與其參與非生物脅迫和發育有關,如種子成熟、花和維管發育、脅迫信號和病原體防御[11]。近年來,在小麥和玉米中bZIP轉錄因子參與淀粉合成的機理也逐漸被深入研究,結果表明,在過度表達TubZIP28的小麥中,成熟籽粒中的總淀粉含量升高,說明TubZIP28上調淀粉合成[12]。塊根是甘薯貯存營養的器官,淀粉在甘薯塊根中大量存在[13]。本研究選用濟薯25和徐紫薯8號兩個品種,對它們不同部位中IbbZIP22基因的表達進行分析,發現該基因在兩個品種塊根中均表達量最高。淀粉型品種濟薯25的淀粉含量為22.6%,比淀粉含量18.0%的徐紫薯8號略高,本研究發現,IbbZIP22基因在濟薯25號塊根中的表達量顯著高于徐紫薯8號,是徐紫薯8號的1.39倍,與這兩個品種淀粉含量的高低趨勢一致,因此推測IbbZIP22基因參與了淀粉的合成和代謝。
啟動子是基因表達調控的重要元件。本研究發現IbbZIP22基因啟動子上不僅包含核心元件TATA-box、CAAT-box等,還包含參與干旱誘導的MYB結合位點,推斷該基因可能受MYB轉錄因子調控。此外,IbbZIP22基因啟動子序列上還包含玉米醇溶蛋白代謝調控中的順式調控元件,有研究表明,bZIP型轉錄因子ZmbZIP22可以直接與27 kDγ-玉米醇溶蛋白啟動子結合,調節27 kDγ-玉米醇溶蛋白基因的表達[14],因此,在甘薯中是否可能存在相應的調控機制也值得被深入探討。該啟動子也含有參與干旱、低溫和光響應元件及與茉莉酸甲酯(MeJA)和脫落酸(ABA)反應的順式作用元件,說明IbbZIP22基因可能在甘薯非生物脅迫應答等過程中發揮作用,而且該基因的表達可能受MeJA和ABA等激素的調控。
本研究克隆了甘薯的IbbZIP22基因及其啟動子,并對該基因在不同淀粉含量品種中的表達進行了分析,可為探究甘薯抗逆和淀粉調控機制提供依據,并為進一步明確IbbZIP22基因的功能奠定基礎。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
