高耐性庫德里阿茲威氏畢赤酵母的篩選及應(yīng)用
2023-02-21 10:14:28林良才白茹高瀅慕濟(jì)鍺盧君李長文張翠英
食品與發(fā)酵工業(yè) 2023年3期
林良才,白茹,高瀅,慕濟(jì)鍺,盧君,李長文,張翠英*
1(天津科技大學(xué) 生物工程學(xué)院,工業(yè)發(fā)酵微生物教育部重點(diǎn)實(shí)驗(yàn)室,天津,300457) 2(貴州國臺酒業(yè)股份有限公司,貴州 仁懷,564501)
傳統(tǒng)白酒發(fā)酵是順其自然之道的開放式發(fā)酵,是多菌種的協(xié)同生態(tài)發(fā)酵過程。發(fā)酵體系中菌群的組成和代謝活性不僅決定了基酒的產(chǎn)量,還與基酒的品質(zhì)息息相關(guān)。然而,獨(dú)特的開放式發(fā)酵過程極易受到外界環(huán)境的影響,從而改變菌群結(jié)構(gòu),尤其是打破核心功能菌群之間的平衡,進(jìn)而出現(xiàn)原料轉(zhuǎn)化率低、風(fēng)味生成不足、有害副產(chǎn)物伴生等問題,如夏季較高的環(huán)境溫度極易造成乳酸、乙酸等有機(jī)酸的過度積累,導(dǎo)致糟醅酸度過高(pH<4),體系中功能酵母菌群的生長受到嚴(yán)重抑制,迫使企業(yè)“壓排”,甚至停產(chǎn)。特別是在醬香型白酒釀造過程中,因其多輪次的特殊工藝,夏季發(fā)酵的輪次糟醅酸度過高的問題尤為突出,嚴(yán)重影響出酒率和產(chǎn)品風(fēng)味。如何提高釀造微生態(tài)中功能菌群結(jié)構(gòu)的穩(wěn)定性和抗干擾性已成為企業(yè)穩(wěn)定、高效、安全生產(chǎn)亟需解決的行業(yè)共性技術(shù)問題。
酵母是白酒釀造過程中的核心功能微生物,是最為重要的乙醇生產(chǎn)菌株,也與風(fēng)味物質(zhì)的合成密切聯(lián)系,其抗逆性能和發(fā)酵性能不僅直接影響整個(gè)釀造微生態(tài)中菌群的演替變化,還與基酒產(chǎn)量和品質(zhì)息息相關(guān)。釀酒酵母(Saccharomycescerevisiae)被廣泛地視為白酒釀造過程中的主要發(fā)酵動力,是基酒中乙醇的主要來源,而發(fā)酵體系中非釀酒酵母因產(chǎn)醇能力相對較弱,往往被認(rèn)為與風(fēng)味物質(zhì)的合成密切相關(guān),如異常威克漢酵母屬和念珠酵母屬在醬香白酒第5輪次發(fā)酵過程中與乙酯含量呈正相關(guān)[1],粟酒裂殖酵母與包括丁醇、戊酸在內(nèi)的136種風(fēng)味物質(zhì)的合成相關(guān)[2],畢赤酵母屬和念珠酵母屬在小曲清香白酒釀造過程中參與高級醇的合成[3]。然而,隨著對傳統(tǒng)白酒釀造體系科學(xué)認(rèn)識的逐步深入,非釀酒酵母的功能已不僅僅局限于生香方面的貢獻(xiàn),它們是釀造核心功能微生物組中不可或缺的組成部分,尤其是這些非釀酒酵母對白酒發(fā)酵環(huán)境中的高溫、高酸、高滲等壓力脅迫具有較好的耐受性[4],對于穩(wěn)定發(fā)酵體系中菌群的組成和代謝活性具有不可替代的重要作用。因此,高耐性核心功能酵母菌株是應(yīng)對白酒發(fā)酵過程酸壓力脅迫下發(fā)酵力不足、保持釀造微生態(tài)平衡的有效工具。
庫德里阿茲威氏畢赤酵母(Pichiakudriavzevii)又稱東方伊薩酵母(Issatchenkiaorientalis),不僅具有較好的耐酸、耐高溫性能,而且在多重壓力脅迫下仍能保持優(yōu)良的發(fā)酵特性[5-6],是重要的釀造核心功能微生物,是醬香、濃香、芝麻香、清香型白酒發(fā)酵過程中的主要功能酵母[2-3,7-8],其代謝物苯乙醇可以顯著抑制發(fā)酵體系中其他真菌的生長,對釀造微生態(tài)中菌群演替具有調(diào)控作用[9]。P.kudriavzevii還具有較強(qiáng)的尿素降解能力,可以較好地降低醬香型白酒發(fā)酵體系中氨基甲酸乙酯(ethyl carbamate, EC)的前體物質(zhì)——尿素,從而降低EC含量,確保基酒的安全性[10]。此外,P.kudriavzevii可以降低發(fā)酵體系中乳酸的含量,調(diào)節(jié)釀造微環(huán)境的酸度,從而促進(jìn)釀酒酵母的生長[11]。盡管P.kudriavzevii在白酒釀造過程中發(fā)揮極為重要的作用,但將其人工應(yīng)用于白酒發(fā)酵過程的研究卻鮮有報(bào)道。因此,篩選具有良好耐受性能及發(fā)酵性能的庫德里阿茲威氏畢赤酵母,并開發(fā)基于強(qiáng)化菌劑的發(fā)酵調(diào)控技術(shù),對實(shí)現(xiàn)傳統(tǒng)白酒釀造可控、高效、穩(wěn)定生產(chǎn)具有重要的應(yīng)用價(jià)值。
本研究采用溫度、酸度雙重脅迫策略從醬香型白酒酒醅中特異性篩選、分離庫德里阿茲威氏畢赤酵母11株,對其耐高溫、耐酸、耐乙醇性能進(jìn)行檢測,并利用模擬發(fā)酵實(shí)驗(yàn)對其發(fā)酵性能及風(fēng)味物質(zhì)的生成進(jìn)行系統(tǒng)評估,最終確定以XDNZ_PK05菌株進(jìn)行原位發(fā)酵實(shí)驗(yàn),評價(jià)其應(yīng)用于醬香型白酒釀造過程中的作用,為傳統(tǒng)白酒人工可控發(fā)酵調(diào)控技術(shù)的開發(fā)提供了新依據(jù)和新思路。
1 材料與方法
1.1 材料與試劑
玉米,市售;酵母浸粉、蛋白胨,北京奧博星生物技術(shù)有限公司;葡萄糖(分析純)、NaCl,天津市永大化學(xué)試劑開發(fā)中心;瓊脂粉、瓊脂糖,北京索萊寶科技有限公司;乳酸、乙酸、乙醇,阿拉丁生化科技有限公司;dNTP、LATaqDNA聚合酶,日本TaKaRa公司;WL營養(yǎng)瓊脂培養(yǎng)基,青島海博生物技術(shù)有限公司。
1.2 培養(yǎng)基
YPD液體培養(yǎng)基(g/L):葡萄糖20,蛋白胨20,酵母浸粉10,pH自然,115 ℃滅菌20 min。固體培養(yǎng)基額外添加20 g/L瓊脂粉。
玉米水解液:稱取玉米醪1 kg,向其中加入3 L 60~70 ℃的自來水,靜置20 min,進(jìn)行玉米醪的糊化,使玉米醪充分膨脹,然后加入600 μL的液化酶(α-淀粉酶2×104U/mL),于85~90 ℃液化1.5 h,期間可進(jìn)行攪拌以提高液化效率,液化結(jié)束后,迅速冷卻降溫至60 ℃,加入2 mL糖化酶(1×105U/mL)糖化20 h,期間攪拌。待糖化結(jié)束后,將糖化液用3層濾布過濾,得到滲出液即為玉米水解液,在105 ℃下高壓滅菌20 min,室溫保存使用。
一級種子培養(yǎng)基:將玉米水解液的糖度調(diào)整到8°Brix,加入0.5%(質(zhì)量分?jǐn)?shù),下同)的酵母浸粉,按需求分裝滅菌備用。
二級種子培養(yǎng)基:將玉米水解液的糖度調(diào)整到12°Brix,加入0.5%的酵母浸粉,按需求分裝滅菌備用。
營養(yǎng)鹽:分別稱取15 g MgSO4,7.5 g KH2PO4,8.1 g尿素,依次加入燒杯中,加入80 mL蒸餾水,充分溶解,定容至100 mL。
濃醪發(fā)酵培養(yǎng)基:稱取60 g玉米醪置于250 mL三角瓶,加入135 mL 60~70 ℃熱水并攪拌均勻,放置20 min。向其中加入30 μL的液化酶(α-淀粉酶2×104U/mL)并攪拌均勻,90 ℃水浴90 min,液化。迅速冷卻至60 ℃,加入90 μL糖化酶(1×105U/mL)并攪拌均勻,60 ℃水浴20 min,糖化。迅速冷卻至40 ℃,加入1.2 mL酸性蛋白酶(2×104U/mL)并攪拌均勻,40 ℃水浴20 min。迅速冷卻至30 ℃,加入1 mL營養(yǎng)鹽溶液,即為發(fā)酵培養(yǎng)基。
1.3 儀器與設(shè)備
UVmini-1240紫外分光光度計(jì),島津儀器(蘇州)有限公司;7890A氣相色譜儀,美國安捷倫科技公司;YXQ-LS-30SH高壓蒸汽滅菌鍋,山東新華醫(yī)療器械廠;全自動凝膠成像儀,美國SYNGENE公司;PCT-200型PCR基因擴(kuò)增儀,德國Eppendorf公司;PowerPacTM型電泳儀,美國BIO-RAD公司;LRH-250A生化培養(yǎng)箱,上海博訊實(shí)業(yè)有限公司;小型蒸酒設(shè)備,河南永康機(jī)械制造有限公司。
1.4 實(shí)驗(yàn)方法
1.4.1 高耐受性酵母的篩選
將10 g醬香型白酒酒醅和90 mL無菌生理鹽水置于250 mL無菌三角瓶中充分混合,在30 ℃,180 r/min條件下振蕩1 h,吸取1 mL菌懸液至YPD培養(yǎng)基中,35 ℃靜止培養(yǎng)48 h,稀釋涂布于含有40 g/L乳酸的YPD平板上,35 ℃倒置培養(yǎng)2~3 d,挑取單菌落進(jìn)行純化,并保存于4 ℃?zhèn)溆谩?/p>
1.4.2 高耐受性酵母菌株的鑒定
(1)形態(tài)觀察
將純化后的菌株劃線接種至WL營養(yǎng)瓊脂培養(yǎng)基,30 ℃培養(yǎng)2~3 d后觀察并記錄菌落形態(tài),同時(shí)在顯微鏡下觀察其細(xì)胞形態(tài)。
(2)生理生化測定
參照楊舒雯等[12]的方法,對篩選出的菌株進(jìn)行生理生化試驗(yàn),包括糖發(fā)酵試驗(yàn)(主要測定葡萄糖、蔗糖、麥芽糖、可溶性淀粉)、碳源同化試驗(yàn)(主要測定果糖、半乳糖、木糖、阿拉伯糖)、氮源同化實(shí)驗(yàn)[主要測定KNO3、(NH4)2SO4、尿素]
(3)菌株分子生物學(xué)鑒定
利用真菌26S rDNA基因通用引物NL1(5′-GCATATCAATAAGCGGAGGAAAAG-3′)、NL4(5′-GGTCCGTGTTTCAAGACGG-3′)進(jìn)行PCR擴(kuò)增,條件為95 ℃預(yù)變性5 min;94 ℃變性45 s,57 ℃退火45 s,72 ℃延伸1 min,30個(gè)循環(huán);72 ℃ 10 min。反應(yīng)體系(20 μL):LATaq聚合酶0.125 μL,10×LA PCR Buffer 2 μL,dNTP Mix 1 μL,模板1 μL,引物NL1 1 μL,引物NL4 1 μL,ddH2O補(bǔ)齊至20 μL。PCR產(chǎn)物經(jīng)電泳驗(yàn)證,純化回收后由天津金唯智生物科技有限公司進(jìn)行測序分析。
將所獲得的PCR序列提交至NCBI進(jìn)行BLAST比對,獲取同源性較高的物種26S rDNA序列,利用MEGA 7.0中Neighbor-joining方法建立系統(tǒng)發(fā)育樹。
1.4.3 菌株耐性能評估
本實(shí)驗(yàn)采用點(diǎn)板實(shí)驗(yàn)法檢測菌株的耐受性,將測試菌株挑取至3 mL YPD培養(yǎng)基中,30 ℃培養(yǎng)12~16 h,將菌液OD600調(diào)整至0.5,吸取2 μL菌液分別滴加至含有不同質(zhì)量濃度乳酸(50、60、70 g/L)、乙酸(3、4、5、6 g/L)或乙醇(90、100、110 g/L)的YPD平板上,30 ℃倒置培養(yǎng)1~2 d,每隔12 h觀察菌株生長情況。類似的,利用點(diǎn)板法檢測菌株的耐高溫特性(37、42、45 ℃)。
1.4.4 發(fā)酵性能檢測
從平板挑取單菌落接種于含有5 mL一級種子培養(yǎng)基的試管中,30 ℃靜置培養(yǎng)24 h;將上述菌液全部接種至裝有54 mL二級種子培養(yǎng)基的150 mL三角瓶中,30 ℃靜置培養(yǎng)16 h;以10%(體積分?jǐn)?shù))接種量將二級種子發(fā)酵液接種至玉米濃醪發(fā)酵培養(yǎng)基中,30 ℃靜置培養(yǎng),通過CO2排放量來監(jiān)測整個(gè)發(fā)酵過程的進(jìn)程,每隔12 h稱重1次,當(dāng)12 h失重≤0.1 g時(shí),表明發(fā)酵已經(jīng)結(jié)束。
1.4.5 原位發(fā)酵實(shí)驗(yàn)
從平板挑取單菌落接種于200 mL YPD培養(yǎng)基中,在30 ℃,200 r/min條件下培養(yǎng)48 h,將酵母菌劑以106CFU/g的接種量加入50 kg六輪次攤涼拌曲后的酒醅中,混合后將其平均裝入5個(gè)純棉紗布袋中,放置在不同位置,待堆積發(fā)酵結(jié)束后,取樣進(jìn)行理化檢測,然后將樣品進(jìn)行翻拌,模擬移堆操作,然后再分裝至紗布袋中,在窖池原位環(huán)境中繼續(xù)發(fā)酵。發(fā)酵結(jié)束后,取酒醅進(jìn)行理化指標(biāo)檢測。
1.4.6 乙醇及風(fēng)味物質(zhì)檢測
濃醪發(fā)酵代謝物檢測:發(fā)酵結(jié)束后,取100 mL濃醪發(fā)酵液與100 mL蒸餾水混合均勻,蒸餾并收集100 mL餾出液,并用0.22 μm濾膜過濾,分別利用液相法和氣相法測定乙醇含量和風(fēng)味物質(zhì)含量。
原位發(fā)酵酒醅代謝物檢測:取50 g酒醅與200 mL蒸餾水混合均勻,蒸餾并收集100 mL餾出液,并用0.22 μm濾膜過濾,分別利用液相法和氣相法測定乙醇含量和風(fēng)味物質(zhì)含量。
液相檢測條件:色譜柱為GH0830078H(300 mm×7.8 mm);檢測器為示差折光檢測器(refractive index detector, RID);流動相為5 mmol/L的H2SO4溶液,流速0.6 mL/min;檢測器溫度45 ℃,柱溫65 ℃,進(jìn)樣量20 μL。
氣相檢測條件:火焰離子化檢測器(flame ionization detector, FID),毛細(xì)管色譜柱HP-INNOWAX(50 m×320 μm×1.0 μm),載氣為純度99.99%的高純N2,分流比1∶10。進(jìn)樣口溫度200 ℃,檢測器溫度200 ℃,進(jìn)樣量1 μL。采用程序升溫,50 ℃保持8 min,升溫速率5 ℃/min,升溫至180 ℃,保持15 min,每個(gè)樣品處理時(shí)間43 min。
1.4.7 殘淀粉的測定
采用斐林試劑法[13]測定樣品中的殘余淀粉含量。
1.4.8 數(shù)據(jù)處理
每個(gè)試驗(yàn)重復(fù)3次,采用軟件GraphPad Prism 8.0.1繪制圖表,表示為平均值±標(biāo)準(zhǔn)差,其中差異顯著性通過單因素方差分析(ANOVA)進(jìn)行,置信度為95%(P<0.05)。
2 結(jié)果與分析
2.1 高耐性菌株的分離篩選
庫德里阿茲威氏畢赤酵母是傳統(tǒng)發(fā)酵中重要的參與者,是酒類釀造過程中重要的功能酵母[14],為了篩選獲得高耐性的菌株,以醬香型白酒酒醅出發(fā),根據(jù)庫德里阿茲威氏畢赤酵母的耐受特性,采用傳統(tǒng)平板培養(yǎng)法在35 ℃和40 g/L乳酸的雙壓力脅迫下特異性分離、篩選獲得11株酵母菌株。
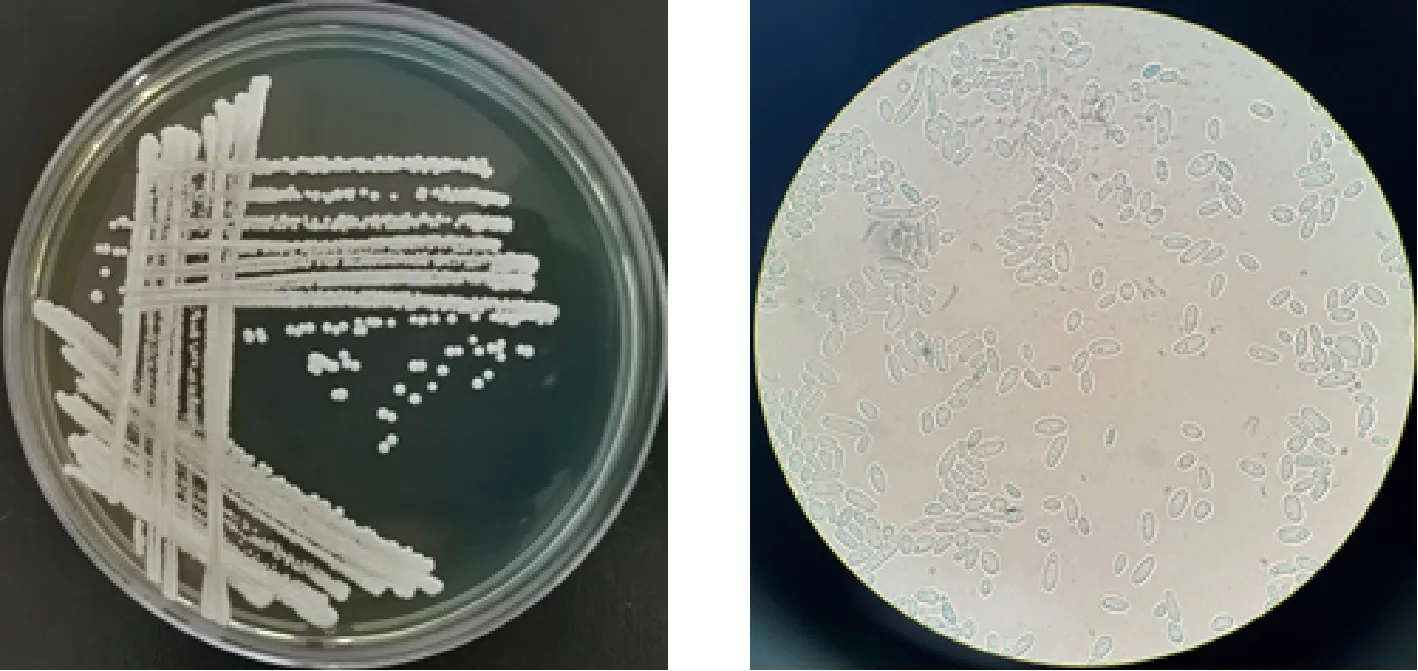
a-在WL營養(yǎng)瓊脂培養(yǎng)基上菌落形態(tài)特征;b-顯微鏡下細(xì)胞形態(tài)特征(10×100)圖1 菌株XDNZ_PK05的菌落及細(xì)胞形態(tài)Fig.1 Colony and cell morphology of strain XDNZ_PK05
對11株菌進(jìn)行形態(tài)鑒定時(shí)發(fā)現(xiàn)所有菌落形態(tài)一致,在此僅展示XDNZ_PK05的結(jié)果,如圖1所示。在WL營養(yǎng)瓊脂培養(yǎng)基觀察其形態(tài)特征,菌落呈圓形,顏色為乳白色,不透明,表面粗糙,凸起,表面及邊緣呈菌絲狀,四周平整;顯微鏡下觀察,大多為長橢圓形的單細(xì)胞,生殖方式為出芽繁殖,是典型的酵母特征。
生理生化試驗(yàn)結(jié)果如表1所示,11株菌的結(jié)果是一致的,可發(fā)酵的糖類為葡萄糖和蔗糖,不可發(fā)酵的糖類為蔗糖和淀粉;碳源同化試驗(yàn)顯示菌株可以利用果糖和半乳糖,不能利用木糖和阿拉伯糖;氮源同化試驗(yàn)顯示菌株均不能利用尿素、(NH4)2SO4、KNO3。

表1 菌株的生理生化試驗(yàn)結(jié)果Table 1 Results of physiological and biochemical tests of strains
26S rDNA序列比對結(jié)果如圖2所示,XDNZ_PK01和XDNZ_PK02與其余9株菌親緣關(guān)系較遠(yuǎn),這暗示著這2株菌的生長或是發(fā)酵性能與其他菌株可能存在較大的差別。值得注意的是,菌株XDNZ_PK05與P.kudriavzeviiY1-9聚于一支,而P.kudriavzeviiY1-9與前期從貴州茅臺酒醅中篩選獲得的高耐乳酸酵母菌株P(guān).kudriavzeviiMT-Y01具有較高的親緣關(guān)系,這意味著菌株XDNZ_PK05可能具有與MT-Y01類似的發(fā)酵特性。
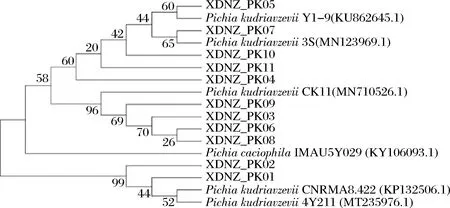
圖2 11株酵母菌株基于26S rDNA基因序列構(gòu)建的系統(tǒng)發(fā)育樹Fig.2 Phylogenetic tree of 11 yeast strains based on 26S rDNA gene sequences
綜合菌落形態(tài)、生理生化試驗(yàn)以及26S rDNA序列比對,確定這11個(gè)菌株均為庫德里阿茲威氏畢赤酵母,分別命名為XDNZ_PK01~XDNZ_PK11。
2.2 菌株的耐受性評價(jià)
溫度對微生物的生長代謝具有重要的影響。白酒釀造外界環(huán)境溫度和微生物發(fā)酵所產(chǎn)生的生物熱是影響發(fā)酵體系溫度最主要的2個(gè)因素,尤其是生物熱更是推動微生物群系演替的推動力[15],而酵母的生長代謝是生物熱的主要來源之一。如果釀造體系中酵母耐熱性能較差,隨著發(fā)酵體系溫度的升高,酵母生長代謝將受到顯著抑制,使得生物熱無法得到繼續(xù)積累,從而嚴(yán)重影響發(fā)酵進(jìn)程。以醬香型白酒堆積發(fā)酵階段為例,堆子頂溫>50 ℃才能達(dá)到工藝要求。由此可見,耐高溫是白酒功能酵母的重要屬性。菌株XDNZ_PK01~XDNZ_PK11在42 ℃培養(yǎng)條件下生長良好,其中XDNZ_PK04和XDNZ_PK05可以在45 ℃生長(圖3),這也與目前報(bào)道的庫德里阿茲威氏畢赤酵母可耐受的最高溫度一致[16]。

圖3 45 ℃高溫脅迫條件下菌株生長情況Fig.3 Growth of strains under high temperature stress at 45 ℃
酸和醇是白酒釀造體系中微生態(tài)調(diào)控風(fēng)向標(biāo),發(fā)酵初期酸度的提高有利于控制雜菌的過度繁殖,中后期的醇濃度提高有助于抑制產(chǎn)酸菌的生長,2個(gè)因素共同控制,從而確保產(chǎn)品的“量”和“質(zhì)”。因此,發(fā)酵體系中的功能酵母應(yīng)具有較好的耐酸、耐乙醇特性,從而確保發(fā)酵體系內(nèi)菌群的平衡。
在白酒釀造過程中,乙酸和乳酸是發(fā)酵體系中酸度的主要貢獻(xiàn)者[17],因此考察了篩選獲得菌株在不同乳酸、乙酸濃度下的生長狀況。結(jié)果如圖4-a所示,XDNZ_PK01~XDNZ_PK11均可在5 g/L乙酸條件下正常生長。而對于乳酸壓力脅迫而言,這些菌株表現(xiàn)出了不同的耐受能力,XDNZ_PK03~XDNZ_PK11都可以在70 g/L乳酸條件下生長,其中XDNZ_PK03、XDNZ_PK06、XDNZ_PK08以及XDNZ_PK11對乳酸的耐受能力顯著高于其他菌株(圖4-b)。
酵母是發(fā)酵體系中乙醇最主要的生產(chǎn)菌株,然而隨著體系中乙醇的不斷積累,高濃度乙醇會對酵母生長代謝產(chǎn)生抑制,從而影響基酒產(chǎn)量。因此,高乙醇耐受性是白酒功能酵母所必備的性能。將11株菌株分別置于含有14%(體積分?jǐn)?shù))乙醇的YPD平板上,30 ℃培養(yǎng)2 d。如圖4-c所示,除菌株XDNZ_PK08和XDNZ_PK11之外,其他菌株均可以正常生長。由此可知,這些菌株通過在釀造環(huán)境中長期的馴化,已進(jìn)化獲得了更好的乙醇耐受特性。

a-5 g/L乙酸;b-70 g/L乳酸;c-14%乙醇圖4 菌株在不同脅迫條件下生長情況Fig.4 Growth of strains under different stresses
2.3 菌株發(fā)酵性能和代謝特征評價(jià)
除了耐受性質(zhì)之外,酵母菌株的代謝特性也是影響基酒質(zhì)量的重要因素。利用玉米濃醪模擬白酒發(fā)酵,對這11株菌的發(fā)酵性能和風(fēng)味物質(zhì)的合成情況進(jìn)行了系統(tǒng)分析,并以實(shí)驗(yàn)室保藏的釀酒酵母AY12作為對照菌株。工業(yè)釀酒酵母AY12具有優(yōu)良的發(fā)酵及產(chǎn)酒性能[18]。如圖5所示,菌株XDNZ_PK01~XDNZ_PK11的發(fā)酵速度顯著低于釀酒酵母AY12,除了菌株XDNZ_PK09之外,其他菌株的乙醇產(chǎn)量也顯著低于AY12,且發(fā)酵失重與乙醇產(chǎn)量之間呈現(xiàn)很好的相關(guān)性。

a-失重變化;b-乙醇產(chǎn)量;c-失重與乙醇產(chǎn)量的關(guān)聯(lián)分析圖5 菌株在玉米濃醪發(fā)酵條件下代謝特性分析Fig.5 Analysis of metabolic characteristics of strains under corn concentrated mash fermentation注:圖b顯著性分析均是以AY12為對照進(jìn)行分析:ns代表無顯著性差異,*P<0.05,**P<0.01,***P<0.001,****P<0.000 1(下同)
同時(shí),利用氣相色譜法對這些菌株發(fā)酵所產(chǎn)生的主要風(fēng)味物質(zhì)進(jìn)行了檢測,結(jié)果如表2所示。
庫德里阿茲威氏畢赤酵母作為生香酵母可以顯著提高乙酸乙酯的含量,是釀酒酵母AY12乙酸乙酯產(chǎn)量的3.51~31.01倍,其中XDNZ_PK02的乙酸乙酯產(chǎn)量最高,達(dá)到了(796.92±1.84) mg/L。值得關(guān)注的是,菌株XDNZ_PK03和XDNZ_PK06的乳酸乙酯產(chǎn)量分別為(36.2±0.51)和(34.6±1.15) mg/L,顯著高于釀酒酵母AY12和其他庫德里阿茲威氏畢赤酵母的乳酸乙酯產(chǎn)量。此外,菌株XDNZ_PK01~XDNZ_PK11的高級醇產(chǎn)量顯著降低,僅為釀酒酵母AY12的45.43%~75.92%。由此推測,將庫德里阿茲威氏畢赤酵母作為強(qiáng)化菌劑應(yīng)用于傳統(tǒng)白酒發(fā)酵過程可以起到提酯降高級醇的效果。
根據(jù)菌株耐受性、發(fā)酵能力和風(fēng)味化合物合成能力分析,結(jié)果發(fā)現(xiàn)菌株XDNZ_PK03和XDNZ_PK06雖然具有較好的耐高溫和耐酸性能,但其乙醇產(chǎn)量過低,不能滿足實(shí)際生產(chǎn)的需要。而菌株XDNZ_PK09的乙醇產(chǎn)量達(dá)到(139.7±0.98) g/L,顯著高于其他菌株,且與釀酒酵母AY12產(chǎn)醇能力相當(dāng),但其延滯期較長,因此也不符合實(shí)際生產(chǎn)應(yīng)用的要求。進(jìn)一步分析顯示,與釀酒酵母AY12相比,菌株XDNZ_PK02、XDNZ_PK05合成的總酯分別提升了27.21倍和20.40倍,總高級醇分別降低了31.65%和36.37%。此外,XDNZ_PK05菌株可以在45 ℃條件下生長,耐熱性能優(yōu)于XDNZ_PK02。

表2 不同菌株在玉米濃醪發(fā)酵條件下風(fēng)味化合物的產(chǎn)量 單位:mg/L
2.4 強(qiáng)化菌劑原位發(fā)酵實(shí)驗(yàn)
根據(jù)以上菌株耐受性能分析和玉米濃醪模擬發(fā)酵的結(jié)果,選擇菌株XDNZ_PK05作為強(qiáng)化菌株進(jìn)行原位生產(chǎn)發(fā)酵驗(yàn)證。菌株XDNZ_PK05培養(yǎng)液按照106CFU/g的接種量應(yīng)用于第7輪次發(fā)酵生產(chǎn)過程中,堆積階段和窖池階段原位發(fā)酵的位置如圖6所示。
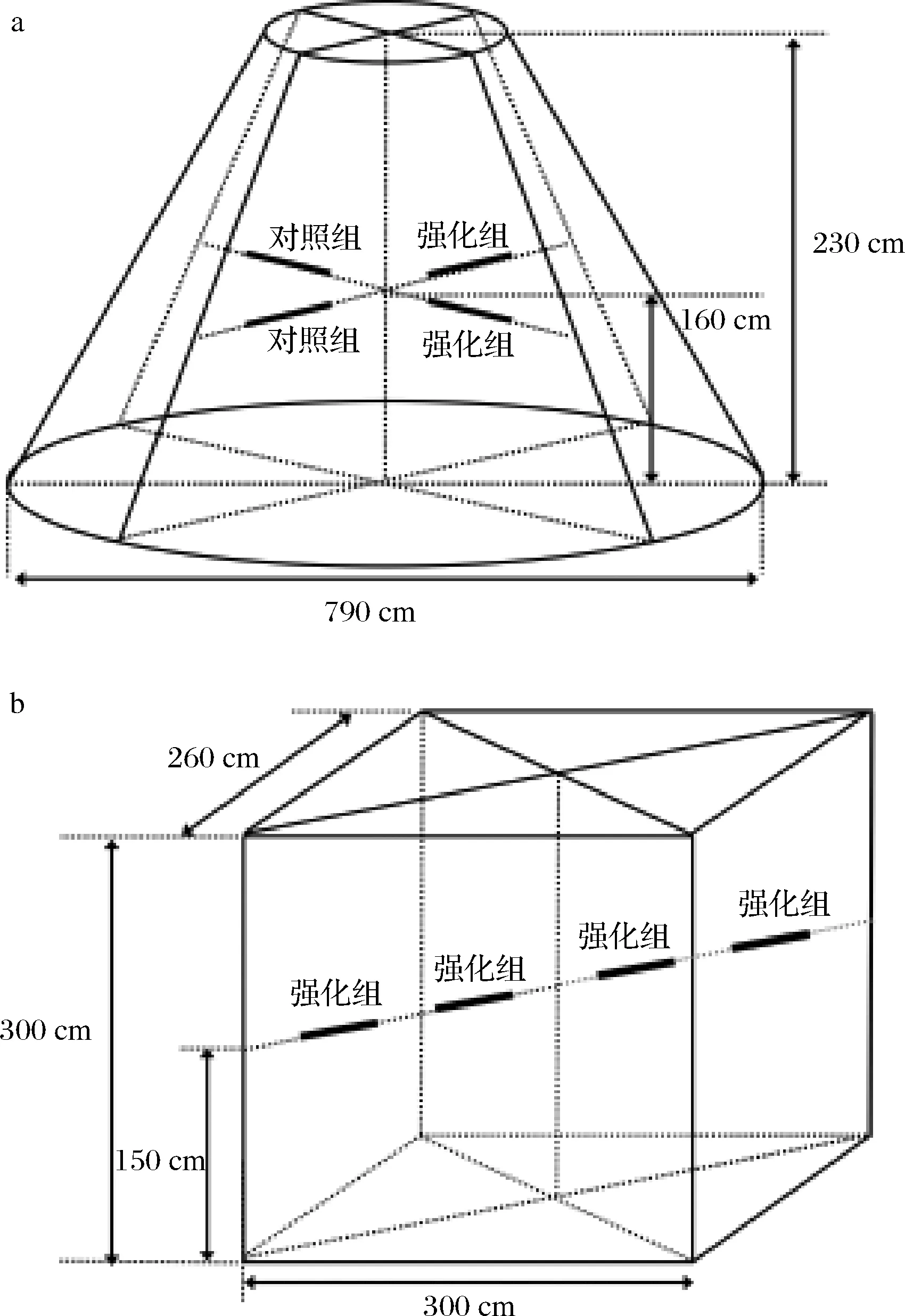
a-堆積階段;b-窖池階段圖6 原位堆積發(fā)酵和窖池發(fā)酵樣品位置示意圖Fig.6 Location diagram of samples during the in-situ stacking fermentation and pit fermentation
由表3可知,添加了XDNZ_PK05可以顯著提高原料的利用,堆積末殘淀粉從對照組的(14.4±0.18)%降低到了(12.10±0.04)%,利用率提高了15.97%,出窖時(shí)殘淀粉從(12.10±0.04)%降低到了(9.59±0.05)%,淀粉利用率提高了20.74%。盡管菌劑強(qiáng)化組的堆積發(fā)酵階段淀粉消耗增多,但酒醅中乙醇含量并未發(fā)生顯著變化,強(qiáng)化組窖池發(fā)酵結(jié)束后酒醅中的乙醇含量顯著提高了15.26%,從對照組的(6.42±0.17) g/L提升到了(7.40±0.10) g/L,這與基酒中乙醇產(chǎn)量提高的比率相似。對照組基酒中乙醇含量為(293.71±13.37) g/L,強(qiáng)化組基酒乙醇含量為(332.46±11.73) g/L,提高了13.19%。

表3 酒醅中淀粉和乙醇含量Table 3 The starch and ethanol contents in Jiupei
這些結(jié)果證實(shí)了耐酸菌劑的使用可以顯著提升高酸脅迫條件下原料利用率,解決發(fā)酵力不足、產(chǎn)能下降的實(shí)際生產(chǎn)問題。盡管目前鮮有關(guān)于利用庫德里阿茲威氏畢赤酵母作為強(qiáng)化菌劑進(jìn)行人工發(fā)酵調(diào)節(jié)的相關(guān)研究,但已有報(bào)道耐酸非釀酒酵母用于應(yīng)對異常天氣導(dǎo)致的糟醅酸度過高的問題,如貴州國臺酒業(yè)有限公司利用耐酸粟酒裂殖酵母作為強(qiáng)化菌劑,并開發(fā)與之適配的調(diào)控技術(shù),可以顯著提高基酒產(chǎn)量13.47%~25.97%[19]。由此可以推斷,高耐非釀酒酵母的篩選及相應(yīng)技術(shù)的開發(fā),是解決發(fā)酵力不足、高酸掉排這些實(shí)際生產(chǎn)問題的有效策略。
風(fēng)味物質(zhì)分析結(jié)果如表4所示,對照組基酒中正丙醇含量為(1 334.29±80.17) mg/L,而強(qiáng)化組中含量僅為(620.75±50.56) mg/L,顯著降低了53.48%,這意味庫德里阿茲威氏畢赤酵母的應(yīng)用可以有效降低基酒中高級醇的含量,有利于改善飲后舒適度,提升產(chǎn)品質(zhì)量。
與之相類似的,在小曲清香型白酒釀造過程中,釀酒酵母是高級醇的主要生產(chǎn)菌株,而畢赤酵母對降低高級醇有著積極貢獻(xiàn)[20]。在白云邊酒釀造過程中庫德里阿茲威氏畢赤酵母的應(yīng)用可以降低正丙醇含量27.4%[21]。然而,在葡萄酒釀造過程中,采用釀酒酵母和庫德里阿茲威氏畢赤酵母順序發(fā)酵工藝顯著提升高級醇產(chǎn)量20%左右[22]。由此可見,庫德里阿茲威氏畢赤酵母對高級醇的調(diào)控與發(fā)酵工藝息息相關(guān)。
此外,先前的玉米濃醪模擬發(fā)酵實(shí)驗(yàn)顯示庫德里阿茲威氏畢赤酵母可以顯著提高發(fā)酵體系中乙酸乙酯的產(chǎn)量(表2)。然而,強(qiáng)化組基酒中酯含量并沒有顯著變化。除此之外,其他風(fēng)味指標(biāo)無顯著性變化,不會對基酒品質(zhì)造成不良影響。

表4 酒樣中風(fēng)味物質(zhì)含量 單位:mg/L
3 結(jié)論
傳統(tǒng)白酒開放式的發(fā)酵方式在賦予酒體獨(dú)特風(fēng)味的同時(shí)也給生產(chǎn)的穩(wěn)定性和安全性帶來了潛在的隱患。釀造環(huán)境的異常波動會給釀造體系功能微生物帶來額外的脅迫壓力,從而導(dǎo)致菌群失調(diào),造成“減產(chǎn)降質(zhì)”的問題。本研究以開發(fā)耐酸脅迫人工可控發(fā)酵技術(shù)為目標(biāo),從醬香型白酒酒醅中篩選獲得了具有不同代謝特征和耐受能力的庫德里阿茲威氏畢赤酵母11株,優(yōu)選出XDNZ_PK05菌株作為強(qiáng)化菌劑,進(jìn)行了原位發(fā)酵實(shí)驗(yàn)。結(jié)果顯示高耐性酵母XDNZ_PK05的使用可以克服高酸壓力,提高糧食利用率,提升乙醇產(chǎn)量13.19%,有效降低高級醇含量50%以上,且不改變其他風(fēng)味指標(biāo)。本研究篩選獲得的高耐性酵母和形成的發(fā)酵調(diào)控技術(shù)為醬香型白酒可控發(fā)酵調(diào)控技術(shù)的開發(fā)提供了良好的素材,也為創(chuàng)建基于合成功能微生物組的現(xiàn)代釀造調(diào)控技術(shù)提供了重要的理論支撐,對我國傳統(tǒng)釀造行業(yè)的技術(shù)升級具有重要的應(yīng)用價(jià)值。
