基于生物信息學及分子動力學探討牛膝影響鐵死亡治療骨質(zhì)疏松機制
2023-02-17 03:13:40肖劍偉蔡旭郭粉蓮黃新民汪榮盛
特產(chǎn)研究 2023年1期
肖劍偉,蔡旭,郭粉蓮,黃新民,汪榮盛
(1.深圳市福田區(qū)風濕病專科醫(yī)院風濕免疫科,廣東 深圳 518000;2.上海市光華中西醫(yī)結合醫(yī)院風濕免疫科,上海 200052)
骨質(zhì)疏松癥(Osteoporosis,OP)被定義為一種全身性骨骼疾病,其特征是骨量、強度和微結構的系統(tǒng)性損害,從而增加了脆性骨折的發(fā)生率[1]。調(diào)查顯示,我國絕經(jīng)后的女性,OP的患病率為48%[2]。骨質(zhì)疏松癥的治療方式包括生活方式措施和藥物治療。生活方式包括改變飲食結構、補充礦物質(zhì)和鍛煉等。而藥物治療最常用的治療骨質(zhì)疏松癥的藥物包括雙膦酸鹽、阿侖膦酸鹽、唑來膦酸、地諾舒單抗和甲狀旁腺激素(PTH)衍生的藥物以及激素替代藥物等。然而,由于部分藥物存在較多不良反應限制了藥物的長期使用。
鐵死亡是最近發(fā)現(xiàn)的一種非凋亡的與脂質(zhì)過氧化相關的細胞死亡的形式[3],大量研究顯示其參與了神經(jīng)系統(tǒng)、癌癥等疾病的發(fā)生發(fā)展過程[4,5]。對鐵死亡通路相關靶點的調(diào)控有望成為治療疾病的一種新策略。
中藥中含有豐富的天然化合物,其在化學結構和生物活性方面也表現(xiàn)出豐富的多樣性,并且毒性更小。因此,使用中藥治療骨質(zhì)疏松成為當前研究熱點之一。在大量治療骨質(zhì)疏松的中藥中,牛膝味苦,性平,歸肝腎經(jīng),具有強筋骨、補肝腎等功效,十分適合治療以“腎虛”“脾虛”為病機的骨質(zhì)疏松。研究顯示,用牛膝提取物對去卵巢骨質(zhì)疏松大鼠治療后,治療組的骨密度(Bone mineral density,BMD)較對照組均有所提升,提示牛膝的活性成分能提升去勢大鼠的BMD[6]。而王艷等[7]的研究發(fā)現(xiàn),用牛膝治療去卵巢大鼠能夠抑制其骨量丟失,從而達到預防骨質(zhì)疏松的目的。以上結果顯示,牛膝具有的抗OP的療效,然而其具體機制尚需進一步明確。
本研究以生物信息學為基礎,篩選出與OP發(fā)病及鐵死亡相關的關鍵基因并進行試驗驗證,同時結合分子對接和分子動力學探討牛膝通過影響鐵死亡治療OP的作用機制。
1 材料與方法
1.1 數(shù)據(jù)檢索及數(shù)據(jù)差異基因分析
以“Osteoporosis”為關鍵詞,在GEO數(shù)據(jù)庫(www.ncbi.nlm.nih.gov/geo) 中檢索,檢索到GSE56116、GSE2208和GSE7158共3個數(shù)據(jù)集,見表1。使用R軟件(V 4.04)的limma包對編碼蛋白的mRNA進行差異分析,取adjust P<0.05及|LogFC|值大于0.5。
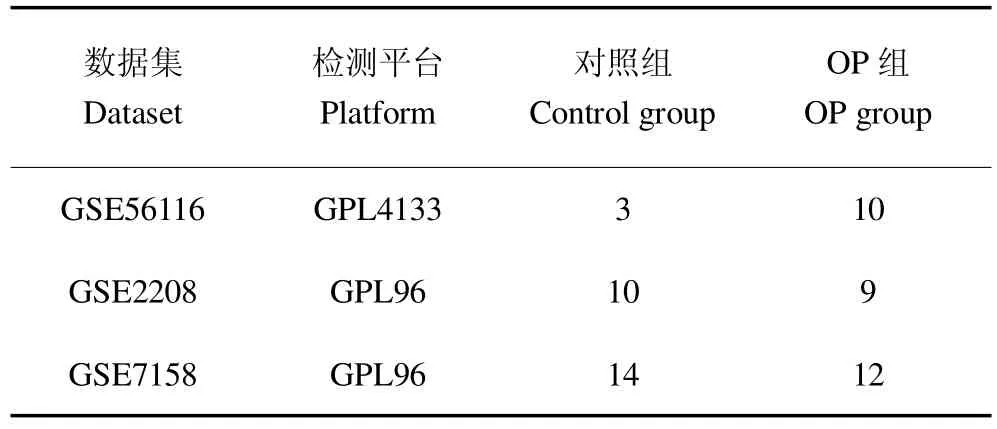
表1 GEO數(shù)據(jù)集Table 1 Details of the data set
1.2 預測牛膝作用靶點
通過中藥系統(tǒng)藥理學技術平臺(TCMSP)[8]檢索牛膝化學成分,將口服生物利用度(oral bioavailability,OB)≥40%及類藥性(drug likeness,DL)≥0.18作為活性小分子化合物的篩選條件,將具有較高活性的化合物進行篩選提取并將小分子化合物作用靶點轉化為基因名并使用Cytoscape 3.7.1軟件繪制“藥物化合物靶基因”相互作用網(wǎng)絡圖。
1.3 鐵死亡相關基因及靶點獲取
通過FerrDb數(shù)據(jù)庫(www.zhounan.org/ferrdb)[9],檢索鐵死亡相關基因。將上述結果與1.1、1.2步驟得到的靶點取交集,獲得牛膝影響OP鐵死亡途徑的潛在治療靶點并繪制韋恩圖。
1.4 試驗驗證
1.4.1 材料試驗采用小鼠單核細胞RAW264.7細胞株購自深圳市豪地華拓生物科技有限公司。ALOX12一抗(Abcam,ab168384)和二抗、GPX4一抗和二抗(Abcam,ab125066);內(nèi)參-actin(CST,#4970)。瓊脂糖凝膠(Solarbio,P1200)。BioTraceTMNT硝酸纖維素膜(Millipore,IPVH00010)。DMEM培養(yǎng)基、胎牛血清和胰蛋白酶(GIBCO,12430054、10100147、12605036);黃芩素(Sigma-Aldrich,491-67-8)、MTT試劑(Sigma-Aldrich,CT02)、二甲基亞砜(DMSO)(Sigma-Aldrich,C6164)、核因子-B受體活化因子配基(receptor activator of NF-B ligand,RANKL)(Sigma-Aldrich,207621-35-0)、巨噬細胞集落刺激因子(macrophagecolonystimulating factor,M-CSF)(Sigma-Aldrich,SRP3221)和青鏈霉素(sigma-Aldrich,P4333)。
1.4.2 蛋白質(zhì)印跡法(Western blot,WB)檢測潛在治療靶點根據(jù)上述步驟得到的差異基因ALOX12,通過WB驗證其在OP患者中的表達情況。選取福田區(qū)風濕病專科醫(yī)院風濕免疫科符合《中國老年骨質(zhì)疏松癥診療指南(2018)》OP診斷標準,排除合并有惡性腫瘤、近6個月內(nèi)服用過激素類藥物以及嚴重肝腎功能障礙的患者,共納入2020年4~6月確診的OP患者4例,其中男性2例、女性2例,年齡52~65歲,平均(58.8±5.4)歲。按照OP患者性別、年齡比例選擇4名深圳市福田區(qū)風濕病專科區(qū)院體檢健康者作為對照組,其中男性2名、女性2名,年齡50~63歲,平均(57.3±5.9)歲。2組之間年齡差異無統(tǒng)計學意義(t=3.757,P=0.624)。本研究通過醫(yī)院倫理委員會審查(倫理審查編號FS202001012),研究對象均自愿參與,簽署知情協(xié)議書。ALOX12一抗和二抗(Abcam,ab168384);內(nèi)參-actin(CST,#4970)。瓊脂糖凝膠(Solarbio,P1200)。BioTraceTMNT硝酸纖維素膜為美國(Millipore,IPVH00010)產(chǎn)品。
抽取靜脈血5 mL,樣本加入5 mL已經(jīng)混有苯甲基磺酰氟(PMSF)的蛋白裂解液(每1 mL裂解液加入10L PMSF)進行蛋白定量。取適量的蛋白樣品上樣,進行SDS變性10%聚丙烯酰胺凝膠電泳(SDS-PAGE),室溫搖動封閉1 h。將膜按一抗蛋白和-actin蛋白的印跡位置剪開,置于含對應一抗的Blotto溶液中,4℃搖動作用過夜。TBST溶液中搖動漂洗后加入二抗室溫作用,搖動漂洗后將膜置于Western LightningTMChemiluminescence Reagent顯色劑中1 min。常規(guī)曝光后進行顯影、定影處理。以-actin為內(nèi)參,使用Image J(V 1.8.0)軟件分析各條帶灰度值與內(nèi)參的灰度值比值作為目的蛋白的相對表達量。
1.4.3 WB檢測黃芩素對RAW264.7細胞GPX4、ALOX12表達的影響RAW264.7細胞在含有100U/mL青霉素和100g/mL鏈霉素、10%FBS、10%M-CSF、10%RANKL的DMEM中培養(yǎng),并置于溫度為37℃、CO2濃度為5%的培養(yǎng)箱中。黃芩素用微量的DMSO溶液溶解,然后用DMEM培養(yǎng)基將黃芩素溶液稀釋至試驗所需濃度,使DMSO的最終濃度不超0.1%。
取對數(shù)生長期的RAW264.7細胞,常規(guī)消化收集細胞,計數(shù)并調(diào)整細胞密度至1 104個/孔,接種到96孔板中,當細胞達到80%~90%融合度時,根據(jù)既往文獻黃芩素體外使用配制濃度報告[10],將試驗空白對照組及不同濃度的黃芩素組(5mol/L、10mol/L和20mol/L)處理48 h。
收集上述各組細胞用裂解液提取細胞蛋白,進行蛋白定量。取適量的蛋白樣品上樣,進行SDS變性10%聚丙烯酰胺凝膠電泳(SDS-PAGE),室溫搖動封閉1 h。將膜按一抗蛋白和-actin蛋白的印跡位置剪開,置于含對應一抗的Blotto溶液中,4℃搖動作用過夜。TBST溶液中搖動漂洗后加入二抗室溫作用,搖動漂洗后將膜置于WesternLightningTMChemiluminescence Reagent顯色劑中1 min。常規(guī)曝光后進行顯影、定影處理。以-actin為內(nèi)參,使用Image J軟件分析計算GPX4及ALOX12蛋白的相對表達。
1.5 分子對接
明確ALOX12在OP患者中的表達情況后,通過RCSB網(wǎng)站檢索得到靶點的晶體結構,Pubchem數(shù)據(jù)庫中獲取中藥活性小分子結構,使用AutoDock Vina對蛋白及小分子進行半柔性對接,選取結合能最小的結果作為最佳配體。
1.6 分子動力學
使用Gromacs進行分子動力學模擬[11]。通過ATB網(wǎng)站(atb.uq.edu.au/)[12]生成配體小分子位置限制文件。通過Gromacs內(nèi)置命令將小分子及靶蛋白轉換為拓撲結構文件。使用Gromos54a7_atb力場及SPC水分子模型,創(chuàng)建大小為1.2的十二面體水盒子并加入6個鈉離子來中和系統(tǒng)電荷。采用最速下降法使系統(tǒng)能量最小化。使用V-rescale進行步長為2 fs,總時間長為100 ps的NVT系綜平衡,然后使用Berendsen算法在1 atm壓力下進行100 ps的NPT系綜平衡。對蛋白質(zhì)中的重原子(非氫原子)施加位置限制后進行總時長為20 ns的動力學模擬,步長設定為2 fs。根據(jù)模擬得到的結果在去除體系的周期性邊界后提取20 ns時的結構作為最終復合物構象。使用Gromacs內(nèi)置命令計算復合物的均方根偏差(rootmeansquaredeviation,RMSD)、溶劑可及表面(solvent accessible surface area,SASA)、蛋白回旋半徑(Gyrate,GR)和氫鍵。使用g_mmpbsa軟件處理GROMACS生成的結構(.tpr)和軌跡(.xtc)文件,并計算小分子與蛋白的結合自由能[13]。使用VMD[14]、PyMOL[15]、LigPlot+軟件[16]對數(shù)據(jù)進行可視化處理。
1.7 統(tǒng)計分析
使用R軟件(V 4.04)進行統(tǒng)計學分析。臨床樣本的蛋白表達水平使用t檢驗進行比較。P<0.05具有統(tǒng)計學意義。
2 結果
2.1 差異分析結果
結果顯示與正常組對比,GSE56116有760個mRNA存在表達差異,其中上調(diào)295個,下調(diào)465個;GSE2208有114個mRNA存在表達差異,其中上調(diào)78個,下調(diào)36個;GSE7158有276個mRNA存在表達差異,其中上調(diào)152個,下調(diào)124個。
2.2 牛膝作用靶點
經(jīng)過篩選后得到牛膝有效成分9個。9個有效成分的作用靶點共106個,將其轉化為官方基因名(Gene Symbol)。使用Cytoscape 3.7.1將藥物、化合物和靶基因繪制“藥物化合物靶基因”相互作用網(wǎng)絡圖,見圖1。該互作網(wǎng)絡包含了116個節(jié)點和222條邊。
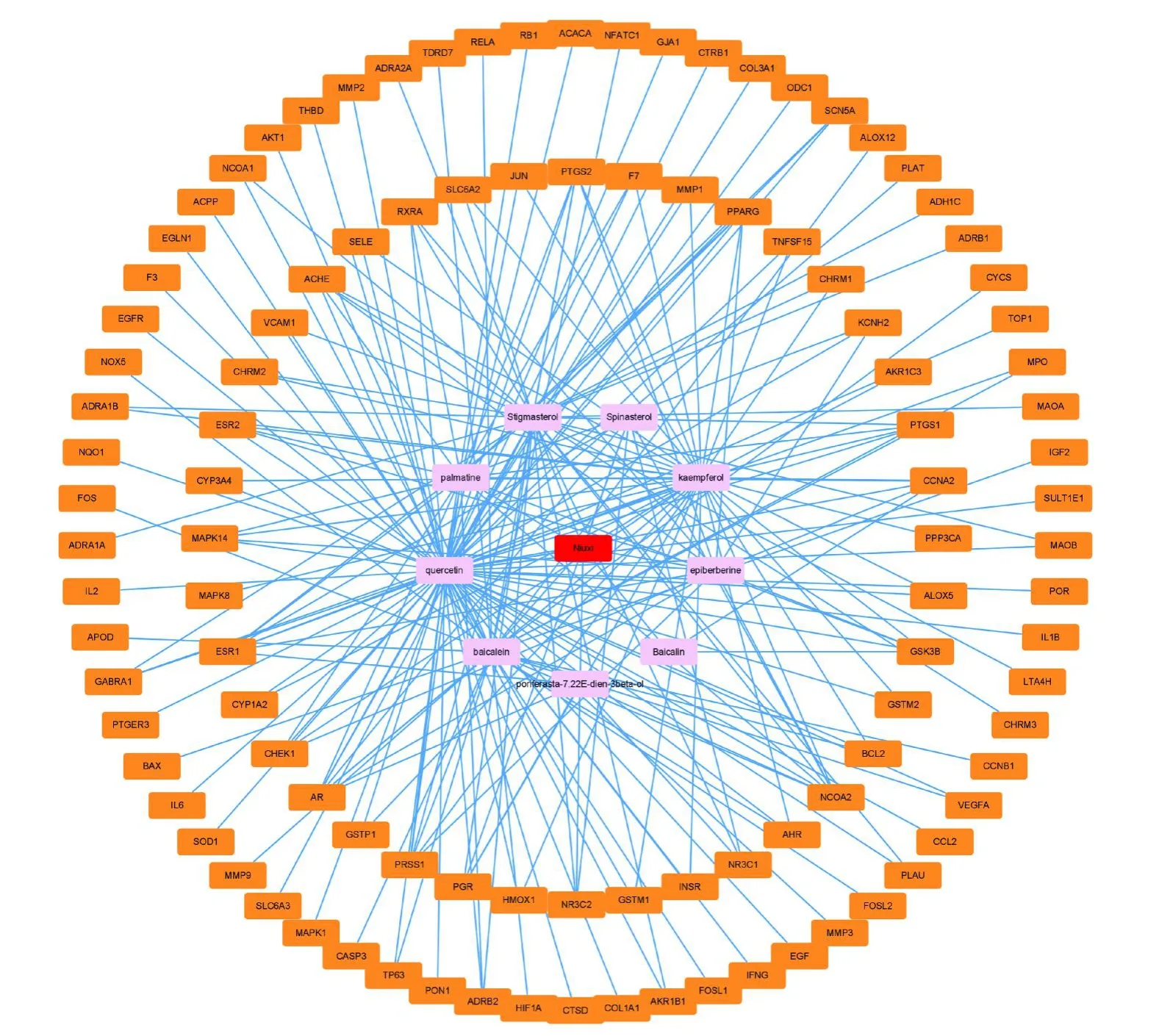
圖1 “藥物 化合物 靶基因”相互作用網(wǎng)絡圖Fig.1” Drug compound target gene” interaction network
2.3 牛膝治療OP靶點
根據(jù)1.3步驟獲得鐵死亡相關基因97個,與2.1、2.2步驟所得結果取交集(圖2),得到潛在通過影響鐵死亡治療OP的的靶基因1個(ALOX12)。
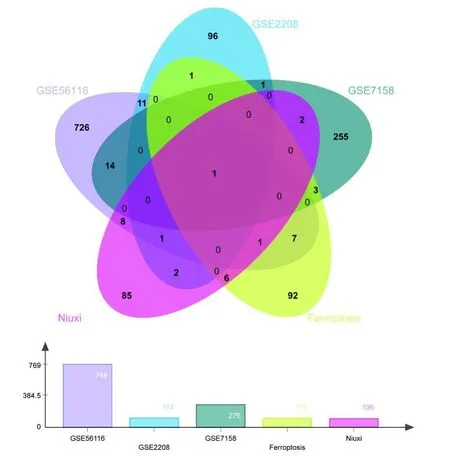
圖2 GSE56116、GSE22085、GSE7158、鐵死亡相關基因及牛膝作用靶基因交集的韋恩圖Fig.2 Venn diagram of intersection genes among the GSE56116,GSE22085,GSE7158,ferroptosis-associated genes and target genes of Achyranthes bidentata
2.4 試驗結果
2.4.1 蛋白質(zhì)印跡法結果以-actin為內(nèi)參,在OP患者全血中,與對照組相比,ALOX12蛋白表達明顯上升(正常組:0.63±0.17,OP組:0.99±0.34,t=19.38,P<0.05),見圖3。
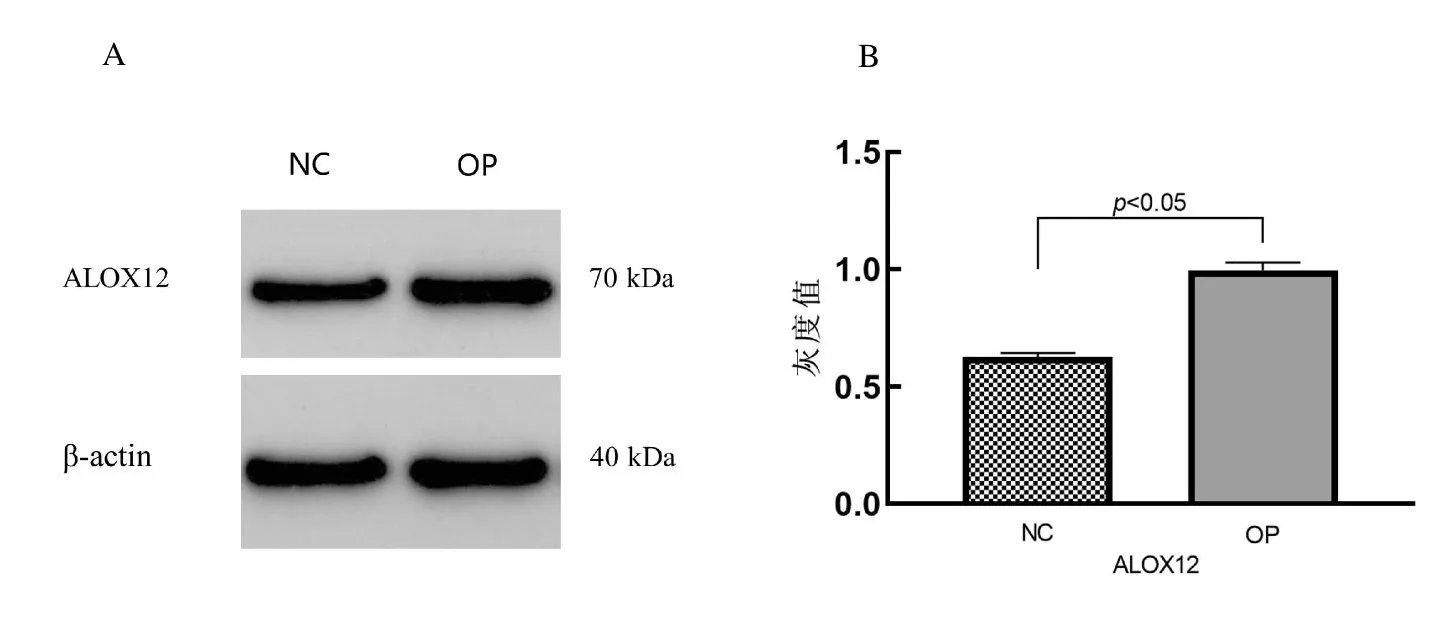
圖3 WB結果顯示OP組患者的ALOX12蛋白表達水平升高Fig.3 WB results showed the levels of ALOX12 protein expression increased in the OP group
2.4.2 黃芩素干預對RAW264.7細胞GPX4、ALOX12表達結果與對照組比較,不同濃度的黃芩素組RAW264.7細胞中ALOX12的蛋白表達水平明顯降低,差異有統(tǒng)計學意義,見圖4。而GPX4蛋白表達無明顯差異,表明黃芩素能抑制RAW264.7細胞中ALOX12的表達,而對GPX4蛋白表達無影響。
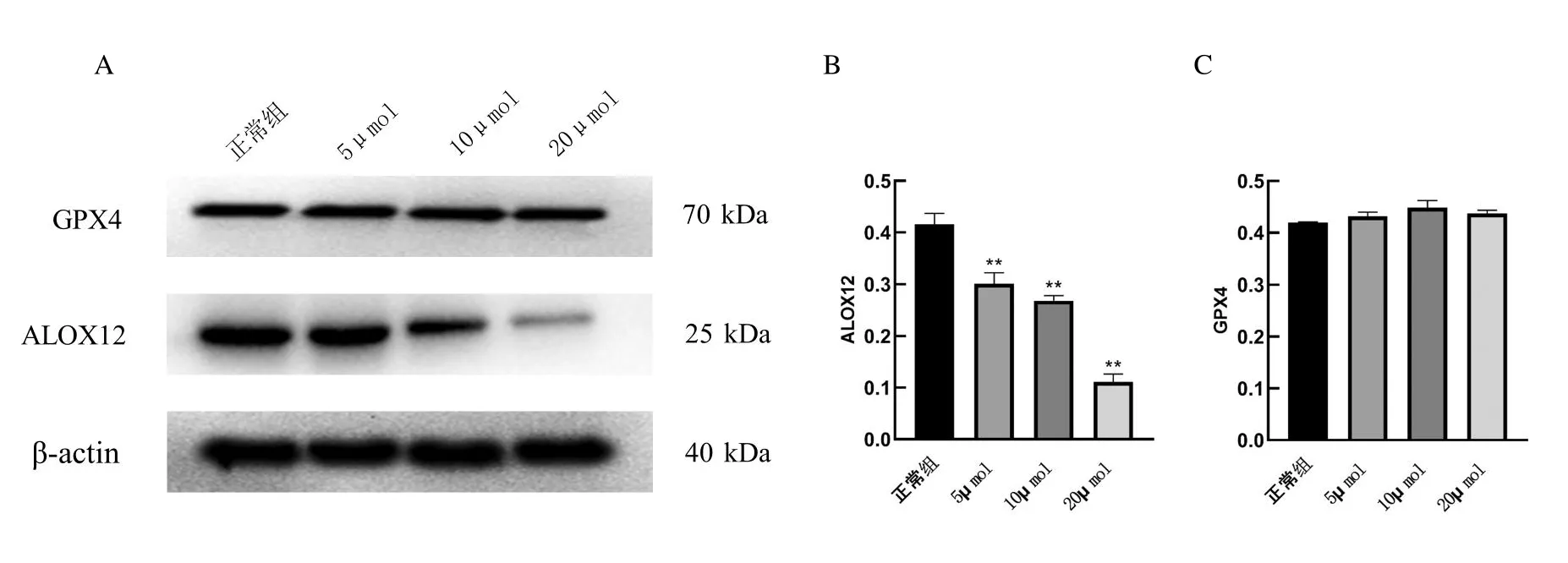
圖4 不同濃度黃芩素處理后對RAW264.7細胞GPX4、ALOX12表達的影響(**<0.001)Fig.4 Effects of different concentrations of baicalein on the expression of GPX4 and ALOX12 in RAW264.7 cells(**<0.001)
2.5 分子對接結果
根據(jù)2.2及2.3步驟篩選得到小分子化合物黃芩素(baicalein)作用的靶點及鐵死亡相關基因ALOX12。分子對接結果顯示,黃芩素與ALOX12的Arg189、His365形成氫鍵,與Leu366、Thr396、Glu369、Asn400、Thr662、Leu193和Val190形成疏水鍵,提示黃芩素與ALOX12有較強的結合能力,可能通過與其結合后抑制ALOX12活性,見圖5 A。
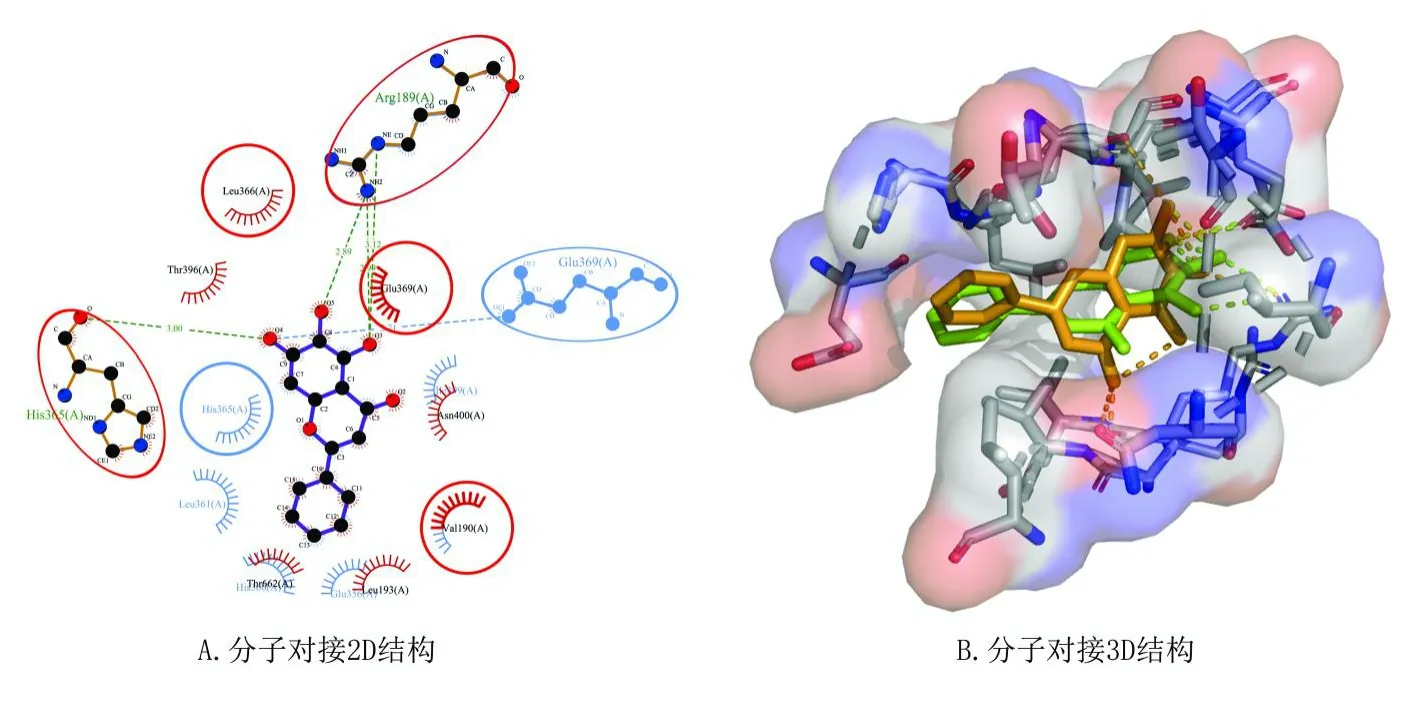
圖5 分子對接結果Fig.5 Molecular docking results
2.6 分子動力學模擬結果
黃芩素與ALOX12的復合體在整個動力學運行過程中逐漸穩(wěn)定,RMSD波動值小于0.1 nm(圖6 A)。復合物的溶劑可及表面呈逐漸縮小趨勢(圖6 B),復合物體系的蛋白回旋半徑(Rg)在模擬的過程中也逐漸縮小(圖6 C),提示蛋白與小分子結合后蛋白逐漸收斂并使得結構趨于穩(wěn)定。復合物體系氫鍵數(shù)量隨著模擬時間變化仍保持2個以上的連接(圖6 D),同時,與模擬前結構,活性小分子對比位置無明顯變化,提示兩者結合穩(wěn)定(圖5 B)。

圖6 蛋白小分子復合物體系的RMSD、SASA、RG和氫鍵數(shù)量Fig.6 RMSD,SASA,RG and number of hydrogen bonds of protein-small molecule complex systems
結合能計算結果顯示,黃芩素與ALOX12的總結合自由能為(107.893kJ/mol±16.671kJ/mol)。結果顯示,黃芩素與ALOX12主要通過范德華力(154.481kJ/mol±10.798 kJ/mol)和靜電勢能(68.847 kJ/mol±11.810 kJ/mol),而極性溶劑化作用則對結合起抑制作用(101.368 kJ/mol±19.941 kJ/mol)。
3 討論
在骨質(zhì)疏松癥發(fā)病的各種原因中,骨穩(wěn)態(tài)的破壞是其發(fā)病的關鍵即由成骨細胞介導的骨形成減少和由破骨細胞驅(qū)動的骨破壞增加,而鐵死亡在這個過程中發(fā)揮著重要的作用。鐵死亡的特征包括細胞內(nèi)鐵過載和活性氧類(Reactive oxygen species,ROS)的大量堆積。而既往研究發(fā)現(xiàn),鐵過載和氧化應激是骨質(zhì)疏松發(fā)病的重要因素之一,高度提示鐵死亡參與了骨質(zhì)疏松的發(fā)病過程。骨質(zhì)疏松癥患者存在明顯的鐵過載[17],其通過氧化應激損傷促進骨質(zhì)疏松癥的發(fā)生和發(fā)展[18]。研究顯示,鐵過載可影響成骨細胞的分化和活性[19],并且可以誘導成骨細胞的前體細胞骨髓間充質(zhì)干細胞的自噬和凋亡[20,21],而使用降低鐵過載的藥物可以改善骨質(zhì)疏松情況[22]。在氧化應激的狀態(tài)下,過載的ROS作為誘導鐵死亡模式啟動的重要因素。ROS在大多數(shù)情況可以被機體內(nèi)抗氧化酶清除。鐵超載可通過Fenton反應導致ROS形成增多[23]。ROS參與了成骨細胞的分化,過多ROS生成使正常的骨重建過程發(fā)生異常,使得骨形成減少而骨吸收增加,從而導致骨質(zhì)疏松[24,25],降低ROS水平能夠抑制破骨細胞分化[26]。另一個報道顯示[27],在去卵巢小鼠中,降低ROS水平可以抑制破骨細胞的產(chǎn)生從而阻止雌激素缺乏引起的骨質(zhì)疏松。試驗顯示在卵巢切除術后小鼠中,Loureirin B可通過抑制ROS活性來抑制破骨細胞的產(chǎn)生從而防止小鼠的骨丟失,進一步證實了ROS在骨質(zhì)疏松發(fā)病中的作用[28]。YANG等[29]發(fā)現(xiàn)內(nèi)皮細胞分泌的外泌體是細胞間通訊的重要介質(zhì),其通過抑制鐵蛋白吞噬作用依賴的鐵死亡,外泌體逆轉了糖皮質(zhì)激素誘導的成骨細胞對成骨的抑制作用。以上多個研究結果表明,鐵離子和氧化應激在骨質(zhì)疏松過程中具有重要的作用。而由鐵介導的氧化應激損傷導致的鐵死亡是近來新發(fā)現(xiàn)的細胞死亡形式,由鐵離子介導的鐵死亡可能導致骨質(zhì)疏松發(fā)生及病情進展。調(diào)控鐵死亡相關的靶點有望成為治療骨質(zhì)疏松潛在手段。
鐵死亡作為一種由鐵過載和脂質(zhì)過氧化驅(qū)動的細胞死亡方式,其實質(zhì)是細胞內(nèi)脂質(zhì)氧化物出現(xiàn)代謝障礙,在鐵離子催化作用下發(fā)生代謝異常,導致脂質(zhì)過氧化從而誘導細胞死亡。鐵死亡有明顯區(qū)別于細胞凋亡的形態(tài)學特征,如線粒體膜破裂增加。脂質(zhì)過氧化作為一種自由基驅(qū)動的反應,主要影響細胞膜中的不飽和脂肪酸。脂質(zhì)合成代謝失調(diào)會導致大量多不飽和脂肪酸積累,同時會消耗細胞內(nèi)的谷胱甘肽(GSH),導致更多的過氧化脂質(zhì)產(chǎn)生,從而提高了細胞對鐵死亡的敏感性。其中,多不飽和脂肪酸(PUFA)的氧化是由脂氧合酶(ALOXs)途徑介導的。LOXs是一種非血紅蛋白含鐵酶,主要功能是過氧化不飽和脂肪酸[30]。人類LOX家族共有6個成員(ALXOE3、ALOX5、ALOX12B、ALXO12、ALOX15B和ALOX15)。在鐵死亡途徑中,鐵可以增加LOXs的活性,隨后LOXs通過介導PUFA過氧化產(chǎn)生花生四烯酸等,促進鐵死亡。其中,ALOX12可以直接過氧化細胞膜上的磷脂形成磷脂過氧化物,然后在亞鐵離子的催化下,脂質(zhì)過氧化物與其他脂質(zhì)發(fā)生鏈式氧化反應,導致鐵死亡的發(fā)生[31]。多個研究顯示,ALOX12的多態(tài)性與骨質(zhì)疏松發(fā)病呈正相關[32,33],提示ALOX12與骨質(zhì)疏松存在密切關系。一項研究顯示,黃芩素能夠與鐵結合,抑制血小板中ALOX12的表達[34]。Dai等[35]發(fā)現(xiàn),黃芩素能夠抑制ALOX12,減少鐵死亡,從而改善CCl4誘導的小鼠急性肝損傷。Bo Chu等[36]通過CRISPR/Cas9技術來生成敲除ALOX12的U2OS細胞的亞克隆,敲除了ALOX12基因的U2OS細胞鐵死亡受到抑制。由此可推斷,ALOX12是黃芩素的作用靶點之一,通過靶向ALOX12抑制其活性,可以影響鐵死亡過程,見圖7。

圖7 牛膝治療OP的機制圖Fig.7 Mechanism diagram of Achyranthes bidentata for the treatment of OP
祖國傳統(tǒng)醫(yī)學根據(jù)OP的表現(xiàn)將其歸屬于“骨痹”、“骨痿”和“骨枯”等病范疇。腎藏精,精血養(yǎng)骨髓,髓枯則筋痿,腎虛能夠誘發(fā)骨質(zhì)疏松。從西醫(yī)學角度來看,骨髓干細胞能夠分化為成骨細胞及破骨細胞。鐵死亡會導致炎癥水平升高,產(chǎn)生的各種炎癥因子如腫瘤壞死因子-等,能夠抑制骨髓間充質(zhì)干細胞成骨分化的生物效應,使骨形成減少導致骨量丟失,這與中醫(yī)腎虛導致的“骨枯”具有異曲同工之處。姚新苗認為OP多屬本虛標實,本虛責于脾腎而標實責于瘀血,在運用補腎壯骨、健脾益氣治療的同時應當輔以活血化瘀之藥,常選用赤芍、川芎、丹參和牛膝等[37]。陳雯等歸納中藥治療骨質(zhì)疏松用藥頻次,活血化瘀藥作為重要治法,在活血兼有養(yǎng)血功能的中藥處方中,當歸使用頻次為44次,骨碎補使用頻次為28次,牛膝使用頻次為26次,排在活血化瘀藥物的前三位[38],提示牛膝在治療OP中有著廣泛的應用。現(xiàn)代研究證實,牛膝及含有牛膝的復方藥物治療骨質(zhì)疏松療效明確,如含有牛膝的左歸丸、無敵丹膠囊具有滋陰補腎、填精益髓的作用,在臨床已被證實能夠改善OP患者癥狀[39,40]。牛膝含有大量的活性小分子成分,其中作為有效成分的黃芩素具有抗炎抗氧化作用[41]。研究顯示黃芩素可以抑制RANKL誘導的破骨細胞的形成以及抑制破骨細胞的骨吸收功能[10],提示黃芩素是治療骨質(zhì)疏松的潛在藥物分子。
本研究顯示,在OP患者中,ALOX12蛋白表達較健康組明顯升高,提示其可能參與了OP的發(fā)病。作為牛膝有效成分之一的黃芩素,通過分子對接及分子動力學結果顯示,其能通過多個氫鍵及疏水鍵與ALOX12緊密對接并形成穩(wěn)定結構。在動力學模擬過程中,整個復合物體系能夠快速收斂并達到穩(wěn)態(tài),說明黃芩素能夠通過影響ALOX12蛋白的結構穩(wěn)定性從而發(fā)揮抑制其活性的作用。試驗結果顯示,不同濃度的黃芩素可以抑制RANKL誘導的RAW264.7細胞ALOX12蛋白的表達,而不影響GPX4蛋白的表達。以上結果提示ALOX12可能作為牛膝治療OP的潛在靶點,抑制ALOX12的活性,達到治療骨質(zhì)疏松的目的。
綜上所述,本研究以生物信息學、網(wǎng)絡藥理學、分子對接及分子動力學結合試驗研究驗證牛膝有效活性成分黃芩素可能通過靶向ALOX12影響OP鐵死亡途徑從而達到治療OP的作用,初步闡明了牛膝有效活性成分治療OP的潛在作用機制,為進一步探討牛膝通過鐵死亡途徑治療OP提供了理論依據(jù)。然而,本研究的結果未通過動物模型研究證實,需要進一步通過使用體內(nèi)外試驗來驗證。
