不同循環饑餓投喂模式對許氏平鲉生長和體成分的影響
2023-02-02 13:14:02劉凱凱宋靜靜遲雯丹于道德
水產科學 2023年1期
關鍵詞:生長
劉凱凱,丁 剛,李 樂,宋靜靜,遲雯丹,于道德
( 山東省海洋科學研究院,山東 青島 266104 )
在自然環境中由于溫度、遷徙或繁殖等因素,魚類在生活周期的一定階段會面臨食物緊缺問題[1-2],人工養殖過程中投餌不足或者不均也會導致魚類遭受饑餓脅迫[3],不同種類的魚類對饑餓的耐受程度不同。遭受饑餓脅迫的魚類在恢復攝食后,會出現一段快速生長期,其生長速度會快于同期未受饑餓脅迫的魚類,這一現象稱為魚類的補償生長[4]。魚類補償生長的程度受各種因素的影響,如魚的種類、生長階段、饑餓的時間和程度等[5-6]。在水產養殖實際應用中,進行適當限飼,激發的魚類補償生長效應,能夠促進魚類生長和飼料利用、降低勞動成本、減少水體污染[7]。然而,魚類的饑餓耐受力是有限度的,超過一定限度,魚類會因攝食不足而發生生理機能衰退,喪失補償生長能力,特別是在幼魚階段[8]。因此,在實際應用到水產養殖之前,有必要確定特定魚類補償生長的適當喂養方案。
在魚類補償生長的研究中,常用的投喂模式有2種,分別是“饑餓-投喂”的單期饑餓模式和“饑餓-投喂-再饑餓-再投喂”的循環饑餓模式[9]。相比于單期饑餓,循環饑餓能夠使魚類補償生長的能力在周期攝食期間逐漸減弱時重新激活,可以不斷增強補償生長的潛力,從而增加代償性生長的持續時間,促進魚類的生長[10]。目前利用循環饑餓進行補償生長研究的魚類較多,如太平洋鮭(Oncorhynchussp.)[10]、大西洋庸鰈(Hippoglossushippoglossus)[11]、異育銀鯽(Carassiusauratusgibelio)[12]和錦鯉(Cyprinuscarpiokoi)[13]等。
許氏平鲉(Sebastesschlegelii),肉質鮮美,可在我國北方海域自然越冬,已成為黃、渤海區域大型海水網箱養殖的主要魚類品種之一,是一種重要的海產經濟魚類和增養殖優良品種[14]。目前關于許氏平鲉生長補償研究主要集中在單期饑餓模式方面[15-16],循環饑餓情況下的相關研究尚未見報道。筆者探討不同循環饑餓投喂模式對許氏平鲉生長和體成分的影響,旨在為建立一種減輕勞動力強度、降低養殖成本的科學投喂策略提供理論參考。
1 材料與方法
1.1 試驗材料
試驗于2020年9月10日—11月16日在中國水產科學研究院長島增殖實驗站進行。試驗用許氏平鲉為該增殖實驗站2020年4月中旬繁育的同批次5月齡幼魚。試驗開始前7 d挑選規格相近的幼魚進行暫養,每日投喂2次海童牌浩食系列配合飼料,每次投喂至表觀飽食。養殖用水為附近過濾海水,水溫(23±1) ℃,pH 7.50~7.95,鹽度28~33,溶解氧7.50~7.74 mg/L,氨氮質量濃度0.04~0.08 mg/L,日換水1次,換水率100%。
1.2 試驗方法
試驗開始前24 h停止投喂,隨機挑選120尾魚測量初始體長、體質量、內臟質量、肝臟質量,-20 ℃保存,取4尾魚測定魚體體成分,作為初始對照值。試驗設置6個組,每組2個平行,每個平行420尾魚。為了貼近生產實際,每組試驗在4.0 m×4.0 m×1.2 m的養殖水泥池中進行,水深1.1 m。投喂模式分別為:正常每日投喂組(對照組,簡稱S0, 下同)、饑餓2 d投喂1 d組(S2F1)、饑餓2 d投喂2 d組(S2F2)、饑餓2 d投喂3 d組(S2F3)、饑餓2 d投喂4 d組(S2F4)、饑餓2 d投喂5 d組(S2F5)。投喂時間分別為7:00和14:00,投喂至表觀飽食后,用虹吸法收集殘餌烘干稱量質量。試驗結束前24 h停止投喂,每組隨機取30尾魚測量體長、體質量等指標,-20 ℃冷凍保存,每組取4尾魚分別測定魚體成分。
1.3 數據測定與處理
采用凱氏定氮法測定魚體樣品的粗蛋白質含量,索氏提取法測定粗脂肪含量,分別用直接干燥法和灼燒法測定水分和灰分含量。計算公式如下:
wWGR=(mt-m0)/m0×100%
(1)
RDFI=mc/[t×(m0+mt)/2]×100%
(2)
RSG=(lnmt-lnm0)/t0×100%
(3)
wVSI=mv/mb×100%
(4)
wHSI=mh/mb×100%
(5)
CF=100mb/L3
(6)
RFC=(mt-m0)/mc×100%
(7)
式中,wWGR為質量增加率(%),RDFI為攝食率(%),RSG為特定生長率(%/d),wVSI為臟體指數(%),wHSI為肝體指數(%),CF為肥滿度,RFC為飼料效率(%),mt和m0為試驗結束和開始時魚體質量(g),mv為內臟質量(g),mh為肝臟質量(g),mb為魚體質量(g),mc為攝食飼料總量(g),t0為試驗時間(d),t為投喂時間(d),L為體長(cm)。
試驗結果用平均值±標準差表示,利用SPSS 20.0軟件采用單因素方差和鄧肯多重比較進行分析,以P<0.05為差異顯著。
2 結 果
2.1 生長性能
試驗過程中,各組許氏平鲉的成活率為100%。不同循環饑餓投喂模式對許氏平鲉生長性能的影響情況見表1。試驗結束時,S2F1組的終末均體長最小,S2F5組魚體終末均體長最大,各試驗組間差異不顯著(P>0.05)。魚體終末均體質量最高的是S2F5組,為(61.15±0.58) g,與S0、S2F1、S2F2、S2F3和S2F4組差異顯著(P<0.05)。S2F5組的質量增加率最高,為264.62%,其次為S0和S2F4組,顯著高于其他試驗組(P<0.05)。S2F5組的攝食率最高,為2.49%,比S0組顯著高約20%,與其他循環投喂試驗組差異顯著(P<0.05)。S2F5、S0和S2F4組的特定增長率較高,3組間差異不顯著(P>0.05)。各循環饑餓投喂試驗組的肝體指數、飼料效率均與S0組差異不顯著(P>0.05)。各循環饑餓投喂試驗組的臟體指數顯著低于S0組(P<0.05)。肥滿度最高的組是S2F4和S2F5組,與S0組差異不顯著(P>0.05)。
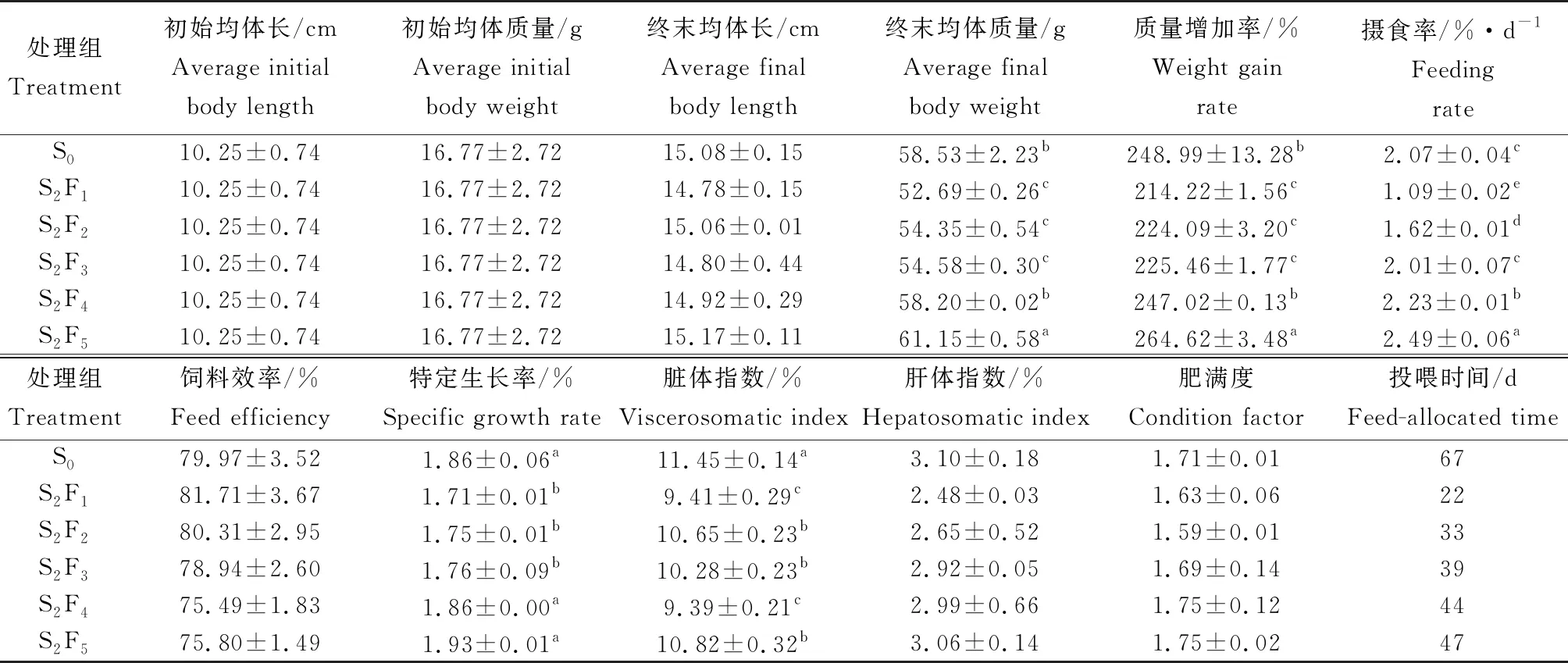
表1 不同循環饑餓投喂模式對許氏平鲉生長性能的影響
2.2 魚體成分
不同循環饑餓投喂模式對許氏平鲉體成分的影響結果見表2。經過循環饑餓投喂處理后,循環饑餓投喂組的魚體粗蛋白質含量較S0組降低,但各組間差異不顯著(P>0.05)。除S2F5組外,各循環饑餓投喂組的魚體粗脂肪含量顯著低于S0組(P<0.05)。S2F1組的魚體水分含量最高,與S2F2、S2F3、S2F4和S2F5組差異不顯著(P>0.05),而與S0組差異顯著(P<0.05)。隨著饑餓時間的增加,魚體灰分含量逐漸增加,但各組間差異不顯著(P>0.05)。
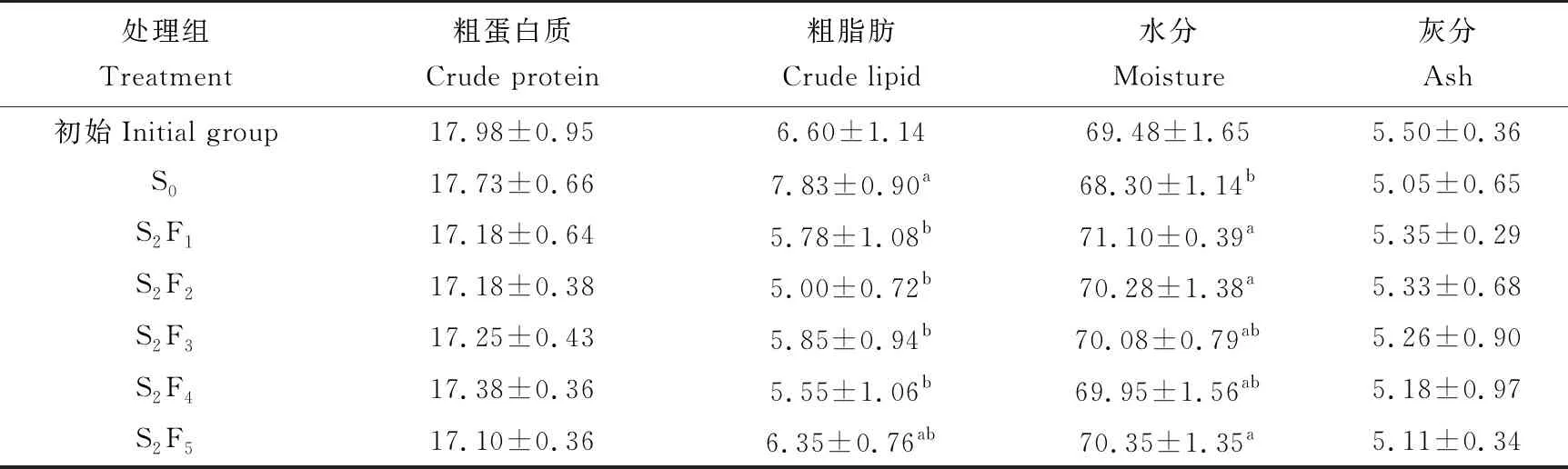
表2 不同循環饑餓投喂模式對許氏平鲉體成分的影響 %
3 討 論
3.1 不同循環饑餓投喂模式對許氏平鲉生長的影響
補償生長是動物遭受饑餓脅迫時采取的生存策略,首先見于畜禽類動物[17],后來在魚類中也廣泛存在。魚類的補償生長可分為4類:超補償生長、完全補償生長、部分(有限)補償生長和不能補償生長[18]。已有大量研究表明,在單期或者循環饑餓投喂模式下,許多魚類均表現出不同程度的補償生長效應[19]。冒樹泉等[16]采用單期饑餓投喂模式喂養許氏平鲉,饑餓3、6、9 d和 12 d后分別投喂30、27、24、21 d和18 d,發現前2組分別出現超補償和完全補償生長,后2組表現為不能補償生長。鐘娟等[20]采用饑餓1 d投喂6 d、饑餓2 d投喂5 d、饑餓3 d投喂4 d的循環饑餓投喂模式飼養草魚(Ctenopharyngodonidella),發現前2種投喂模式組獲得補償生長,而最后1組未獲得補償生長。本試驗結果顯示,5種循環饑餓投喂模式下,隨著總饑餓時間的增加,魚類消耗自身的儲能物質,體質量逐漸下降。根據動物補償生長的定義:本試驗中饑餓2 d投喂5 d組魚類的質量增加率明顯高于對照組,表現為超補償生長;饑餓2 d投喂4 d組魚類的質量增加率和特定生長率與對照組差異不顯著,表現為完全補償生長;饑餓2 d投喂1 d組、饑餓2 d投喂2 d組和饑餓2 d投喂3 d組魚類的質量增加率和特定生長率顯著低于對照組,表現為不能補償生長。
3.2 許氏平鲉循環饑餓投喂模式下補償生長的生理機制
關于補償生長機理的理論主要存在以下3種觀點:(1)饑餓使魚類代謝降低,魚類通過提高飼料效率獲得補償生長[21];(2)恢復投喂后,魚類食欲和攝食量增加,表現為攝食率提高[19];(3)補償生長是由飼料效率提高和攝食率增加共同作用的結果[22]。本試驗中,各組間的飼料效率無明顯差異,而饑餓2 d投喂5 d組和饑餓2 d投喂4 d組的攝食率明顯高于對照組。由此推測,饑餓2 d投喂5 d組和饑餓2 d投喂4 d組的許氏平鲉是通過提高攝食率來獲得補償生長。
3.3 不同循環饑餓投喂模式對許氏平鲉形體指標的影響
在魚體饑餓和恢復投喂情況下,肥滿度、肝體指數和臟體指數的變化一定程度上可以反映其自身營養物質的消耗和積累[16]。不同魚類在饑餓和恢復投喂情況下,肥滿度的變化情況不同。金頭鯛(Sparusaurata)[23]在饑餓和恢復投喂后只有體質量出現了補償性生長,體長無明顯變化,導致肥滿度增加。黃河鯉(C.carpiohaematopterus)[24]經過15 d饑餓后,與對照組相比肥滿度降低,恢復投喂后無明顯改善。本試驗中,許氏平鲉各組的肥滿度雖無統計學意義上的顯著性,但饑餓2 d投喂4 d組和饑餓2 d投喂5 d組的肥滿度與對照組相比有所增加。循環饑餓投喂組魚體的肝體指數和臟體指數均比對照組小,推測饑餓條件下,魚體消耗肝臟和內臟貯存的營養物質,導致內臟和肝臟質量降低,肝體指數和臟體指數降低。
3.4 不同循環饑餓投喂模式對許氏平鲉魚體成分的影響
魚體受到饑餓脅迫后會消耗機體的營養物質,粗蛋白質、粗脂肪、水分等體成分含量會發生變化。多數魚類饑餓狀態下優先消耗糖原和脂肪,其次消耗蛋白質[25]。本試驗結果顯示,與對照組相比,除饑餓2 d投喂5 d組外,各循環饑餓投喂組魚體的粗脂肪含量顯著降低。魚體粗蛋白質含量,各組間無明顯差異,說明魚體在饑餓脅迫下首先消耗機體內貯存的脂肪,隨著饑餓時間的延長,才開始消耗蛋白質。與對照組相比,循環饑餓投喂組魚體灰分和水分含量增加,這是由于魚體消耗了機體內的貯能物質,導致魚體灰分和水分等的相對含量增加,這與對中華鱘(Acipensersinensis)幼魚[26]和錦鯉[13]的研究結果一致。
4 結 論
經過“饑餓—投喂—再饑餓—再投喂”的循環饑餓模式后,饑餓2 d投喂5 d組的許氏平鲉幼魚獲得超補償生長,饑餓2 d投喂4 d組的幼魚獲得完全補償生長,而饑餓2 d投喂1 d組、饑餓2 d投喂2 d組和饑餓2 d投喂3 d組幼魚未獲得補償生長。饑餓2 d投喂5 d組和饑餓2 d投喂4 d組的許氏平鲉幼魚攝食率明顯高于對照組,推測其補償生長效應是通過提高攝食率來實現的。在本試驗條件下,饑餓2 d投喂5 d的循環饑餓投喂模式的投喂時間比對照組縮短29.85%,縮減了勞動力成本,從補償生長效果和實際投喂時間考量,是最佳的投喂模式。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
