木薯MeMYC2.2基因耐低溫功能研究
2023-02-02 06:46:52于曉玲李文彬李智博阮孟斌
生物技術通報 2023年1期
于曉玲 李文彬 李智博 阮孟斌
(1 中國熱帶農業科學院熱帶生物技術研究所 農業農村部熱帶作物生物學與遺傳資源利用重點實驗室,海口 571101;2 海南熱帶農業資源研究院 海南省熱帶農業生物資源保護與利用重點實驗室,海口 571101)
木薯(Manihot esculentaCrantz)起源于南美洲,是世界上第六大糧食作物[1],其最適生長溫度為25-29℃[2],低于10℃發育停滯[3]。低溫條件下木薯發育遲緩、毒性物質累積,品質及產量都受到很大的影響[4]。因此木薯耐低溫研究,對木薯產業發展具有重要的作用。
國內外目前對木薯低溫響應的分子機理尚不清晰,研究主要集中在活性氧(reactive oxygen species,ROS)系統方面。增加ROS 的清除能力可以提高木薯對低溫的耐受性[5-6];Cheng 等[7]的研究表明,植物特異轉錄因子MeTCP4(teosinte branched/ cycloidea / PCF,TCP)的過量表達可以誘導ROS途徑相關基因的表達,增強擬南芥對冷脅迫耐受性; 過表達MeCu/ZnSOD和MeAPX2(ascorbate peroxidase,APX)提高活性氧清除能力,降低H2O2積累,提高轉基因木薯對低溫的耐受性[6]。另外,CBF3(c-repeat-binding factor,CBF)在木薯中過表達,可增強木薯對低溫的耐受性[8];干擾木薯MeMYB2的表達也可以提高木薯轉基因植株對低溫的耐受能力[9]。此外,也可以通過施肥提高木薯的抗寒能力[10]。
MYC2 轉錄因子是bHLH(the basic helix-loophelix)轉錄因子家族成員之一,其bHLH 保守結構域可與DNA 結合發揮作用[11]。近年來,在擬南芥、番茄、煙草、長春花、丹參等多種植物中對MYC2 基因進行了研究[12],結果顯示,MYC2 在植物應答逆境刺激時起重要調控作用,是茉莉酸(jasmonic acid,JA)信號傳導途徑的核心調控元件,同時也是JA 通路與其他植物激素信號通路交互的關鍵節點[13]。赤霉素(gibberellin acid,GA)途徑DELLA 阻遏物和JAZ 與MYC2 相互作用以抑制花中倍半萜的生物合成[14];而脫落酸(abscisic acid,ABA)促進ABA 受體PYL6(pyrabactin resistance 1-like 6)與MYC2 的相互作用,并阻遏MYC2 與靶啟動子的結合[15];擬南芥MYC2 可能在EDR1(enhanced disease resistance 1)基因上游調控JA-SA(Sal Icylic acid)的信號互作[16];蘋果MdMYC2 與ERF3(ethylene response factor 3)啟動子結合,調控乙烯生物合成[17]。低溫及外源MeJA 可誘導香蕉MYC2 的表達,且MYC2可與ICE1(inducer of CBF expression 1)互作調節可香蕉的抗凍性[18]。
生產中木薯表現為對干旱具有良好的耐受性,但對低溫敏感,但其分子機理尚不清楚。MYC2 在其他作物中表現出良好的抗寒作用,其在木薯中的作用尚未清楚。本研究通過前期低溫轉錄組測序結果,發現MeMYC2 基因受低溫脅迫誘導表達[19]。為進一步明確MeMYC2 基因的功能,本研究借助過表達木薯MeMYC2 基因的擬南芥,驗證MeMYC2 影響低溫耐受性的功能,旨在為進一步利用該基因提高栽培木薯對低溫的耐受性提供理論參考。
1 材料與方法
1.1 材料
擬南芥(Col-0)由中國熱帶農業科學院熱帶生物技術研究所保存。
1.2 方法
1.2.1 木薯MeMYC2.2 基因的克隆 利用TaKaRa 高保真酶(R045),以木薯cDNA 為模板,以特異性引物(表1)PCR 克隆木薯MeMYC2.2 基因[19]。PCR反應程序:98℃變性10 s,55℃退火 5 s,72℃延伸10 s,共計35 個循環。PCR 產物通過1.2%瓊脂糖凝膠電泳回收純化、與T 載體連接后轉化大腸桿菌DH5α;經菌落PCR 及酶切鑒定為陽性克隆的,進行測序(上海生工)。所用引物及其序列如表1所示。

表1 本研究所用引物Table 1 Primers used in this study
利 用NCBI Conservered Domain(https://www.ncbi.nlm.nih.gov/cdd)預測基因序列的功能結構域;運用在線工具PI_tool(https://www.expasy.ch/tools/pi_tool.html)預測基因編碼蛋白的分子量和等電點。
1.2.2 木薯MeMYC2.2 基因轉錄活性分析及亞細胞定位 將MeMYC2.2 基因開放閱讀框插入pGBKT7載體多克隆位點,并轉化酵母Y187 感受態細胞。將鑒定為陽性的轉化細胞與轉化空白對照載體pGBKT7 的酵母菌液活化,濃度調整一致后,劃線于SD/-Trp 平板(+40 μg/mL X-α-Gal),30℃倒置培養3-5 d 后,觀察酵母顯色情況。
利用XbaI 和BamH I 限制性內切酶,將MeMYC2.2 基因全長編碼序列(不包括TGA 終止子)克隆到帶有35S:GFP 植物表達載體中,形成35S:MeMYC2.2:GFP 融合表達載體。將上述重組質粒采用化學法分別轉化到農桿菌LBA4404 中。將陽性克隆菌通過葉盤注射法瞬時轉化煙草葉片。利用激光共聚焦顯微鏡(Olympus FV1000),在GFP 的激發波長(488 nm/505-550 nm)下檢測熒光。
1.2.3 農桿菌介導的浸花法遺傳轉化擬南芥 將攜帶有35S:MeMYC2.2:GFP 重組質粒的農桿菌LBA4404 菌株接種至LB 液體培養基中,28℃培養至OD600為1.0。短暫離心收集菌體,然后以懸浮液(MS培養基+60 g/L 蔗糖+0.02% Silwet)重懸菌體。選取生長旺盛、剛剛開花的擬南芥(Col-0),將花序浸入懸浮好的菌液中10 s,20℃保濕狀態下過夜暗培養,然后恢復正常(22℃)光照培養1 周后,重復侵染1 次。待果莢成熟后,收獲T0代種子。篩選壓力(30 mg/L 潮霉素)下篩選T0代種子,獲得T1代轉化苗。隨后T2代經3∶1 分離比篩選培養后,T3代獲得單拷貝純合植株,收取T3代純合子種子備用。提取3 個T3代純合轉化株系葉片RNA,反轉錄后,qPCR 檢測過量表達擬南芥株系中MeMYC2.2 基因的轉錄水平。
1.2.4 轉基因擬南芥株系耐低溫實驗 將篩選的轉基因和野生型擬南芥種子表面消毒(75%乙醇+0.1% Triton X100,10 min)后,無水乙醇洗滌一次,超凈工作臺吹干種子,播種于1/2 MS 平板上,待長至四葉期,移栽于裝有營養土(營養土∶蛭石=1∶2)的營養缽中,置于光照培養房中。待擬南芥葉盤鋪開,進行低溫(-20℃)處理。低溫處理0、1、3 和6 h 后,取葉片于液氮中速凍后,置于-80℃冰箱保存備用。
1.2.5 基因定量表達分析 低溫處理后的轉基因及野生型擬南芥葉片,提取其總RNA,反轉錄成cDNA。進行目的基因檢測及低溫信號途徑相關基因的熒光定量分析。以AtActin1為內參基因。
2 結果
2.1 木薯MeMYC2.2基因的克隆與生物信息學分析
木薯中有12 個MYC2 相似的基因,通過序列比對發現其中2 個(Manes.17G016000,Manes.15G-182700)基因與蓖麻、擬南芥、玉米等的MYC2 親緣關系最近[19]。Manes.17G016000,Manes.15G182-700 分別命名為MeMYC2.1 和MeMYC2.2。采用在線生物信息學軟件對其進行分析,結果如表2。NCBI在線預測蛋白保守功能域發現,MeMYC2.1 和Me-MYC2.2 都具有bHLH-MYC 保守結構域和HLH DNA結合域,屬于典型的MYC 轉錄因子。

表2 MeMYC2 生物信息學分析Table 2 Bioinformatics analysis of MeMYC2
2.2 木薯MeMYC2基因表達模式分析
為探究木薯MeMYC2 在低溫脅迫過程中的作用,我們對野生型木薯組培苗葉片在低溫處理(10℃)不同時間段(0、12、24 和48 h)進行取材。提取葉片總RNA,并反轉錄合成cDNA 作為模板,利用實時熒光定量PCR 技術(RT-qPCR)對MeMYC2.1/2.2基因的表達量進行分析。結果表明,MeMYC2 基因的mRNA 在低溫脅迫12 h 時豐度達到最高,隨后逐漸降低,其中MeMYC2.2 的表達差異更明顯(圖1)。上述結果表明MeMYC2 為低溫脅迫早期誘導基因,而MeMYC2.2 表達差異更顯著。因此選擇MeMYC2.2基因作為目標基因進行進一步研究。
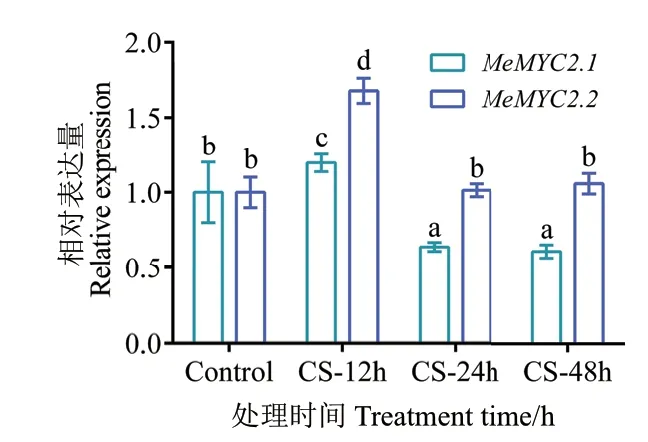
圖1 不同冷脅迫時間下MeMYC2 的表達分析Fig.1 Expression patterns of MeMYC2 under different cold stress(CS)time
2.3 MeMYC2.2可能作為轉錄因子在細胞核中行使功能
MeMYC2.2 蛋白定位預測分析顯示其可能定位于細胞核。為驗證MeMYC2.2 蛋白的亞細胞定位,本研究將MeMYC2.2 基因與GFP 進行融合后,構建相應的植物表達載體。利用煙草葉片瞬時轉化系統驗證蛋白的定位,在農桿菌轉化、共培養3 d 后,使用共聚焦顯微鏡(Olympus FV1000,Japan)在GFP 激發光波長下檢測熒光。結果表明MeMYC2.2蛋白在細胞核中表達(圖2-A)。
為驗證MeMYC2.2 蛋白是否具備轉錄自激活作用,本研究將MeMYC2.2-pGBKT7 轉化酵母Y187感受態細胞,在SD/ -Trp + X-α-gal 培養基上對MeMYC2.2 蛋白的轉錄激活功能進行分析。結果(圖2-B)表明,MeMYC2.2 基因的表達產物能夠激活酵母α-半乳糖苷酶(MeL1)基因的表達,使得酵母細胞在SD/ -Trp + X-α-gal 平板上形成藍色菌斑,說明MeMYC2.2 具備轉錄自激活功能,結合MeMYC2.2 蛋白定位于細胞核的結果,由此推斷MeMYC2.2 可能是一個轉錄因子。
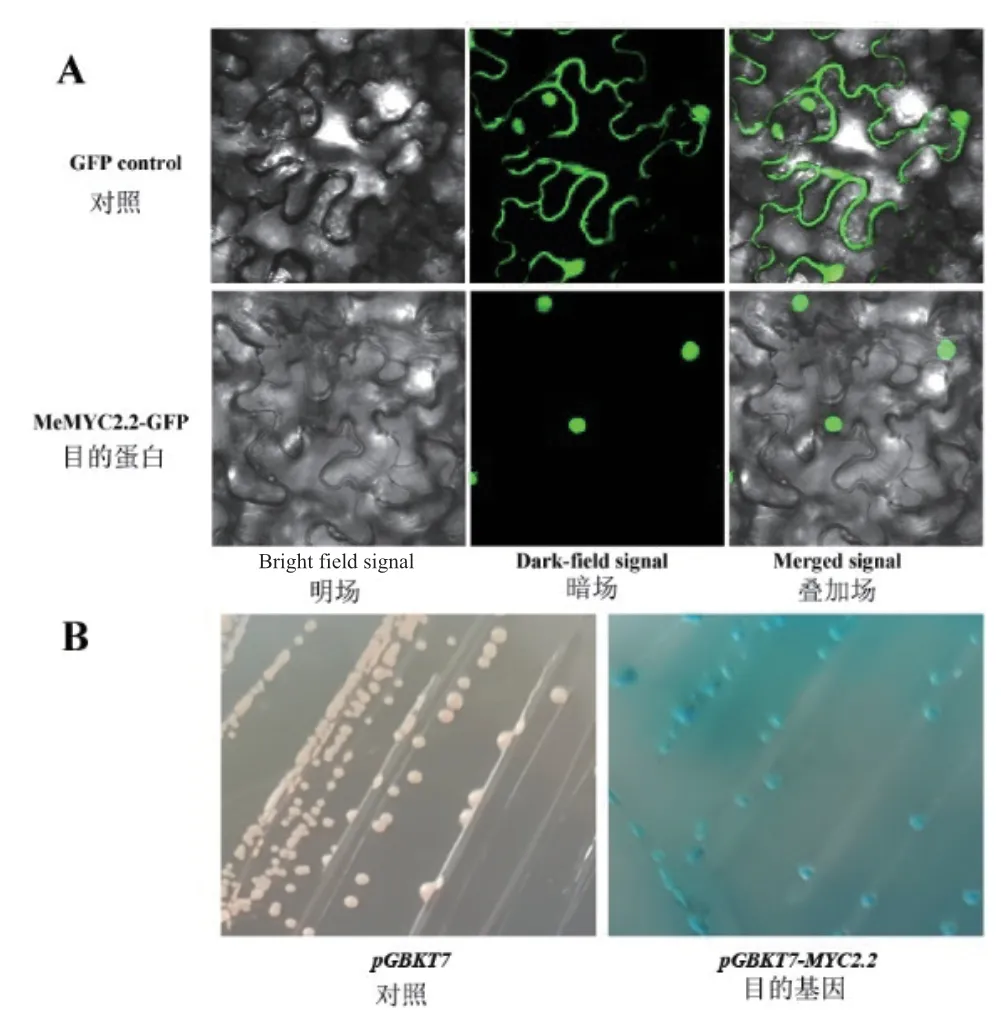
圖2 MeMYC2.2 轉錄因子驗證Fig.2 Validation experiments of transcription factor MeMYC2.2
2.4 MeMYC2.2過量表達能夠提高擬南芥植株抗凍能力
采用農桿菌介導的浸花轉化方法,獲得MeMYC2.2 過量表達轉基因擬南芥。經潮霉素抗性篩選獲得的16 個T1代陽性轉基因擬南芥,PCR 檢測為陽性。T2代轉基因擬南芥進一步在潮霉素抗性平板上通過3∶1 的篩選比,獲得單拷貝轉基因株系;T3代轉基因擬南芥繼續抗性篩選,最終得到單拷貝的轉基因擬南芥純合子。選取3 個T3代轉基因純合株系,采用RT-qPCR 方法檢測過量表達株系中MeMYC2.2 基因的轉錄水平。結果(圖3)顯示,3個被檢株系中MeMYC2.2 基因皆被高水平轉錄。
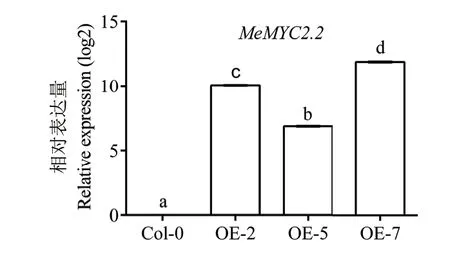
圖3 轉基因擬南芥中MeMYC2.2 表達分析Fig.3 Expression analysis of MeMYC2.2 in transgenic Arabidopsis plants
為驗證MeMYC2.2 基因的過量表達對擬南芥植株抗凍性的影響,我們采用-20℃模擬凍害。結果(圖4)顯示,未經凍害處理時,MeMYC2.2 過量表達轉基因擬南芥植株和野生型在形態特征上未有明顯差異。冷凍處理4 h 的野生型擬南芥葉片逐漸開始出現萎蔫;而冷凍處理6 h 的野生型擬南芥葉片大面積出現枯萎,凍害嚴重,植株死亡率較高。而冷凍處理4 h 的MeMYC2.2 過量表達轉基因擬南芥葉片可以正常伸展;如表3所示,與野生型對比,凍害延續至6 h 的轉基因擬南芥凍害程度較輕,死亡率顯著降低。經冷凍處理后,在22℃條件下恢復生長1周后,MeMYC2.2 過量表達轉基因擬南芥的成活率顯著高于野生型擬南芥。

表3 野生型和轉基因植株冷凍處理后的存活率統計Table 3 Survival rates of wild-type and transgenic plants with cold treatment (n>30)

圖4 轉基因和野生型植株表現出不同的耐凍能力Fig.4 Wild-type and transgenic plants demonstrating different freezing tolerance abilities
由此可見,MeMYC2.2 基因的過表達對轉基因擬南芥抗凍能力的提高有正調控作用。
2.5 CBF基因在轉基因擬南芥中的表達分析
對冷凍脅迫(-20℃)下MeMYC2.2 過表達擬南芥及野生型擬南芥葉片中CBF基因的表達量進行RT-qPCR 分析。結果(圖5)顯示,與野生型擬南芥相比,CBF3基因的表達在MeMYC2.2 過表達擬南芥中受低溫誘導更為明顯。但其他3 個CBF基因在轉基因擬南芥中受低溫誘導的表達量明顯低于其在野生型擬南中的表達量。
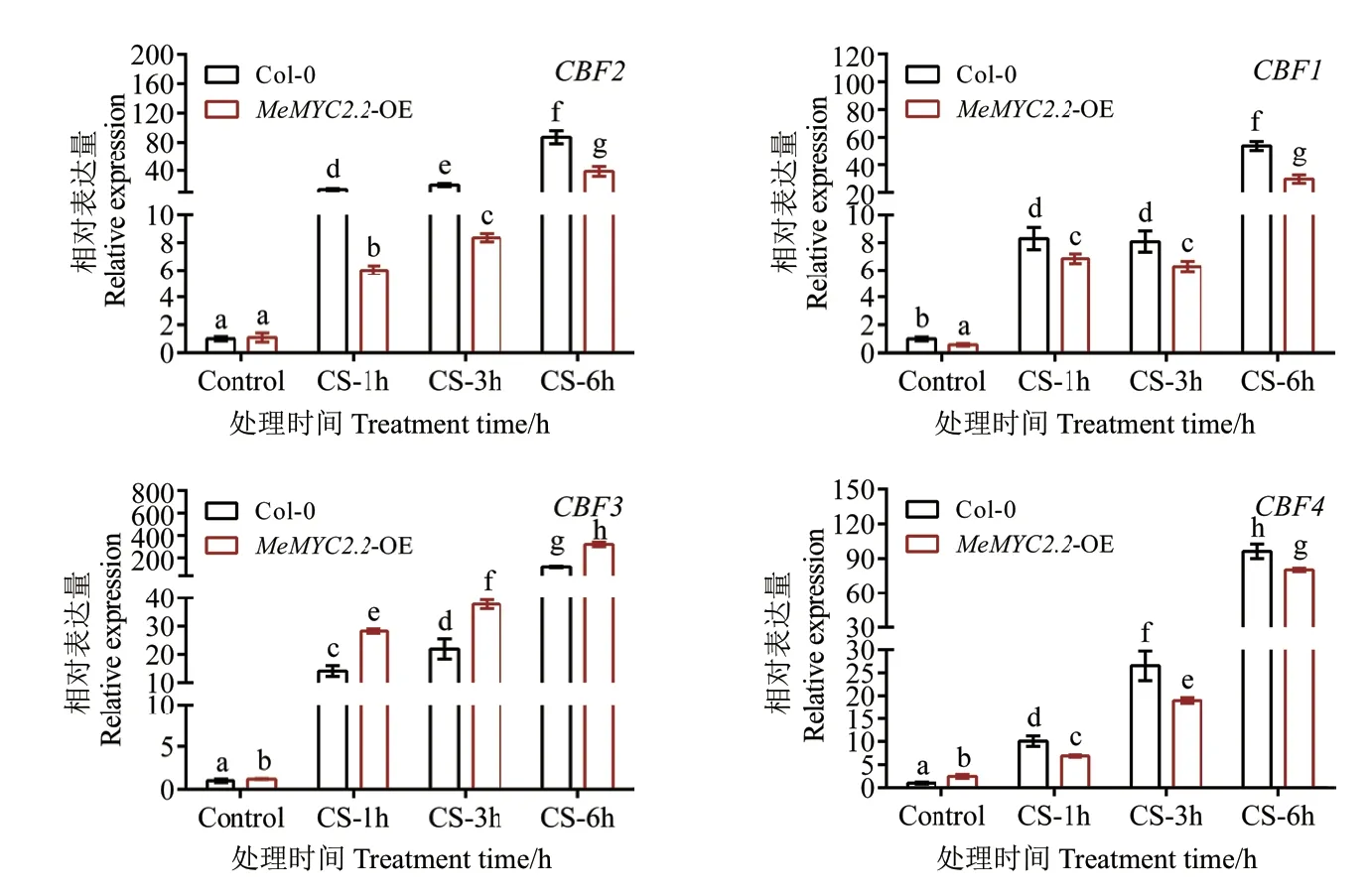
圖5 冷凍脅迫下轉基因擬南芥中CBF 基因的表達分析Fig.5 Expression analysis of CBF genes in transgenic Arabidopsis under cold stress
3 討論
作為一種熱帶塊根作物,木薯原產于溫暖的熱帶地區,被歸類于對寒冷敏感的物種[20]。低溫對木薯儲藏根及種莖損害極大,2008年1月份我國南方發生的冷凍災害,導致廣西、廣東等省木薯產業造成巨大的破壞[21]。在10℃以下低溫,木薯表現出明顯的損害癥狀,包括產量下降、葉片擴展減少、褪綠甚至死亡[22]。本研究采用10℃做木薯低溫脅迫處理與生產實際密切相關,并且對篩選基因做抗寒機理研究非常重要。
MYC2 轉錄因子是bHLH 轉錄因子家族成員之一,廣泛存在于動植物中。MYC2 在植物應答逆境刺激時起重要調控作用,是JA 信號傳導途徑的核心調控因子,同時也是JA 與其他植物激素信號交互的關鍵節點[13]。已有研究表明,擬南芥MYC2 轉錄因子通過特異性結合CBF3啟動子上的MYC 結合位點(CANNTG),正調控CBF3基因的表達[23]。大量研究證明CBF基因在植物抗寒方面是一個保守的信號途徑,被認為是調控非生物脅迫轉錄因子中的核心調控因子[8]。CBF 轉錄因子屬于AP2/ERF多基因家族,是植物抗凍性的關鍵調節因子[24-25]。CBF 轉錄因子能直接激活一系列提高冷凍耐受性的COR(冷調節)基因[26-28],對于植物的冷馴化和抗凍性至關重要。已有研究顯示,干擾CBF3 基因后,二穗短柄草在低溫下的存活率顯著降低[29];而增強CBF3的表達,植株表現出更好的抗凍性[30-31]。
栽培木薯對低溫敏感,但其低溫敏感的分子基礎尚不明確。CBF參與了多個低溫響應基因(COR)表達的調控,直接影響植物的抗寒性。基因表達分析發現CBF基因在木薯頂芽中的表達缺失,這可能是造成木薯低溫敏感的原因之一[20]。在擬南芥中,CBF基因表達在低溫處理15 min 內顯著上調,在2 h 后達到最高[32]。而木薯CBF基因表達對低溫脅迫的響應明顯滯后,需要數個小時后才開始響應,且其表達要在8 h 以上才達到最高[33]。過表達木薯CBF1基因的轉基因植株對低溫的耐受性明顯提高[33];研究也發現過表達木薯CBF4基因也可以提高轉基因植物的抗寒性[34]。這說明木薯CBF基因同樣具有調控抗寒性的功能,但木薯中CBF表達調控途徑無法迅速響應低溫脅迫信號。
為研究MeMYC2.2 轉基因擬南芥抗凍性顯著增強的原因是否與CBF 信號途徑關聯,對冷凍脅迫(-20℃)下MeMYC2.2 過表達擬南芥及野生型擬南芥葉片中CBF基因的表達量進行RT-qPCR 分析。結果顯示,MeMYC2.2 過表達擬南芥植株中,僅CBF3的表達量較野生型植株顯著提高,且隨著冷凍脅迫時間的持續,MeMYC2.2 過表達擬南芥中CBF3 的表達豐度持續升高。這說明木薯MeMYC2.2 具有調控CBF 途徑影響低溫耐受性的可能性。栽培木薯中MeMYC2.2 的表達雖然對低溫有響應,但其差異表達量變化倍數并不高,這一點是否與低溫脅迫下木薯CBF基因表達模式有關還需要進一步研究。
4 結論
木薯MeMYC2.2 基因,屬于bHLH 轉錄因子家族成員,其表達受低溫誘導。在轉基因擬南芥中過表達MeMYC2.2 可上調CBF3 基因對低溫脅迫的響應,從而增強轉基因擬南芥植株對低溫的耐受性,為進一步利用MeMYC2 基因改良木薯的低溫耐受性奠定理論基礎。
