紅樹林沉積物反硝化和厭氧氨氧化速率的時空變化及環境響應
2023-01-18 09:26:22林雨彬秦永強馬思遠曾若菡陳雨梅刁曉平2
熱帶生物學報 2023年1期
林雨彬,秦永強,馬思遠,曾若菡,陳雨梅,刁曉平2,
(1.海南大學 生態與環境學院,海口 570228;2.海南大學 南海海洋資源利用國家重點實驗室,海口 570228;3.海南師范大學 生命科學學院,海口 571158)
由于沿海養殖業發展、施肥量增加和土地利用的變化,地表水污染程度穩步上升[1],導致徑流污染嚴重。濕地作為天然的環境屏障,能夠降低陸地向海洋的徑流污染排放。紅樹林生態系統是全球濕地的重要組成部分[2],由于其高效的營養循環系統而具有極高的生產力[3],能夠凈化和過濾由內陸向海洋的徑流中的有機物質和營養鹽[4]。1960年以來,不合理的開發活動使得紅樹林面積銳減[5]。由于其獨特的地理環境和系統特點[6],近年來紅樹林的研究日益增加,在紅樹林的清退修復工作及可持續發展方面,先后實施了藍碳戰略和“藍色港灣”、“南紅北柳”等生態工程[7]。
氮素循環是整個生物地球循環的重要組成部分,對維持整個生物圈的生態平衡意義重大。氮循環過程主要有硝化過程、固氮過程、反硝化過程、硝酸鹽銨化過程、厭氧氨氧化過程和氨化過程等重要步驟。其中反硝化和厭氧氨氧化過程被廣泛認為是環境中活性氮去除的最主要途徑[8],其中反硝化是全球氮循環的關鍵步驟,其貢獻遠遠超出其他過程。反硝化是在低氧條件下的反硝化菌將硝酸鹽或者亞硝酸鹽轉化為氮氣或者氧化二氮的過程,可以降低活性氮,維護氮循環平衡[9]。厭氧氨氧化則指在厭氧條件下,以亞硝酸鹽作為電子受體,將氨氮氧化成氮氣的過程。不同種類的生態環境中,厭氧氨氧化在氮循環中的貢獻比例有較大差異,但仍舊不可忽視[10]。反硝化和厭氧氨氧化作用能將固定態的氮素轉化為氣態氮歸還大氣圈,被廣泛認為是海洋生態系統中活性氮去除的兩個主要途徑,在維持各個圈層的氮素平衡中極其重要[11],而微生物是物質循環的主要驅動者。隨著越來越多針對紅樹林物質循環的研究,人們對紅樹林在全球生物化學循環中的作用越來越重視。氮素作為生物的必須元素,往往是濕地沉積物中的主要限制性養分,因此研究不同生態系統氮循環具有十分重要的意義。已有部分研究發現,反硝化和厭氧氨氧化受到溫度、溶解氧(DO)、硝氮、pH、有機碳、總鐵等許多因素的影響,存在明顯的時空異質性[12-13]。為了研究紅樹林退塘還林過程對沉積物氮循環速率的影響,實驗選取海南省文昌市丹場村紅樹林清退工作區域,探究紅樹林修復過程的不同生境對沉積物理化因子、反硝化及厭氧氨氧化速率的影響,旨在為紅樹林的生態修復和清退工作的效果及階段評估提供科學支持。
1 材料與方法
1.1 研究區概況文昌市紅樹林地處海南省東北部,地理坐標為110°30′~110°02′E,19°15′~20°09′N。屬于熱帶海洋性季風氣候,全年日照時間長,年均氣溫約24.1℃,年均降雨量約1750 mm,雨季為5—10月,旱季11至翌年4月[14]。文昌市自2017年開始啟動保護紅樹林工程,計劃3內退塘還林還濕760 hm2。本研究在2021年3月(旱季)及8月(雨季)于文昌市丹場 村(19.62°N,110.81°E)的原生紅樹林濕地(原生區)、紅樹林修復區(修復區)及退養魚塘區域(魚塘區)3個區域選取樣點進行采集,原生紅樹林濕地未進行開墾養殖,優勢樹種為對葉欖李(Laguncularia racemosa),修復區為進行了人工種植的幼年紅樹區域,魚塘區為已經退養的原養殖魚塘(已經推翻圍墻與外界紅樹林水體相連接)。
1.2 樣品采集采樣使用內徑8 cm、長50 cm的透明亞克力管垂直采集10~25 cm深度的紅樹林沉積物,在每個樣點進行3次隨機采樣,帶回實驗室后將平行樣在厭氧培養箱中使用聚乙烯無菌密封袋進行混勻,混勻后將樣品存放于4℃冰箱,等待進行速率測定和相關理化性質測定[15]。
1.3 環境因子測定沉積物的溫度、鹽度、pH等在現場用土壤YSI(COMBI5000)采集數據。沉積物可溶性硝酸鹽氮(NO3--N)、氨氮(NH4+-N)的測定參照《中華人民共和國國家環境保護標準》(HJ634-2012)進行。沉積物TOC使用重鉻酸鉀外加熱法進行測定[16]。實驗吸光度使用Perkin-Elmer Lambda-25分光光度計進行測定。所有環境參數進行3次重復測定。
1.4 反硝化與厭氧氨氧化速率測定沉積物反硝化及厭氧氨氧化速率采用同位素示蹤法結合泥漿培養實驗測定[15],將沉積物與同鹽度人工海水按照1∶7的質量比例混勻制成均質泥漿,通入氦氣充分曝氣20 min以上,使用無菌注射器將泥漿注入到Labco頂空進樣瓶內,放入與原位溫度相同的恒溫培養箱內預培養24 h。選取其中1個小瓶測定硝酸根離子濃度,用以計算Fn(Fn=100×15NO3-/14+15NO3-),其余所有小瓶加入0.1 mL濃度為12.5 mmol·L-1的15NO3-,加入后充分搖勻,起始樣加入200μL質量分數為50%的ZnCl2,終止樣繼續培養8 h之后加入ZnCl2。反硝化與厭氧氨氧化速率通過培養過程產生的溶解性氣體29N2和30N2來確定。溶解性氣體29N2和30N2采用薄膜進樣質譜儀(Membrane Inlet Mass Spectrometry,MIMS)測定。
1.5 統計分析使用IBM SPSS Statistics 25軟件對環境因子與速率的相關性數據進行Person和Spearman分析,并使用Origin 2018以及Adobe Illustrator 2021進行實驗結果的柱狀圖繪制,使用word插件對數據進行單因素方差分析以確定顯著性關系。
2 結果與分析
2.1 環境因子時空分布特征采樣區域不同生境沉積物理化因子隨著季節和生境的變化而發生改變,并存在時空分布差異(圖1)。除TOC外,其余理化因子均受到季節的顯著影響;所有理化因子都受到采樣生境的顯著影響。
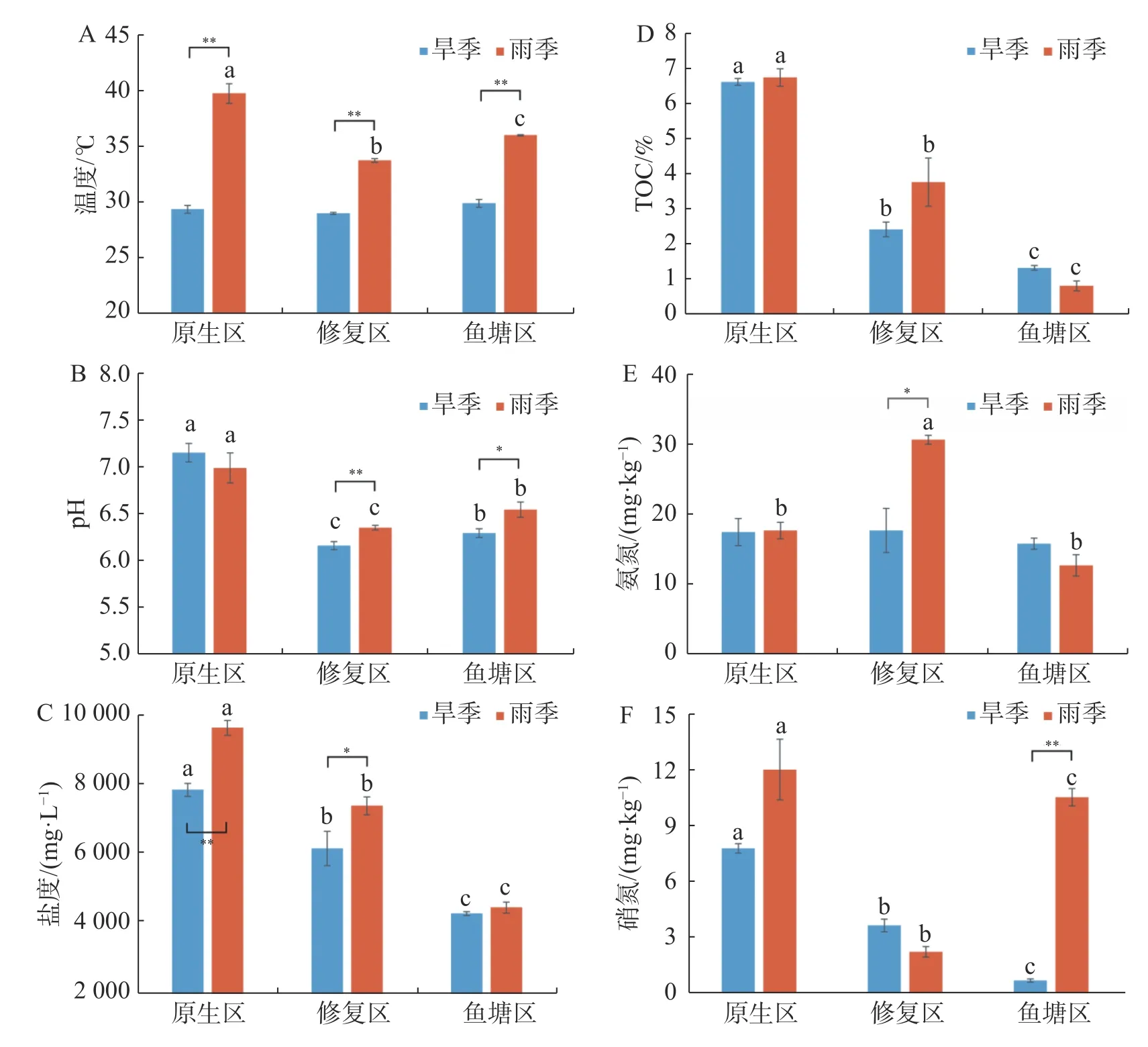
圖1 不同季節各生境沉積物的理化指標
沉積物溫度范圍為旱季29.03~29.93℃,均值29.46℃、雨季為33.80~39.84℃,均值36.57℃,不同生境沉積物溫度存在極顯著季節差異(P<0.01),雨季采樣點之間也存在極顯著差異(P<0.01)。
旱季沉積物pH范圍為6.17~7.17,均值6.55;雨季pH為6.36~7.01,均值6.64,其中修復區pH存在極顯著的季節差異(P<0.01),魚塘區存在顯著的季節差異(P<0.05)。沉積物pH呈現原生區極顯著高于魚塘區(P<0.01)和魚塘區顯著高于修復區的趨勢(P<0.05)。
旱季沉積物鹽度范圍為4338~7902 mg·L-1,均值6151 mg·L-1;雨季為4509~9691 mg·L-1,均值7214 mg·L-1。總體呈原生區高于修復區和修復區高于魚塘區的趨勢,并且除旱季原生區和修復區差異顯著外,其余差異均極顯著。沉積物鹽度在原生區表現出極顯著的季節差異(P<0.01),修復區表現出顯著的季節差異(P<0.05)。
旱季沉積物氨氮(NH4+-N)范圍為15.92~17.59 mg·kg-1,均 值17.11 mg·kg-1,雨 季12.78~30.93 mg·kg-1,均值為20.51 mg·kg-1。旱季修復區氨氮顯著地低于雨季(P<0.05),且雨季修復區顯著極顯著地高于其余2個區域(P<0.01)。
旱季沉積物硝氮(NO3--N)范圍為0.72~7.86 mg·kg-1,均 值4.09 mg·kg-1,雨 季2.27~12.11 mg·kg-1,均值8.33 mg·kg-1。整體兩個季節沉積物硝氮具有顯著的季節差異(P<0.05),原生區和修復區季節差異不顯著,魚塘區季節差異極顯著(P<0.01)。且生境之間均具有極顯著的差異(P<0.01)。
相同生境的沉積物TOC沒有季節性差異,不同生境的沉積物TOC存在明顯的季節性差異(P<0.05),原生區沉積物TOC均顯著高于修復區和魚塘區(P<0.05)。
2.2 反硝化速率與厭氧氨氧化速率的時空分布不同生境紅樹林沉積物的反硝化和厭氧氨氧化速率見圖2。由圖2可知,沉積物速率指標之間存在較大時空差異。旱季反硝化速率區間為8.08~14.39 nmol·g-1·h-1,其中修復區的反硝化速率最高(14.39 nmol·g-1·h-1),顯著高于原生區(P<0.05);雨季反硝化速率則介于9.62~11.67 nmol·g-1·h-1之間,無顯著生境差異。同生境下反硝化速率沒有顯著季節差異。
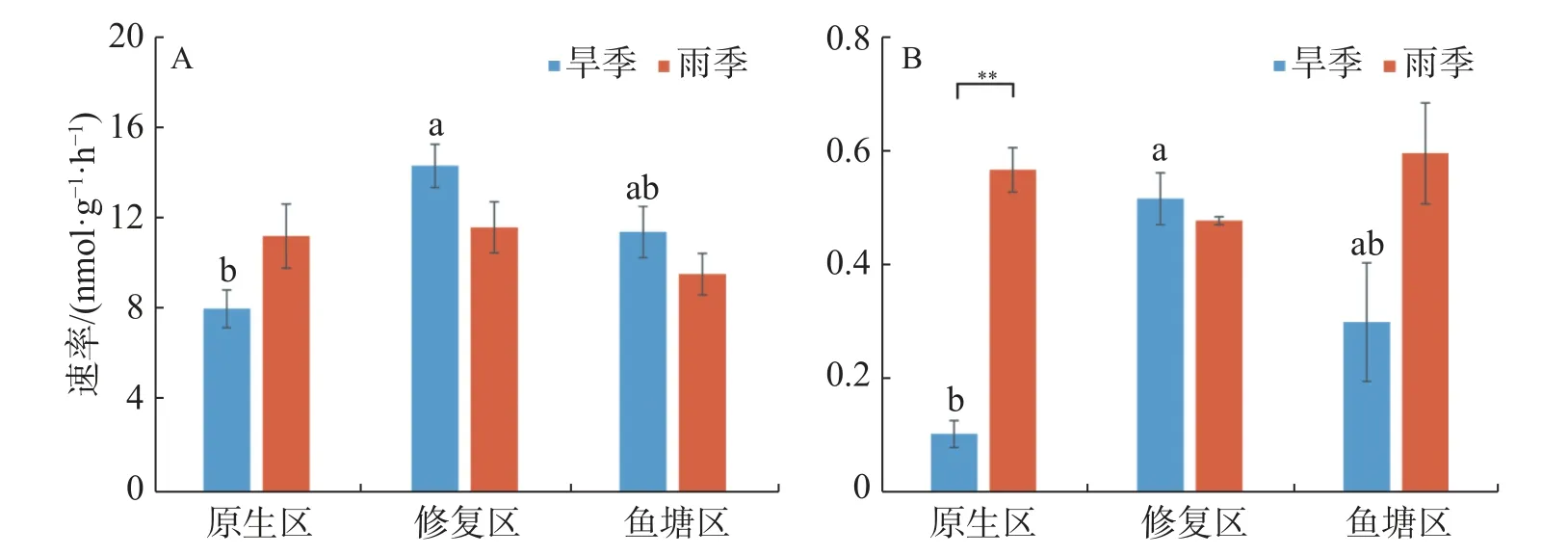
圖2 不同季節各生境沉積物反硝化(A)及厭氧氨氧化速率(B)
旱季厭氧氨氧化速率區間為0.10~0.52 nmol·g-1·h-1,修復區的反硝化速率最高(0.52 nmol·g-1·h-1),顯著高于原生區(P<0.05)。雨季厭氧氨氧化速率介于0.48-0.60 nmol·g-1·h-1之間,不同生境之間沒有顯著差異。原生區厭氧氨氧化速率存在極顯著的季節差異,雨季明顯高于旱季(P<0.01)。
2.3 不同生境環境因子對反硝化及厭氧氨氧化速率的影響沉積物反硝化及厭氧氨氧化速率受到多項因子的綜合影響,本研究結果(表1)表明,反硝化和厭氧氨氧化速率呈現高度正相關(雨季修復區除外);氨氮和硝氮對3種不同生境沉積物的反硝化和厭氧氨氧化速率均有顯著影響。選取部分與脫氮速率相關度較高的理化因子與反硝化和厭氧氨氧化速率進行RDA分析,結果如圖3所示,RDA1軸解釋度為78.13%,RDA2軸解釋度為10.10%。結果表明,溫度和沉積物硝氮含量對反硝化和厭氧氨氧化速率產生了重要影響,沉積物氨氮含量的影響則較小。
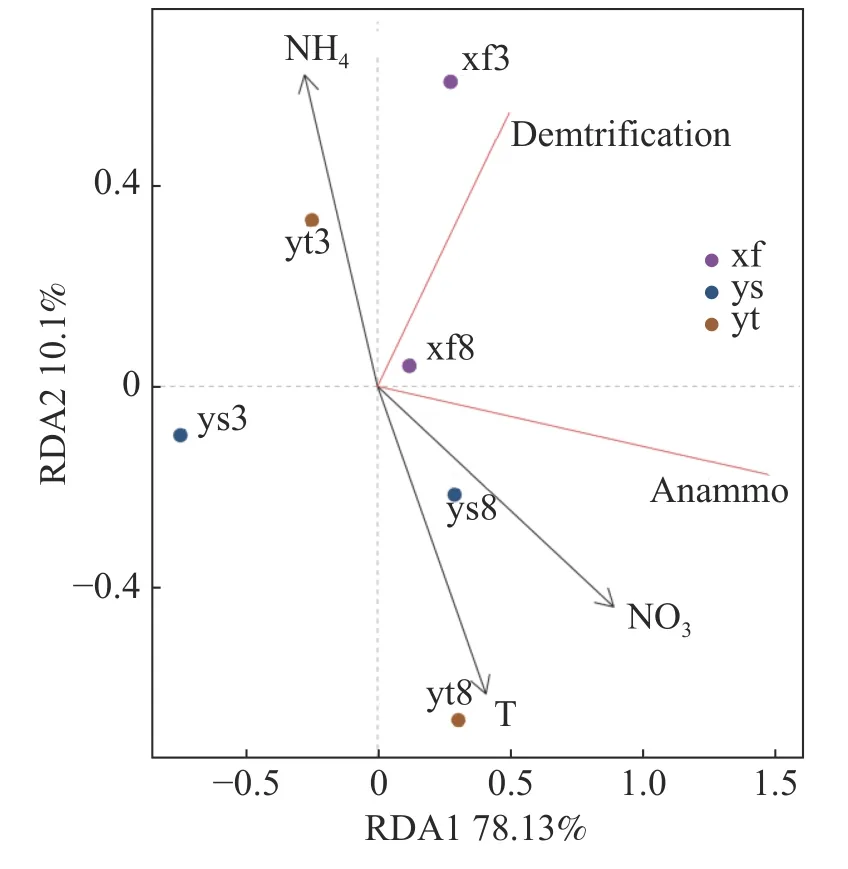
圖3 部分理化因子和反硝化及厭氧氨氧化速率的RDA分析
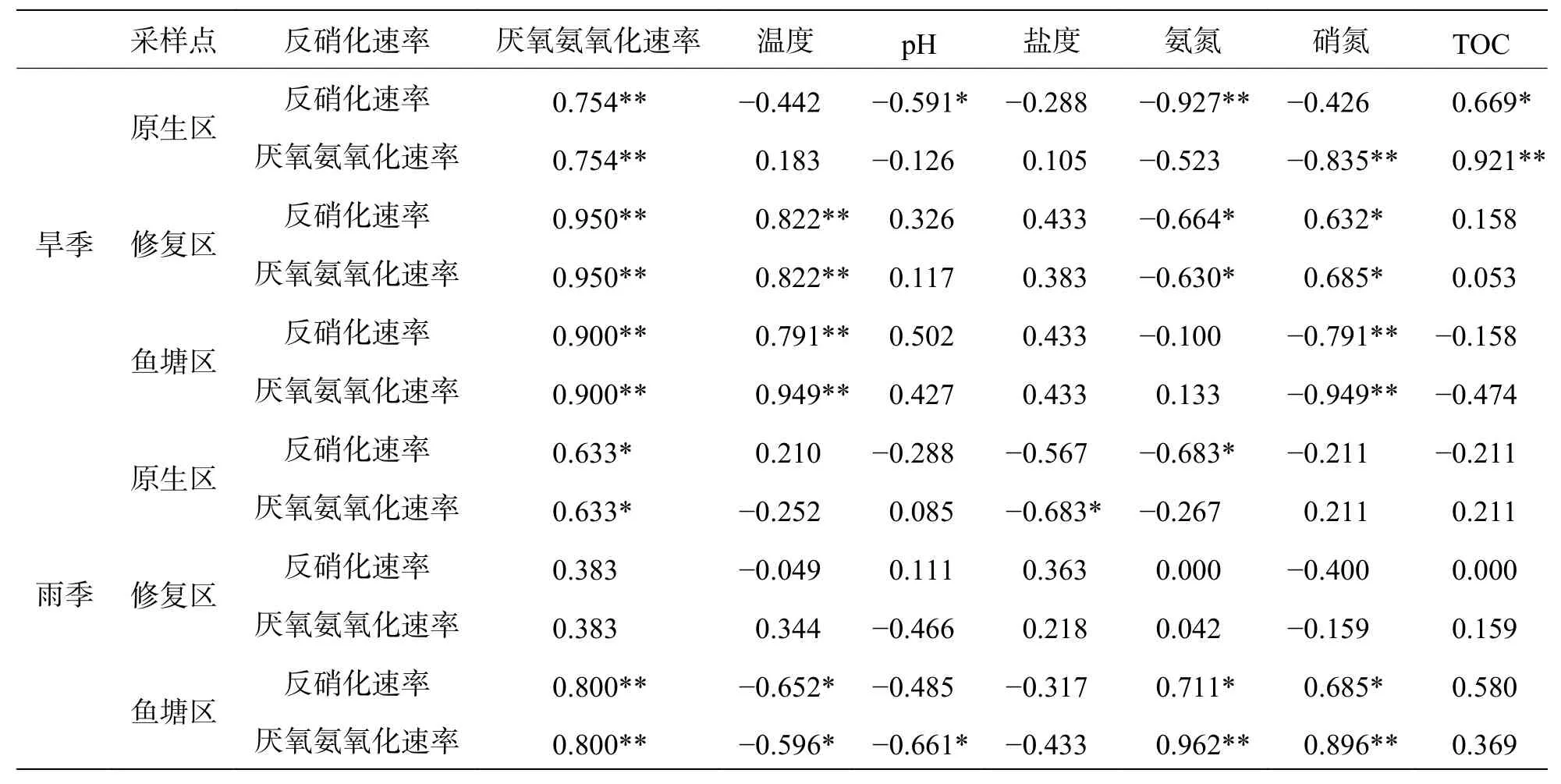
表1 基于Spearman分析的N循環速率與沉積物理化因子的關系
3 討 論
研究區域沉積物pH低于海水的pH,整體沉積物偏酸性是紅樹林的特色,也可能與附近養殖魚塘廢水排放污染有關,3個區域中原生區受影響最小,故pH最高。而魚塘區由于質地呈砂紙,淤泥含量少,有機質含量也較少(如TOC),產生的還原性氣體較修復區少,故酸性較弱。區域鹽度差異是由于海陸距離差異導致,內陸徑流多為淡水,在紅樹林入海過程存在鹽度漸變過程。紅樹林沉積物出現溫度、pH、鹽度等理化因子的季節性差異與Li等[17]的研究相同。旱季修復區較高的氨氮可能是因為生物活動較旱季更活躍而導致,氨是魚類和加殼類生物主要的代謝產物之一,導致沉積物氨氮含量升高[18]。其中,雨季魚塘區硝氮的異常高含量可能是由于采樣區域現有養殖魚塘進行排水換水導致。
有研究表明,細顆粒沉積物對有機質的富集能力更強[19],筆者在采樣時發現,魚塘區域的沉積物大部分為粗糙顆粒,這可能是導致該區域TOC含量最低的原因。土壤反硝化過程主要是由放線菌門( Actinobacteria)、變形菌門(Proteobacteria)、厚壁菌門(Firmicutes)、擬桿菌門(Bacteroidetes)等微生物群調控,而厭氧氨氧化微生物主要歸為浮霉菌門(Planctomycetes)[19],種類繁多的微生物參與調控脫氮并且發揮重要作用[20],而溫度、水分、pH、溶氧、資源可用性、植被類型等均會對微生物種群多樣性和豐度產生不同的效應[21],因此脫氮是許多不同的微生物與環境共同協作的結果。本研究發現修復區是反硝化和厭氧氨氧化速率最高的生境,可能是由于原生區過高的鹽度抑制了該生境氮循環相關微生物,如厭氧氨氧化菌的活動導致該生境反硝化和厭氧氨氧化速率最低[17,22],王勇等[23]發現厭氧氨氧化的最適環境溫度為(35±1)℃,而本研究厭氧氨氧化在原生區的極顯著季節差異也可能是因為雨季的沉積物溫度極顯著地高于旱季,因為雨季沉積物溫度更加接近最適值。且該處沉積物硝氮雨季高于旱季,反硝化潛力大[24],有更高的反硝化速率[25],提供了更多的厭氧氨氧化反應底物。溫度和硝氮同時作用大幅提高了該生境的厭氧氨氧化速率。也可能是此處微生物群落結構受其余未測定環境因素和人類活動的影響所致[26],故不同生境氮循環速率的差異性不能局限于單一理化因子的影響。
兩項速率出現的耦合效應與大多數研究相似,該關系的出現可能歸結于反硝化在厭氧條件下提供了厭氧氨氧化的底物NO2-。有研究發現隨溫度升高,反硝化速率提高[27],本研究在旱季的修復區和魚塘區,反硝化速率和溫度呈現極顯著的正相關關系,同時該區域的厭氧氨氧化速率同樣受到溫度的密切影響,與大多數結果相似。但可能由于采樣生境整體溫度較高,整體未能成為限制N循環速率的關鍵因素。反硝化作用的最佳pH為7~8,研究表明,高于或低于這個范圍反硝化速率將會下降[28]。本實驗除原生區外其余生境的pH均低于該范圍,因此在旱季原生區和雨季魚塘區反硝化速率出現了明顯的下降。雨季原生區沉積物的鹽度最高,該生境反硝化速率下降與鹽度有關。有研究表明,鹽度升高會導致反硝化速率的降低[20],本研究結果可能是該處鹽度極顯著高于其他點而抑制微生物活動所致(P<0.01)。
硝氮和氨氮會顯著地對反硝化速率造成影響[29-30]。一般來說,沉積物反硝化速率與離子交換態(IEF)硝氮和氨氮會存在較為明顯的正相關性[31],而分析結果表明僅有雨季魚塘區存在該正相關。本實驗結果表明紅樹林沉積物中硝氮賦存形態較為特殊,在雨季原生區和修復區可能有較多碳酸結合態(CF)及氧化物態的(IMOF)硝氮和氨氮,導致所測沉積物硝氮和氨氮含量與速率并未呈現大多數研究的正相關關系,也可能是采樣地點水質營養素較豐富,硝態氮未成為反硝化與厭氧氨氧化速率的限制因素,反而超過正相關閾值導致。張波等[32]研究發現太湖反硝化速率與硝氮存在負相關關系,認為反硝化速率越大,厭氧程度越高而導致硝氮含量較低,沉積物的高反硝化速率會降低環境硝氮負荷。吳耀國[33]研究結果表明,適當增加有機物可以提高反硝化速率,而本研究中雨季修復區較高的反硝化速率與沉積物TOC含量較高有一定相關性。
4 結 論
不同生境紅樹林區域理化因子存在復雜的時空變化特征,其中修復區各項理化因子相較于另外2個生境更低且季節性差異最為明顯,原生區理化因子受季節影響較小且除沉積物氨氮以外的理化因子數值均為原生區最高。沉積物反硝化速率具有空間差異,厭氧氨氧化速率具有時空差異。反硝化及厭氧氨氧化速率受到不同采樣生境、理化因子等的重要影響,反硝化和厭氧氨氧化速率均在修復區最高,原生區最低,總體季節差異不顯著,反硝化和厭氧氨氧化速率之間有十分明顯的耦合特性。所測理化因子中溫度、pH、鹽度、電導率、氨氮、硝氮、TOC均與紅樹林脫氮過程存在較為復雜的關系,其中溫度和硝氮與反硝化和厭氧氨氧化過程關系較緊密,是該實驗中紅樹林脫氮過程的關鍵影響因素。
猜你喜歡
歌海(2024年6期)2024-03-18 00:00:00
音樂探索(2022年2期)2022-05-30 21:01:37
兒童故事畫報·自然探秘(2022年3期)2022-04-27 00:17:10
幼兒園(2021年4期)2021-07-28 07:38:04
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
瘋狂英語·初中天地(2018年6期)2018-11-24 02:39:24
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
歌海(2018年4期)2018-05-14 12:46:15
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
